1. Introduction
Onion (
Allium cepa L.), of the Liliaceae family, is a species of worldwide economic importance [
1]. Under appropriate photoperiod and temperature regimes, onion leaves progressively change from photosynthetic to storage units [
2]. Onions are characterized as edible bulbs rich in sugar and pungent oil that gives them distinctive flavors [
3]. Bulbing is mostly considered as a carbohydrate accumulation [
4] and is easily observable as basal swelling [
5]. Since photosynthesis and water loss through transpiration share the same pathway, physiological measurements commonly include estimation of photosynthesis, stomatal conductance, and leaf internal CO
2 concentration, thus providing mechanistic insights into variation in growth among varieties and the growing conditions.
California is the USA’s largest producer of onions, with over 26% of the nation’s total production and generating about
$206 million revenue in 2010 [
6]. While there are many onion varieties, they can be generally classified by daylength sensitivity and bulb color. The sensitivity of plants to daylength, or photoperiodism [
3], is crucial for designating planting schedules. Several researchers suggested that, if long day promote bulb formation, it should be related to photosynthesis [
7,
8]. The researchers also stated that a daylength of 6 h decreased daily photosynthetic rates by decreasing the time for carbon uptake. Further, cells grown with this daylength had maximal chlorophyll contents, suggesting a physiological adaptation by photosynthetic units to short light periods. A long-day (LD) photoperiod of 18.6 h at 25 °C decreased daily rates of net photosynthesis by reducing the hourly rate of net photosynthesis via an unidentified mechanism [
9]. The importance of rates of net dark respiration in controlling daily net photosynthesis was small, with carbon lost during dark periods varying between 4% and 14% of that gained during light periods. In addition, the influence of net excretion was small, varying between 1.0 %and 5.5% of daily net photosynthesis [
9]. These observations suggest that there exist controversies on effects of daylengths on crop photosynthesis.
Short-day onion varieties bulb at 10–11 h of daylength, while the long day onions require greater than 12 h for bulbing. Most California varieties begin to bulb at 12–15-h daylengths. Therefore, onion varieties and planting dates are extremely important for proper bulb development. Growers would also need to know conditions that may enhance plant growth and bulb formation. With such knowledge, growers can match planting dates with the onset of longer daylight and temperature in order to maximize bulb formations. Although this information is important to the onion growers, little or no research has been conducted to document the combined effects of daylength and temperature, particularly for growing onions in the high desert regions of California.
The objective of this experiment was to evaluate the effect of photoperiod length and temperature regimes on the physiological activities of various onion varieties commonly grown in the high desert regions and elsewhere. The facts of developmental plant physiology clearly show that the course of development can be altered by environmental factors. With the onions, changes in light and temperature factors often express themselves in organ formation and growth.
2. Materials and Methods
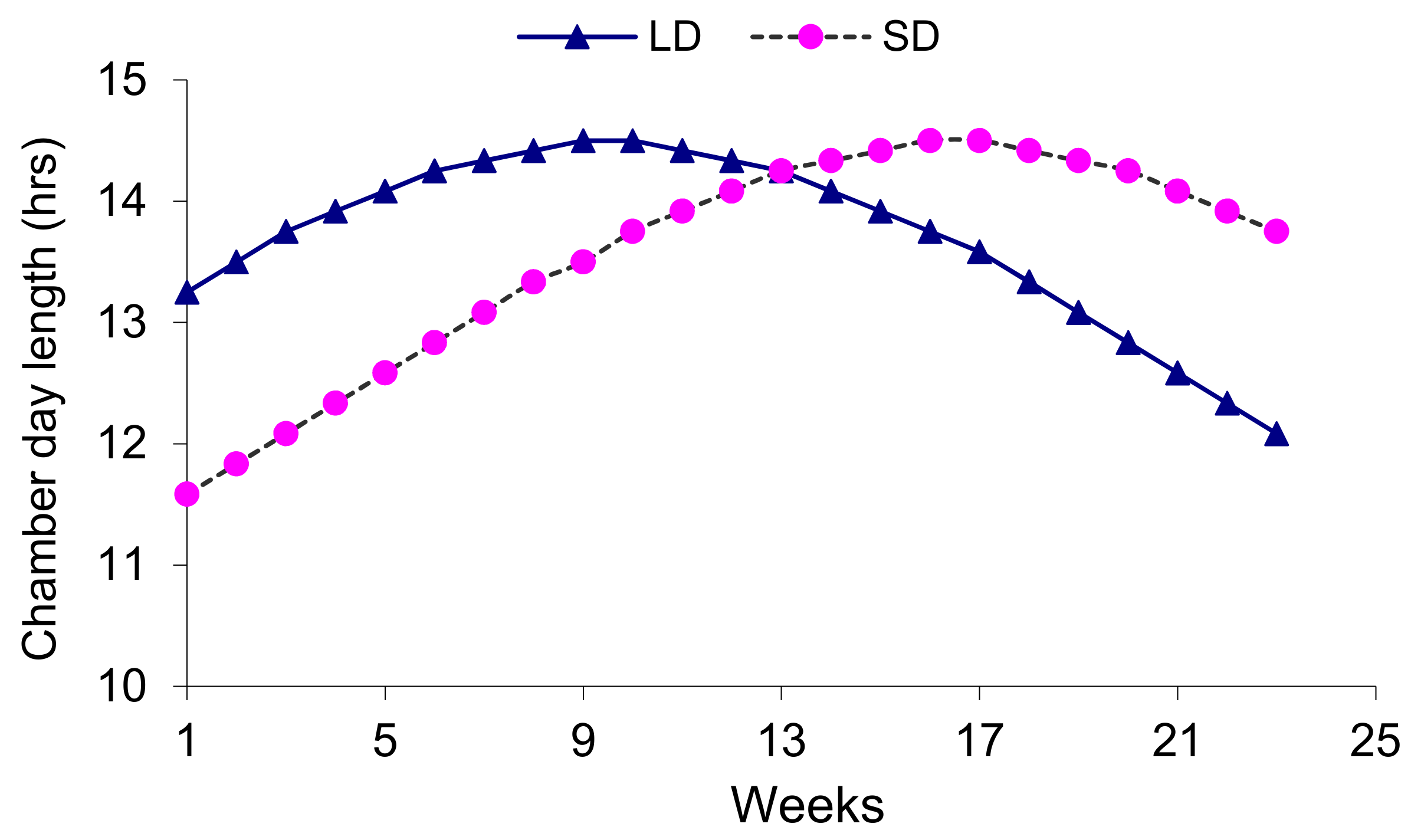
Six onion varieties were grown in two sets of controlled growth chambers, namely the short-day (SD) and long-day (LD) chambers. Each chamber was calibrated with the short and long daylengths (
Figure 1) settings in order to simulate the early and late planting seasons, respectively, common to the Antelope valley of California’s Mojave Desert, where onions are largely grown.
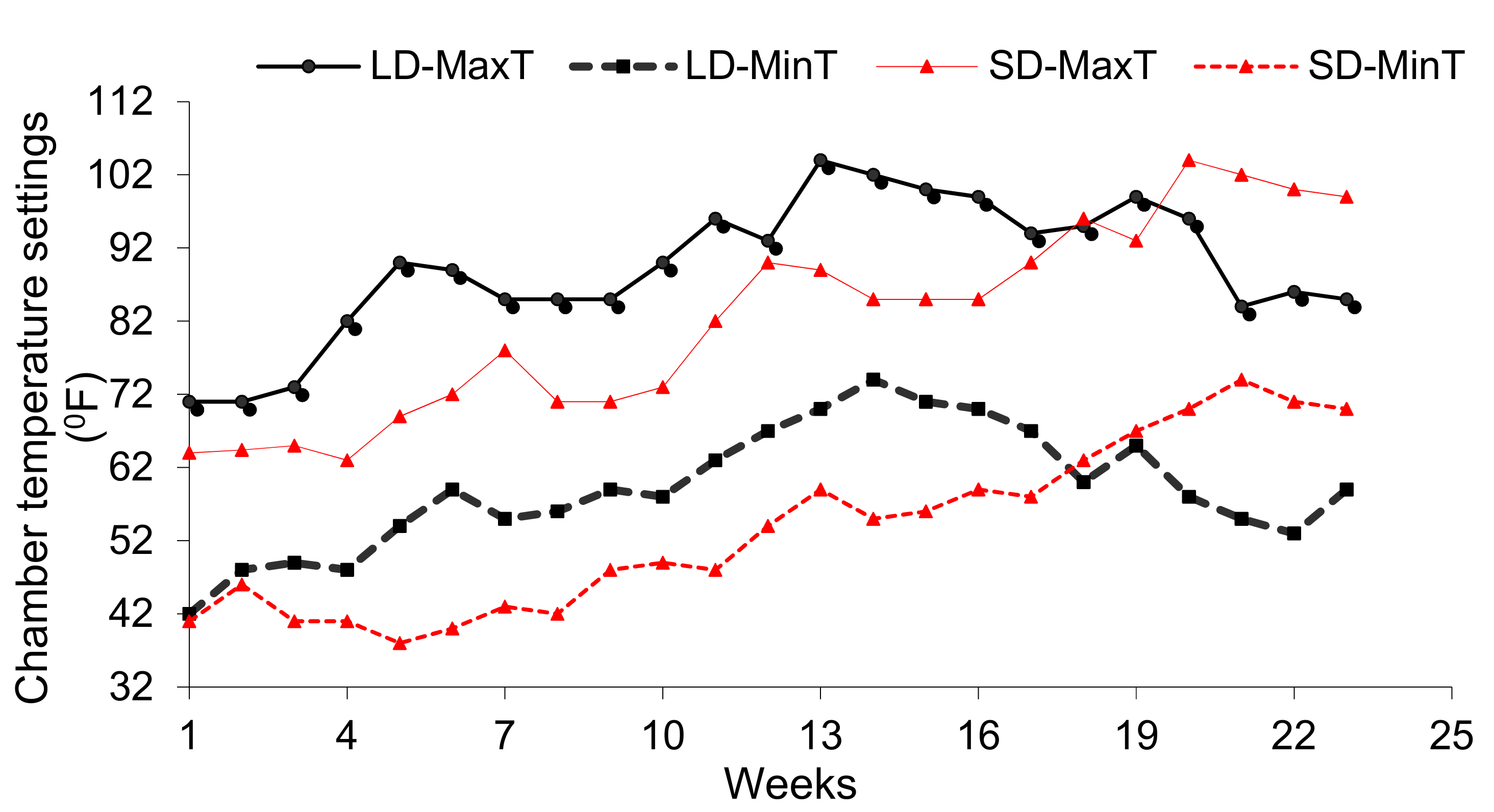
The associated maximum and minimum temperature setting for each growth chamber (
Figure 2) mimicked the actual onion growing field temperatures of the region during the early and late planting seasons. Daylength and temperature settings for the chamber were calibrated based on 15-year average weather data for Lancaster, CA, a high desert onion production site in the Antelope valley. Long-term climate data associated with the source of the simulated daylength and temperature settings are available from California Irrigation Management Information System (CIMIS) (
http://www.cimis.com).
The daylength and temperature settings of both growth chambers during which the physiological plant data were collected are shown in
Table 1. Physiological plant date were measured before midday, on well-watered plants, such that midday stomatal closure and water deficit did not reduce the rates of photosynthesis. The short-day growth chamber represents the early planting season of the high desert region, which is normally the first week of March. The long-day chamber represents farmers late planting of 20 April.
Six varieties of onion, namely RedBull (RB), RedWing (RW), Hybrid Cometa (HC), Granero (YG), Hybrid Valero (HV) and Hybrid SR (HS), we selected. All varieties, except HS, are known to be long-day varieties. The varieties were selected for testing because they all are grown in the Antelope valley of California’s Mojave Desert, where onions are largely grown. Each of the varieties were planted from seeds in a shallow depression in 20 L pots filled with UC#3 potting medium. Into each pot was planted 10 seeds. After germination, seedlings were thinned to three plants per pot. Each of the six varieties were replicated four times within each chamber and laid out in a completely randomized design. Plants were supplied with uniform light intensity (irradiance levels) of 280 µmol m−2 s−1 at the leaf surface with fluorescent light bulbs. A complete nutrient solution was applied on a weekly basis and plants were watered every other day.
Leaf physiological parameters (photosynthesis, stomatal conductance and leaf internal CO
2 concentration) were measured in-situ using a portable photosynthesis system equipped with an infrared gas analyzer (model LI-6400; Li-Cor Biosciences, Inc. Lincoln, NE, USA). Following photosynthetic measurements, leaf area of the plants was measured using a leaf area meter (LI-3100C; Li-Cor Biosciences, Inc., Lincoln, NE, USA). At the time of physiological data collection, plants grown in the short-day chamber were at the bulbing stage, whereas plants in the long-day chamber were at the post-bulbing stage. Leaf photosynthetic rates, stomatal conductance and internal CO
2 concentration variations were analyzed using Analysis of Variance (ANOVA) and a
t-test for mean comparisons (SAS 9.1 statistical systems, SAS Institute Inc., Cary, NC, USA). Bivariate relationships testing for differences in slopes and elevation (
y-intercept) among chamber growth conditions for relationships between maximum photosynthetic CO
2 assimilation per leaf area (
Aarea), stomatal conductance to water vapor (
gs), and internal CO
2 concentration (
Ci) were analyzed using standardized major axis estimation (model II regression) with (S)MATR ver 1.0 software [
10,
11], following analysis procedures used by Santiago and Wright [
12]. The program first tested for differences in slope between chamber conditions using likelihood ratios. If no significant difference in slope was detected, tests for differences in elevation (
y-intercept) and whether chamber conditions were separated along the standardized major axis with a common slope were performed using randomization routines that are analogous to ANCOVA.
3. Results
There were differences in leaf photosynthetic rates and stomatal conductance of onion varieties based on the daylength settings of the chambers, although there was no significant difference between their internal leaf CO
2 concentrations,
Ci (
Table 2). The findings suggest that onion leaf photosynthetic rates and stomatal conductance may be more sensitive to environmental conditions than other leaf physiological traits.
Within the current daylight and temperature settings (
Figure 1 and
Figure 2), four out of six onion varieties grown in the short-day (SD) chamber had higher leaf photosynthesis than the respective varieties grown in the long-day (LD) chamber (
Table 2). The differences in leaf photosynthetic rates of short-day versus long-day grown plants ranged between 49% for the HS and 64% for the HC varieties. Two of the six varieties (RW and HV) grown in the short-day condition had higher stomatal conductance than the respective varieties grown under long-day conditions (
Table 2).
Similarly, the variation between leaf stomatal conductance of the short-day and long-day plants was 40% for the HS and 58% for the RW varieties (
Table 2). These results also suggest that there are variations among onion varieties in their responses to onion growing season environmental conditions. For example, the photosynthetic and stomatal conductance of the HS variety showed less variation between the short-day and long-day growing conditions compared to the other five varieties (
Table 2). The observed differences among onion varieties grown under different chamber environmental conditions reveal that onion leaf functional traits may also vary under natural conditions, if the planting dates were to take place at different periods of onion growing seasons.
Although the long-day plants appeared to be more vigorous than their short-day counterparts (data not shown) [
13], plants grown in the short-day chamber appeared to be physiologically more active than these grown in the long-day chamber (
Table 2). The comparative advantages of the short-day plants may have been due to the higher weekly daylength and temperature levels that the short-day chamber plants were receiving during the current crop growth phase, revealing that physiological activities of onion plants are enhanced by the length of daylight and the degree of temperature. Since short-day plants were also at the initiation of bulbing during these environmental conditions, it seems that longer daylength and temperature might have been vital in triggering onion bulbing. Bulbing in turn demands higher photosynthetic rate to balance the need for additional storage, which in this case is in the form of bulbing. Steer [
14] showed that cultivars grown at 13 h daylength and 26 °C day temperature bulbed faster than those exposed to lower daylength and temperature levels. Heath and Hollies [
15] suggested that basal swelling was due to the increases in easily assimilated sugars in the culture medium, but not to the enhancement of osmotic pressure.
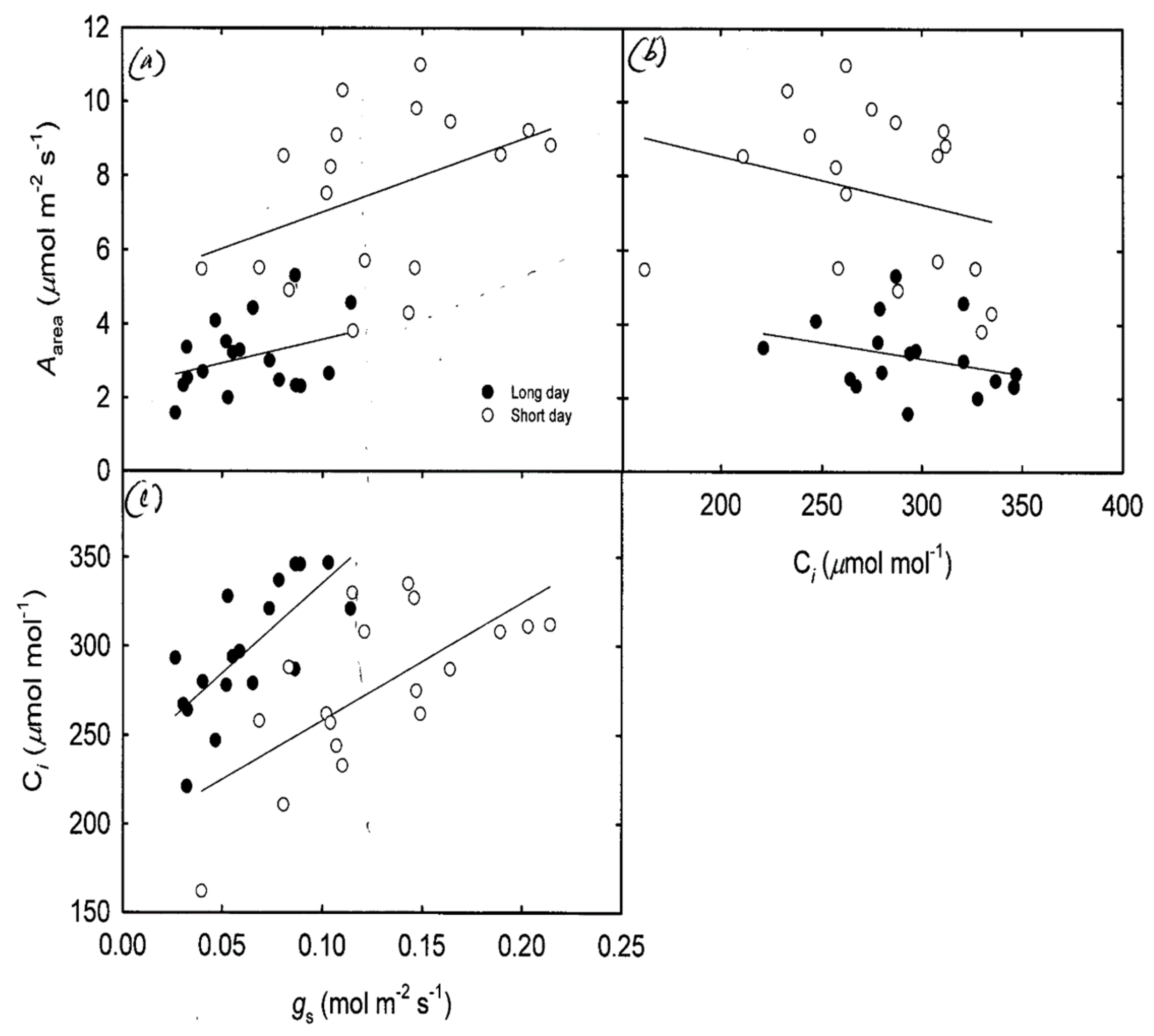
There was a positive linear relationship between the
gs and
Aarea of onion varieties (
Figure 3a). However, the slopes of varieties grown under short-day and long-day conditions showed no significant difference (
Figure 3a,
p = 0.480). However, there was a significant difference in
y-intercept (
Figure 3a,
p = 0.009), indicating a significant shift in elevation of the mean values between crops grown under long-day and short-day conditions. The relationship reveals greater leaf photosynthetic activity of short-day plants relative to the long-day plants for a given stomatal conductance of each of the onion varieties. There was a negative linear relationship between
Ci and
Aarea, but there was no significant difference between the slopes of plants under contrasting chamber conditions (
Figure 3b,
p = 0.104). There was a significant difference in
y-intercept (
Figure 3b,
p < 0.001), indicating that plants in the short-day chamber conditions had a higher
Aarea per unit
Ci than the long day plants. There was a positive relationship between
gs and
Ci (
Figure 3c) with no significant difference in slopes between groups (
Figure 3c,
p = 0.137). There was a significant difference in
y-intercept (
Figure 3c), indicating that plants in the long day chamber conditions had a greater
Ci per unit
gs than the short-day plants. The higher
Ci per unit
gs in long-day plants may be associated to the crops lower photosynthetic activity when grown in a long-day chamber.
4. Discussion
As seen from the results, onion varieties varied in their physiological responses to environmental conditions at their bulb setting stage. However, not all onion physiological responses are sensitive to variation in environmental conditions. While leaf photosynthetic rates and leaf stomatal conductance of all tested onion varieties were affected by crop growth conditions, the internal CO2 concentrations of the varieties were not affected, indicating that onion photosynthetic rates and stomatal conductance are more sensitive to environmental conditions than some other physiological conditions. Our results also indicate that most tested varieties had enhanced leaf photosynthetic activity and higher leaf stomatal conductance if shorter daylength and higher temperature conditions coincided with the onset of onion bulbing stages. The increased physiological activities of onions depending on daylength exposures reveals the importance of selecting onion planting dates so that onion bulbing coincides with crop physiological enhancing weather conditions.
The elevated shift in mean leaf photosynthetic rate of short-day plants per given leaf stomatal conductance and leaf internal CO
2 concentration (
Figure 3) indicates a substantial change of a leaf functional trait in response to growth stages. Furthermore, the later growing stage under short daylength (post 13 weeks of growing) and higher temperature settings in the short-day chamber (
Figure 1) might have hastened crop bulb initiation. It may also be true that bulb formation created a strong photosynthetic sink and hence demanded higher leaf physiological activity to fulfill the storage demands of onion bulbs. The latter may be more relevant as the short-day plants were at the bulb initiation stage, thus having a greater demand for photosynthates than the post-bulbing long-day plants, hence a relatively lower sink strength.
The short-day plants that received long-daylengths and higher temperature had a comparative advantage in their leaf physiological activities over those that received short daylength and lower temperature during their initial growing seasons. These contrasting physiological variations at different growth stages of onion varieties can exhibit variations in physiological traits to growing conditions and how photosynthetic resources are deployed into the bulb (storage), which may then result in variations of onion yield, as can be measured by bulb size and quality. However, there were differences among onion varieties as to the degree of responses to variation in onion growing environmental conditions. For example, the HS variety responded at relatively lower variation of leaf physiological activities to varying degree of environmental conditions (
Table 2) than the other varieties.
One of the most remarkable patterns revealed in our results was that short-day onions exhibited a lower internal leaf CO
2 concentration per given leaf stomatal conductance, even though these plants showed a larger range of stomatal conductance and similar slope ranges (
Figure 3c). This lower internal leaf CO
2 concentration may be attributed to the faster CO
2 assimilation of the plants grown in the short-day chamber, caused by higher metabolic activities when exposed to short daylength and higher temperatures at the time of bulb settings. Elevated CO
2 increases crop yields [
16], primarily because of higher rates of net photosynthesis [
17]. However, Long [
18] stated that it is the increase in photorespiration that reduces the rate of net photosynthesis at warmer temperatures. Similarly, Denne and Smith [
19] observed a decreased rate of CO
2 uptake in
Picea sitchensis under long-day compared to the short-day conditions. These rates may be controlled by temperature and photoperiod, which also affects rates of uptake and loss of carbon from cells. Hobson et al. [
9] stated that the increased daily uptake rate is due to the longer time for photosynthesis and not to an increased rate of photosynthesis. These discrepancies could be due to the variation in the potential responses of plant species to varying daylength and temperature levels of a growing season. The wider range of leaf stomatal conductance for the short-day plants, relative to these grown in the long-day chamber, may mean that the higher leaf stomatal conductance does not necessarily result in a higher leaf photosynthesis. It may also mean that the daylength and temperature levels must be coupled with a higher leaf conductance to result in a higher photosynthetic capability of onion leaves.
With the settings of our current experimentation, it is not easy to differentiate whether the physiological responses in onion varieties were attributed to the length of the day, temperature levels or the combination of the two factors because our long-day treatment was characterized by initial longer daylength and higher temperature while the short-day plants were grown under initial short daylength and lower temperature ranges (
Figure 1). Steer [
14] observed a significant interaction of cultivar, daylength and temperature on onion bulbing, in which case bulbing was most rapid at the highest temperature and longest daylength. Therefore, the interaction between photoperiod and temperature is so important that the minimum photoperiod for a cultivar should never be specified without the corresponding temperature specification.
In general, higher temperatures accelerate, and lower temperatures may delay bulb development, which could be accompanied by changes in physiological activity of onion plants. The slower physiological activities of plants in the long-day chamber compared to plants grown in the short-day chamber have to do with the timing of bulbing. According to Wheleer et al. [
20], acclimation of the rate of light-saturated photosynthesis was apparent at the time of onion bulbing, perhaps due to the onion’s clearly defined source and sink organs at the time of bulbing. At post bulbing, the leaves transfer from a photosynthetic organ into a storage structure (bulb) after bulb initiation, reducing the plant’s photosynthetic activity. The leaf traits and relationships to the environmental conditions and plant growth stages may reflect resource allocation differences to maximize carbon gain of storage parts of a plant. The fact that bulbing is mostly considered as a carbohydrate accumulation, as easily observable through basal swelling of the most external sheaths of the plant, was also suggested by Lercari [
4]. Increased source activity relative to sink activity may then increase carbohydrates in the plant and, depending on the ability of onions to store these carbohydrates, may lead into a higher yield.
Since variation in our leaf functional traits among growth conditions were driven from the onion growing conditions of high desert conditions and that our crop growth chambers were calibrated to mimic the late and early planting dates of the high desert farmers, our findings can be utilized by high desert onion farmers. Growers can choose best planting seasons as to match plant bulbing stages with the onset of long-days and higher temperature ranges, because longer daylength and higher temperatures at this growth stages of onions may further enhance bulb formation. Matching growing environments with potential plant growth stages could be advantageous to onion growers for early and increased bulbing and bulb sizes. The coincidence of favorable growing conditions with the bulbing stage could enhance the speed at which nutrients may be partitioned into the bulbs. Therefore, matching plant growth and development with favorable conditions could ultimately enhance yield and profitability of onion crops.
In summary, onion plants strongly respond to growing daylength and temperature ranges of onion growing regions and seasons. The increased physiological activities as measured from onion crop photosynthetic and stomatal conductance explains the potential reaction to daylengths and temperature regimes. Since the long-day and short-day growth chambers were calibrated to mimic the late and early planting dates of the farmers, the results can be replicated by onion farmers, at least for those located within the high desert areas of California and similar regions. All varieties other than HS are well suited for late planting, indicating that the former varieties are long-day varieties and the latter a potential short-day variety. The late planting days (simulated by the long-day chamber) are exposed to higher growing daylength and temperature during their early growing stages. Accordingly, they could bulb early and hence maximize profitability by producing vigorous bulbs providing early harvesting.






{kind=link}
{kind=link}
{kind=link}