The data set consists of ~92,000 measurements of cotton canopy temperature collected over a 120-day period from July through September, with ~11,000 measurements per variety in each of four treatments. Thus, while the measurements are not replicated, the magnitude of the dataset allows for a detailed analysis of canopy temperatures over a production season.
3.2. Seasonal Canopy Temperatures
It was expected that canopy temperatures of the two leaf shapes would differ under certain conditions and would be more similar under others. In an effort to accentuate differences, different water regimes were implemented and the season was divided into several periods. As the season progressed and the water demand of the crop increased, the water deficits in the rainfed, 1.5 mm, and 3.0 mm irrigation regimes increased in intensity. The effect of water on temperature differences over the season should be minimal in the 6 mm treatment. From a seasonal perspective, in addition to the 120-day period (full-season period), the experimental period was divided into three sequential 40-day periods for analysis in an effort to detect canopy-relates developmental differences in the canopy temperature patterns. To understand diurnal influences, the canopy temperature data was also divided into 24-h, day, and night periods on the basis of solar radiation.
Figure 2 shows the canopy temperatures over the 120-day measurement period beginning 50 DAP. The effect of irrigation on the canopy temperatures is clearly evident with temperatures above 28 °C more common as irrigation level declines. The horizontal line on the figure indicates 28 °C which represents the optimal temperature for cotton [
16]. Around DOY 250 (7 September) the temperatures declined in response to end-of-season changes in the weather as evidenced in the air temperatures.
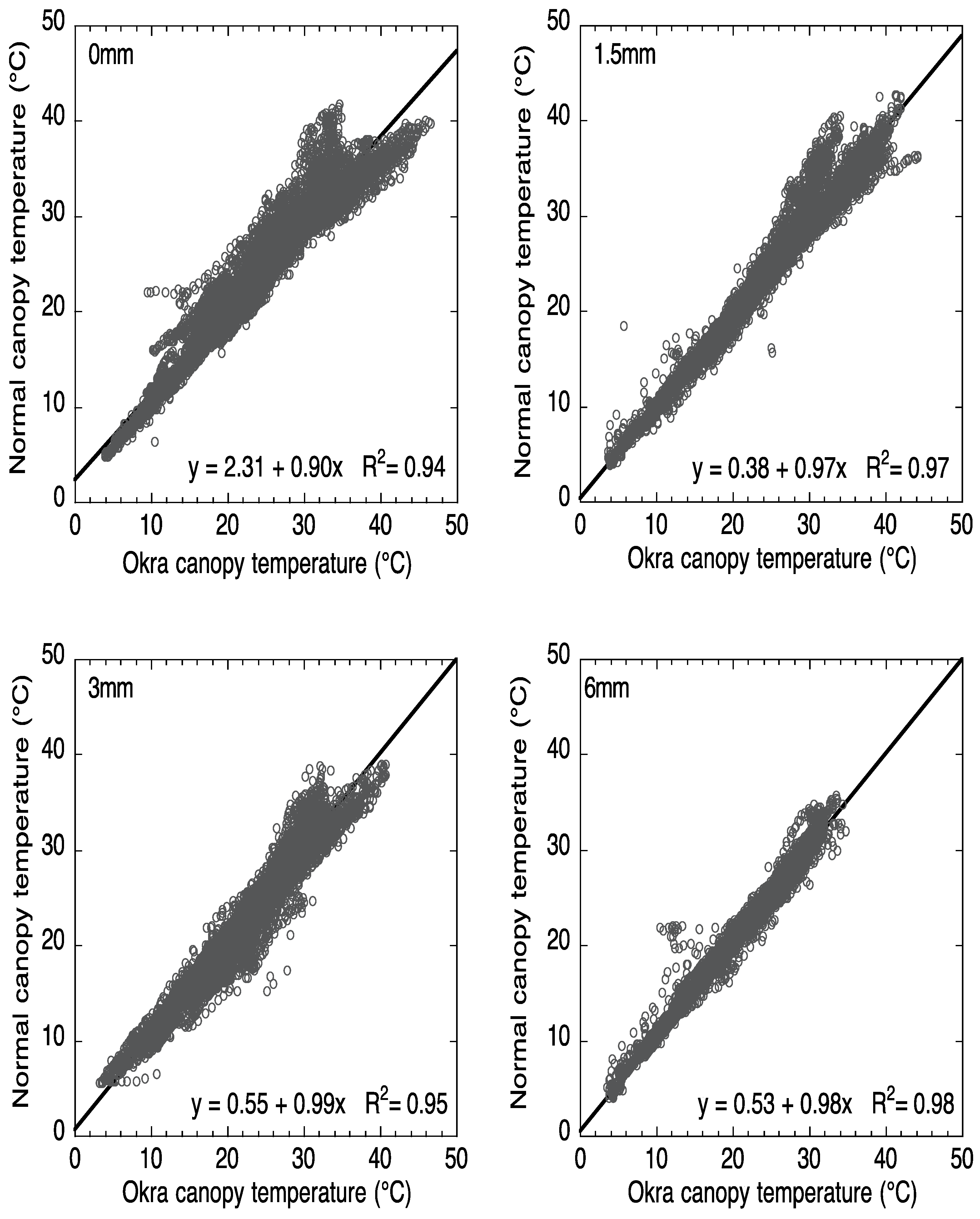
Figure 3 compares the canopy temperature of the normal and okra leaf over the 120-day Full-season period. As the irrigation level increased the canopy temperatures of the two varieties became more similar. In the 1.5 mm, 3 mm, and 6 mm irrigation treatments the y-intercepts are <0.55 °C which is near the resolution limit of the IRTs, while it is higher, at 2.3 °C, in the 0 mm condition. The
R2 value is lowest for the 0 mm (0.94) and >0.95 for the 1.5 mm, 3 mm, and 6 mm treatments. Thus, for the entire study period there are differences in canopy temperature between the varieties but they are generally small.
Table 1 shows the air and canopy temperatures over the study period. In the “full”, “early”, and “mid” periods of the study, there was a trend of lower average canopy temperatures as irrigation amount increased. The maximum temperature followed a similar pattern, as well. These trends were not as pronounced in the “late” period measurements, perhaps indicating that at season’s end the water deficits were diminished. The trends in canopy temperature were similar for both leaf shapes. Canopy temperatures for day and night periods over several multi-day periods of the season were analyzed using a two-factor ANOVA. Over the full-season period, the canopy temperatures were significantly different (
p < 0.05) during the night period while differences were not significant (
p > 0.05) during the day period. Over the full-season period, the effect of irrigation was significant (
p < 0.001) for both varieties for both the night and day periods. Over the full-season period, the interaction between variety and irrigation was significant (
p < 0.01) for both the night and day periods. The full-season period was divided into three 40-day periods for analysis that were not analyzed in terms of night and day periods. During the early-season period the canopy temperatures were significantly different (
p < 0.01) for varieties and irrigation. The interaction between variety and irrigation was significant (
p < 0.001). During the mid-season period, the canopy temperatures were not significantly different (
p < 0.01) for variety. Both the effect of irrigation and the interaction between variety and irrigation was significant (
p < 0.01). During the late-season period, the canopy temperatures were significantly different (
p < 0.05) for variety and irrigation with a significant (
p < 0.001) interaction between variety and irrigation. While instances of significant differences between the varieties were detected in all of the periods, irrigation treatments, and times of day, they were generally small (<1 °C) and their relevance in an agricultural context is questionable.
The normal leaf and okra leaf canopy temperature pairs at each point in time were compared using a paired
t-test. The results of these comparisons are shown in
Table 2 with significant differences (
p < 0.001), denoted with ***. With the exception of four instances in the 0 mm treatment and one instance in the 1.5 mm, the canopy temperatures were significantly different between the okra and normal leaf types. The differences are generally quite small and close to the resolution of the IRTs and, thus, their importance could be questioned from an agronomic perspective.
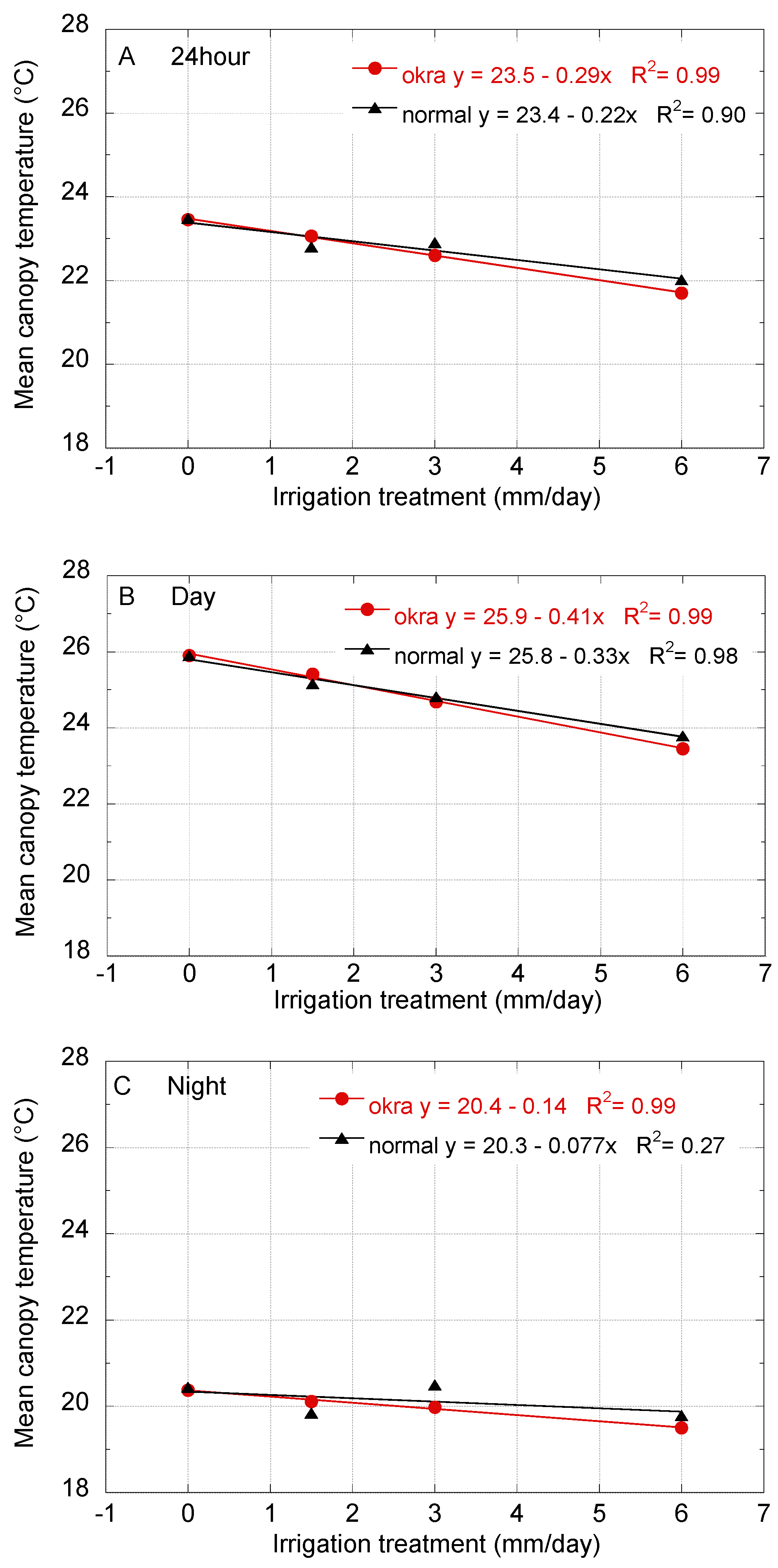
Figure 4 shows the mean canopy temperature for the normal and okra leaf cottons across the water levels when sorted into 24-h, day, and night groupings. The effect of irrigation on the canopy temperature is apparent and most pronounced in the daytime canopy temperatures. The mean canopy temperatures, with the exception of the night period are highly correlated (
R2 > 0.9) with irrigation treatment. The canopy temperatures at the 0 mm irrigation level are most similar. Canopy temperatures in the okra variety were higher than the normal variety at the 1.5 mm irrigation, and the okra was lower than the normal variety at the 3 mm and 6 mm levels. The trend is consistent across the 24 h, day, and night intervals.
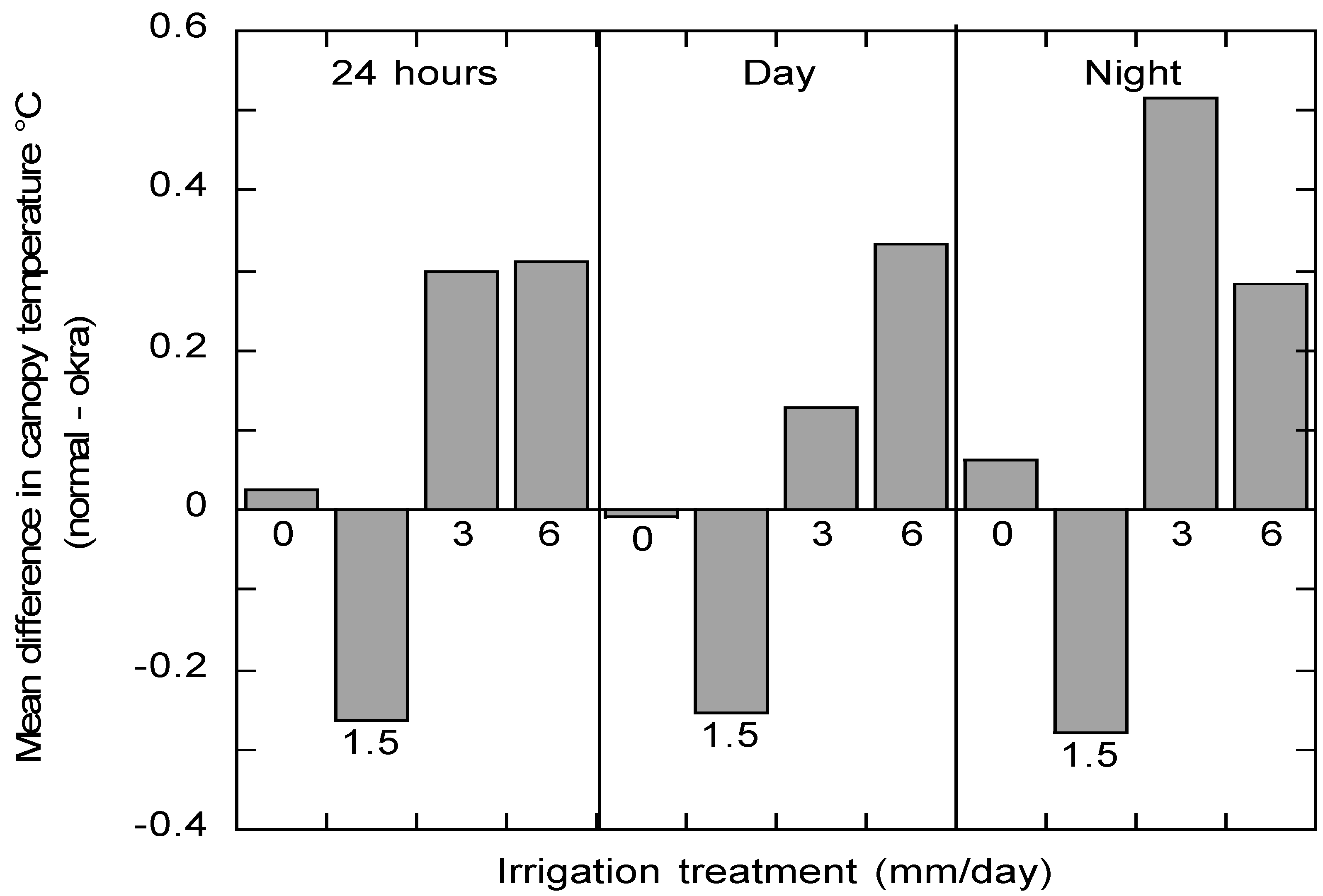
Figure 5 shows the seasonal mean of differences between the normal and okra for the 120-day measurement period as a function of irrigation treatment and time of day. Positive values indicate normal leaves are warmer and negative values indicate okra leaves are warmer. At the 0 mm irrigation level, the canopy temperatures of the two leaf types are very similar while in the 1.5 mm irrigation treatment the okra leaves were warmer than the normal leaves. At the 3 mm and 6 mm irrigation levels the normal leaves are warmer than the okra leaves. The pattern is consistent between the night and day periods.
The analysis of canopy temperatures across several multi-day periods of the season underscores the well-established effect of irrigation on canopy temperature [
1,
2]. While differences in canopy temperature between the two varieties are evident and often significant, they are generally small in magnitude. Viewing the canopy temperatures on a daily basis as a time series may provide additional insight into differences between the varieties.
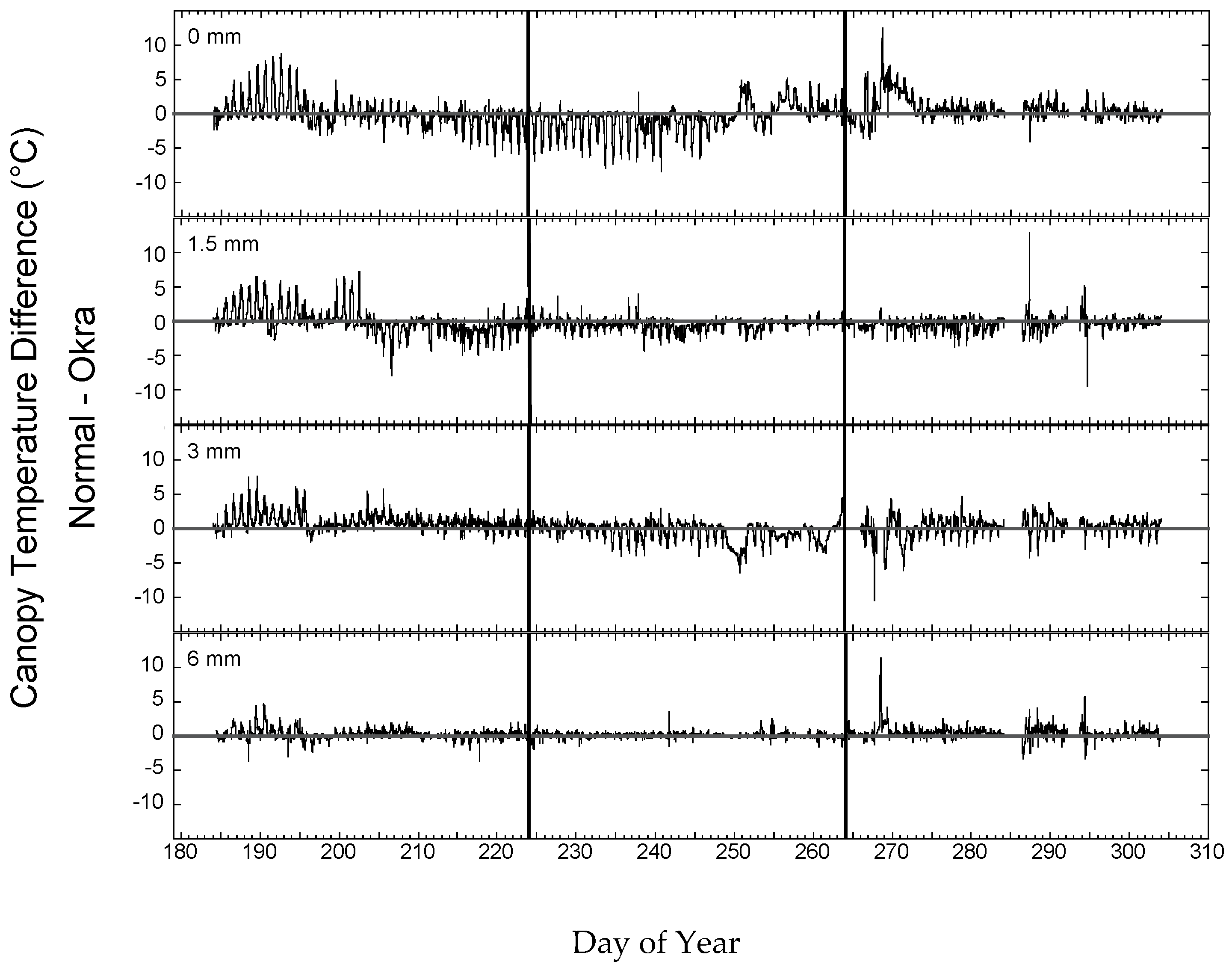
Figure 6 shows the difference in canopy temperature (normal leaf–okra leaf) over the 120-day Full-season period. The effect of irrigation on the differences is evident with the 6 mm irrigation treatment consistently showing the smallest differences, and the rainfed treatment showing the largest differences between the two leaf shapes. The two intermediate irrigation regimes showed temperature differences between the irrigation extremes. The patterns of differences over the season were generally consistent in the 6 mm irrigation while the three other irrigation treatments all showed a seasonal pattern with the normal leaf warmer (difference values positive) early in the season. The rainfed treatment showed the largest differences between the two leaf types with the normal leaf warmer in the early period, cooler in the middle period and warmer later in the season.
The distribution of the differences over the full-season period helps to understand how the differences in seasonal means between the two varieties across water levels are relatively small (<0.5 °C) while, for any point in time, differences >5 °C are not uncommon. Irrigation consistently reduces the differences between the varieties as evidenced in the pattern of variance in
Table 1.
Since the multi-day means reduce the variation apparent in seasonal differences, it might be instructive to look at the time series of canopy temperature data. While the effect of irrigation on canopy temperature is apparent, not all days of the season look the same (see
Figure 6).
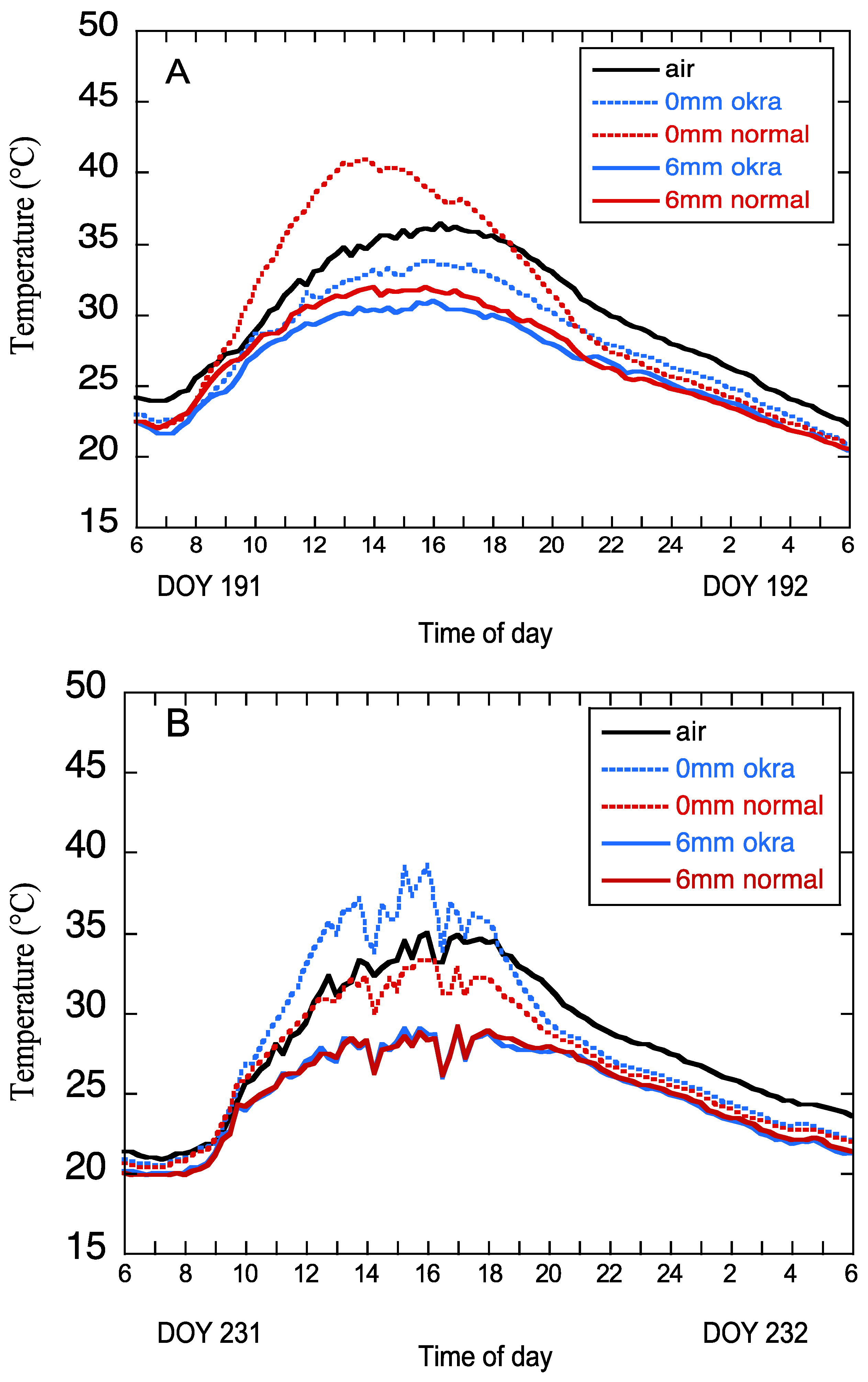
Figure 7 shows the pattern of air and canopy temperature for two 24-h periods (DOY 191 and DOY 231) of the study period that were chosen to demonstrate some characteristics of canopy temperature in the two varieties at two water levels.
Figure 7A shows the patterns of air and canopy temperature for DOY 191. At 6 a.m. the temperatures of the varieties are quite similar, the water effect is minimal and canopy temperatures are below air temperature. As radiation increases over time, both air and canopy temperatures increase. The canopy temperatures of both varieties in the 6 mm irrigation treatment are similar throughout the period. In the 0 mm treatment, the temperatures of the varieties begin to diverge by 8 a.m., with the normal leaf variety becoming warmer than the okra leaf and remaining higher until approximately 8 p.m. The temperature of the normal leaf variety remains lower than the okra over the 8 a.m. to 8 p.m. period. After 8 p.m. the temperatures of the varieties remain similar and below air temperature for the remainder of the 24-h period. On this day canopy temperatures for both varieties in the 6 mm treatment and the normal leaf in the 0 mm treatment remained below the air temperature over the period. Under such conditions, transpirational cooling largely influences the canopy temperatures, and leaf shape-related differences would be minimal. During DOY 191, any effect of leaf shape on canopy temperature would probably be relevant only for the normal leaf in the 0 mm treatment.
Figure 7B shows air and canopy temperatures for DOY 231. At 6 a.m. the temperatures of the varieties are quite similar and the water effect is minimal. Canopy temperatures are below the air temperatures. As radiation increases over time, both air and canopy temperatures increase. In the 6 mm irrigation treatment, the temperatures of the varieties increase and remain very similar. In the 0 mm treatment, the temperatures of the varieties begin to diverge by 8 a.m., with the okra leaf variety becoming warmer than the normal leaf and remaining higher until approximately 8 p.m. The temperature of the normal leaf variety remains lower than the okra over the 8 a.m. to 8 p.m. period. After 8 p.m. the temperatures of the varieties remain similar and below the air temperature for the remainder of the 24-h period. On this day canopy temperatures for both varieties in the 6 mm treatment and the normal leaf in the 0 mm treatment remained below the air temperature over the period. Under such conditions, transpirational cooling largely influences canopy temperatures and leaf shape-related differences would be minimal. During DOY 231, any effect of leaf shape on canopy temperature would probably be relevant only for the okra leaf in the 0 mm treatment.
The analysis of two days of temperature variation demonstrates that the canopy temperature differences between the two varieties change over time. In each of the single days subjected to analysis, the 6 mm irrigation level produced similar canopy temperatures patterns. At the 0 mm irrigation level, the temperatures of the varieties differed over the course of the day with the normal variety warmer than the okra variety on DOY 191, with the reverse being true on DOY 231.
Since the difference in canopy temperature between the two leaf shapes is variable over the season, the distribution of those differences over time might be instructive.
Figure 8 shows the distribution of the difference between normal and okra leaf canopy temperatures over the study period. The distributions for the 3 mm and 6 mm treatments are symmetrical with a peak at 0 °C though the 3 mm distribution is slightly broader. The peak in the 1.5 mm distribution is at −1 °C indicating a shift toward the okra leaf being warmer. The distribution of the 0 mm is broader and generally shifted toward okra leaf being warmer. These data indicate that at high irrigation the differences between the okra and normal leaf varieties small and random while, at the lower irrigation levels, the differences are larger and skewed toward the okra leaf being slightly warmer than the normal leaf. These results suggest that the okra leaf canopy temperatures are not lower than the normal leaf temperatures based on the possibility of enhanced sensible energy exchanges associated with the okra leaf shape, particularly in the lower irrigation regimes.
The distribution of canopy temperature differences shown in
Figure 8 suggests that canopy temperatures in the okra leaf are not cooler than those of the normal leaf under water deficits. To further investigate this finding, the canopy temperature data has been analyzed from the perspective of conditions that enhance or restrict sensible energy exchanges between the canopy and the environment. Leaf shape differences in sensible energy exchanges would be enhanced when canopy temperature is greater than air temperature, generally under low irrigation. The
Figure 9 [
1,
2] compares the canopy temperatures of the okra leaf to the normal leaf as a function of the difference between the okra leaf and air temperatures. On the x-axis, as the difference between okra leaf canopy and air temperature increases, the ability of the canopy to cool by sensible energy exchanges increases (left to right). On the y-axis, positive values of the difference between the normal and okra leaf canopy temperatures indicate that the okra leaf is cooler than the normal leaf. Temperatures in the lower right quadrant indicate that the normal leaf is cooler than the okra. Enhanced energy exchange caused by the okra leaf shape would be indicated by canopy temperatures in the upper right quadrant of the figure. Under well-irrigated conditions (6 mm), it would be expected that latent energy exchanges would predominate and canopy temperatures would be similar between leaf shapes and largely independent of air temperatures. The data agree. Under water deficit conditions (0 mm), when latent energy exchanges are limited, enhanced sensible exchanges by the okra leaf would be indicated by temperature values in the upper right quadrant. The predominance of cooler normal leaf values in the lower right quadrant suggest better cooling of the normal compared to the okra under conditions where sensible energy exchanges should predominate. This evidence suggests that canopy temperature differences between the varieties are not a result of enhanced sensible energy exchanges in the okra leaf variety.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}