Trophic and Microbial Patterns in the Ross Sea Area (Antarctica): Spatial Variability during the Summer Season

 ,
,  ,
,  , , ,
, , ,  ,
,  ,
, 
Abstract
:1. Introduction
- (i)
- to perform a quali-quantitative assessment of the trophic conditions of the water column along coastal to offshore and vertical profiles covering from deep to surface layers in terms of particulate and dissolved organic matter and of the CDOM component in two different areas of the Ross Sea;
- (ii)
- to depict the distribution patterns of microbial prokaryotic and eukaryotic communities and their main components (living, metabolically active, culturable cells)
- (iii)
- to highlight the interactions of the microbial assemblage with available organic substrates
- (iv)
- to find the main variables affecting the microbial community dynamics and the response of these latter to the different environmental scenarios.
2. Materials and Methods
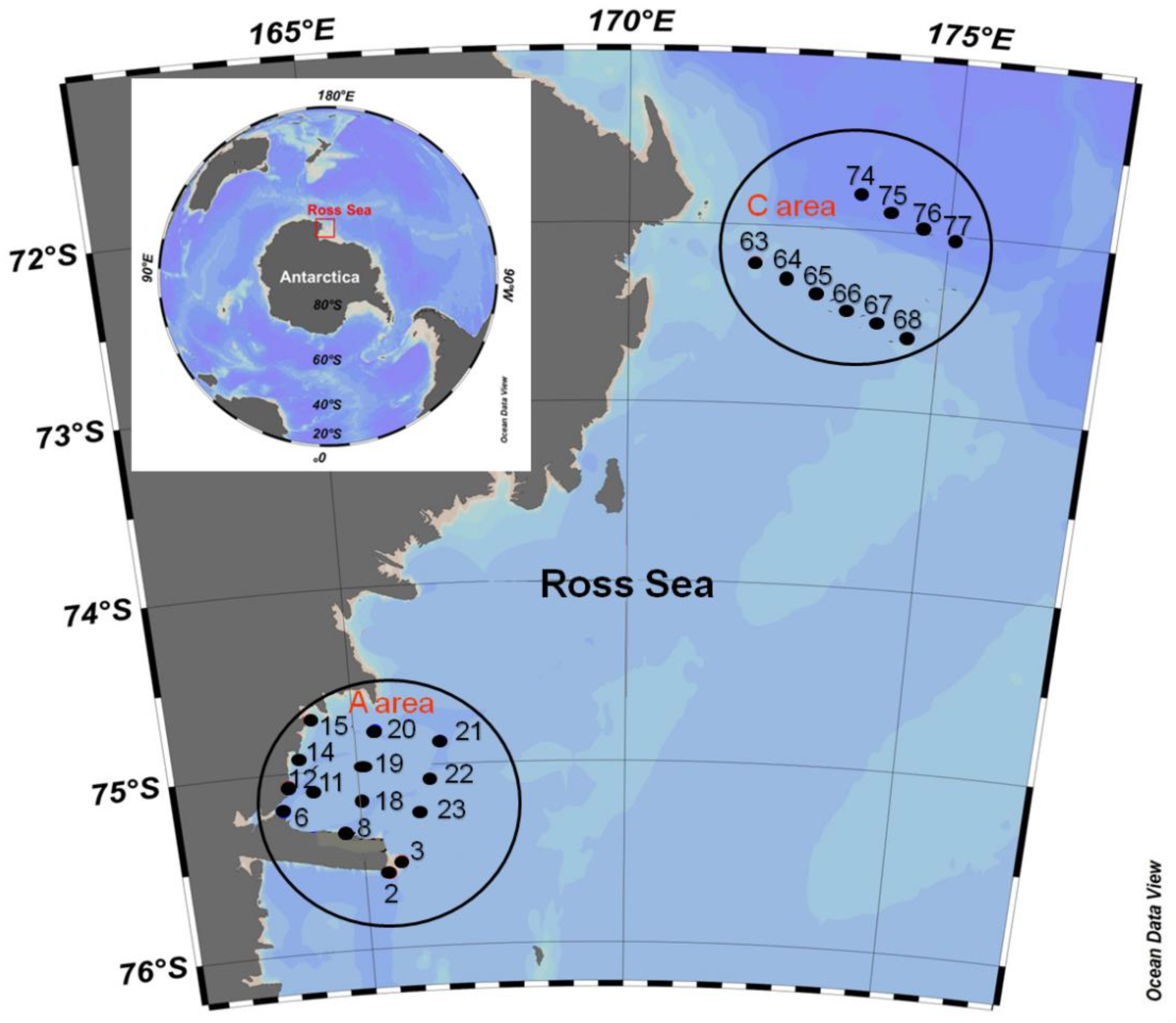
2.1. Description of the Area and Sampling Strategy
2.2. Measured Variables
2.2.1. Hydrological, Trophic and Optical Variables
2.2.2. Microbial Abundance Data
Total Prokaryotic Abundance, Volume and Biomass
Microbial Assemblage Components
Prokaryotic, High and Low Nucleic Acid Content, Pico- and Nano-Eukaryotic Cell Abundance by Flow Cytometry
2.2.3. Microbial Metabolism
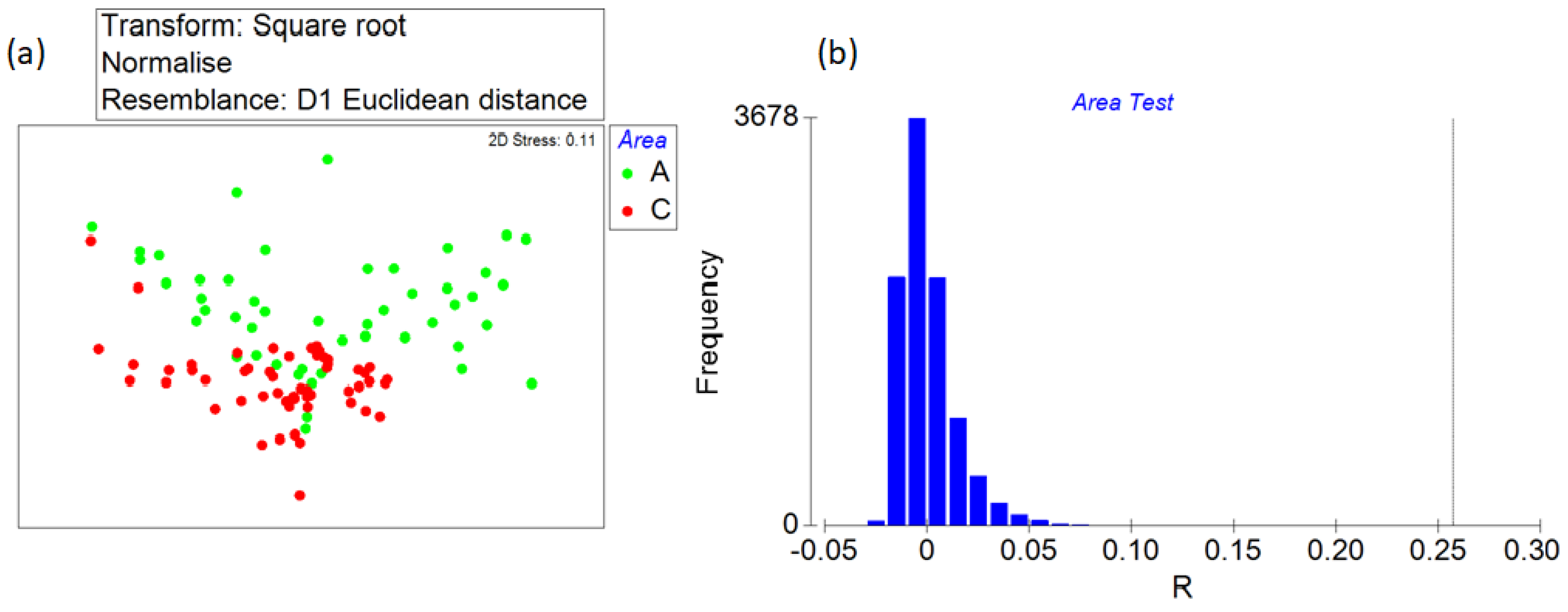
2.3. Data Analysis
3. Results
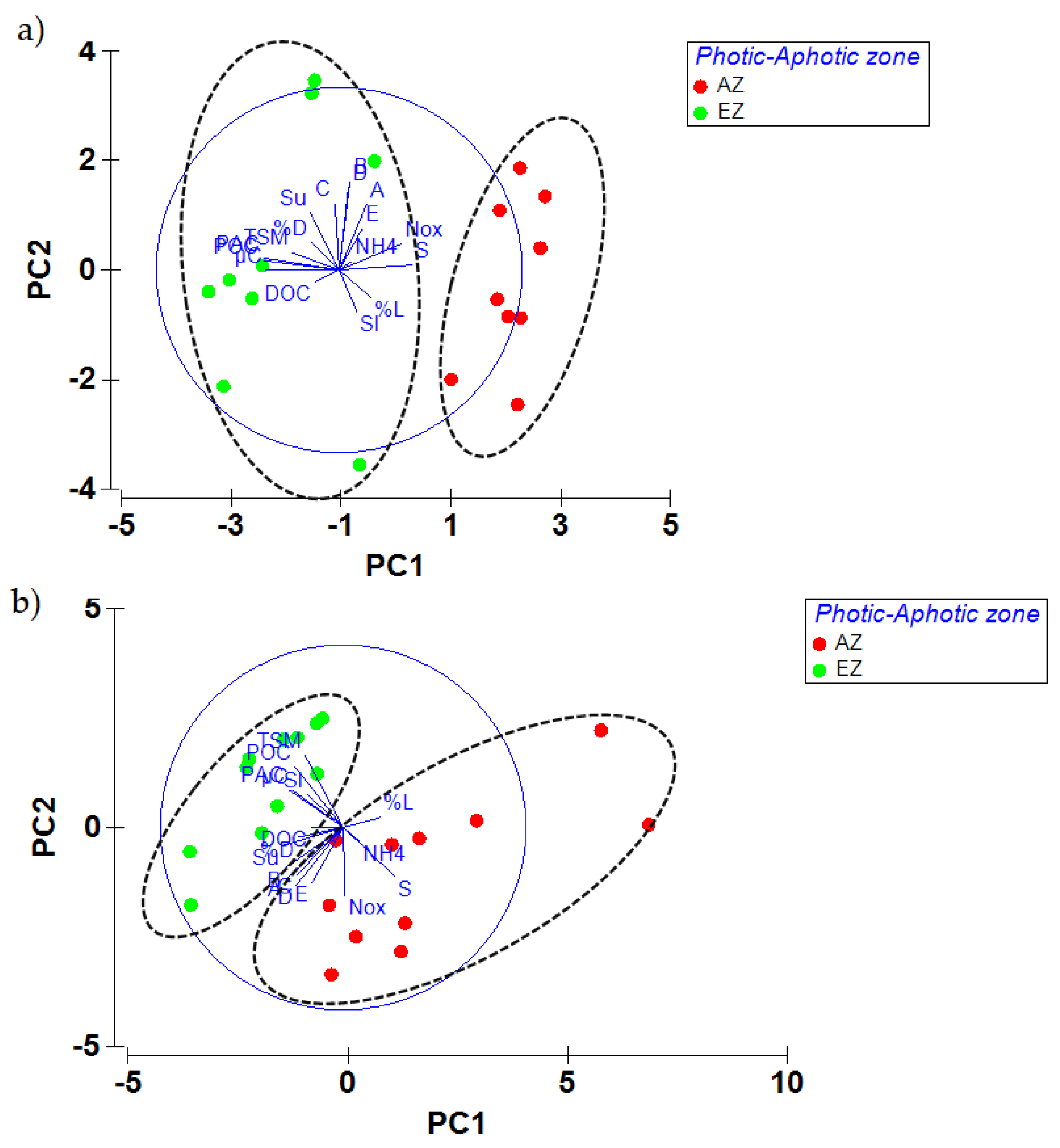
3.1. Significant Differences between the Two Areas
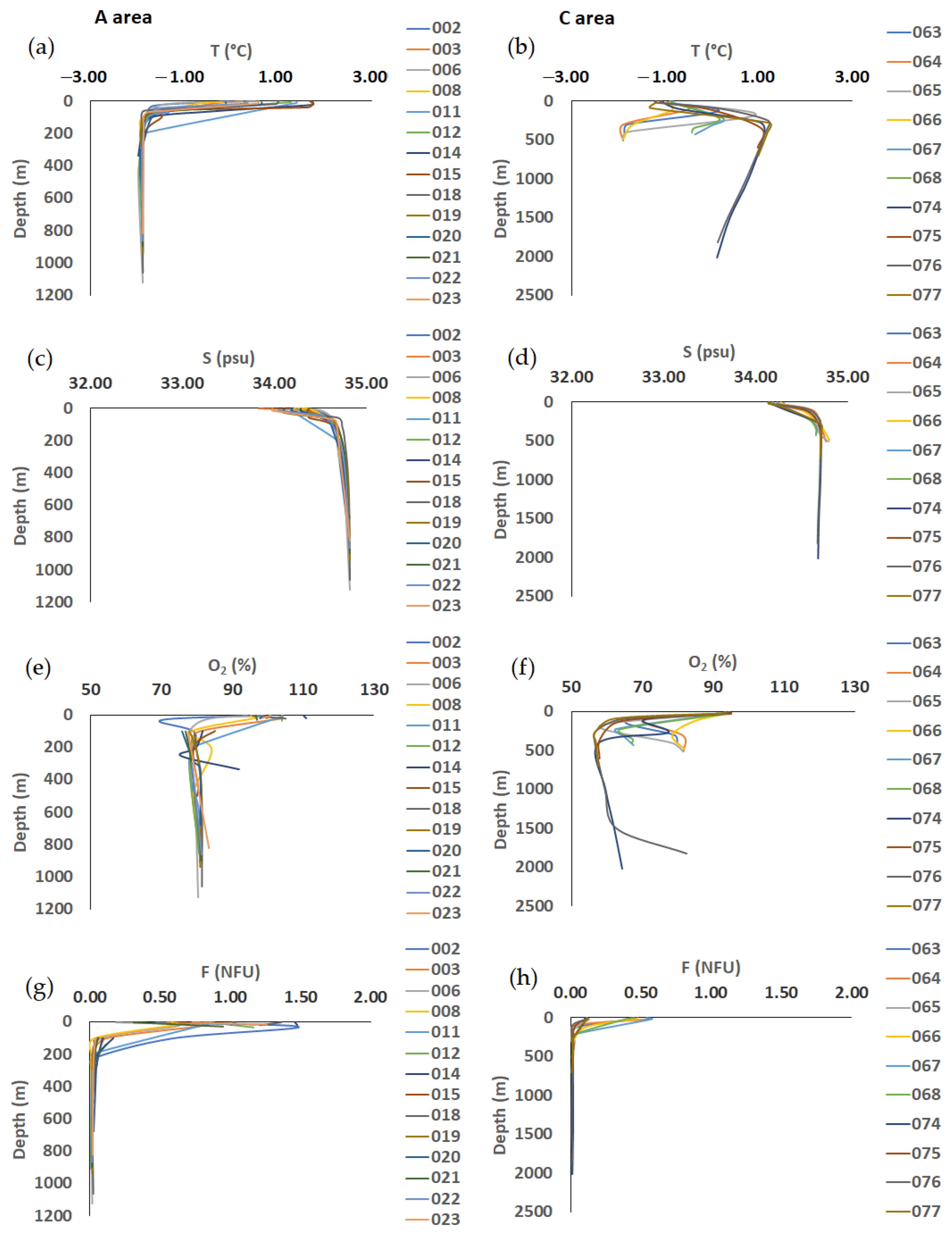
3.2. Hydrological Properties
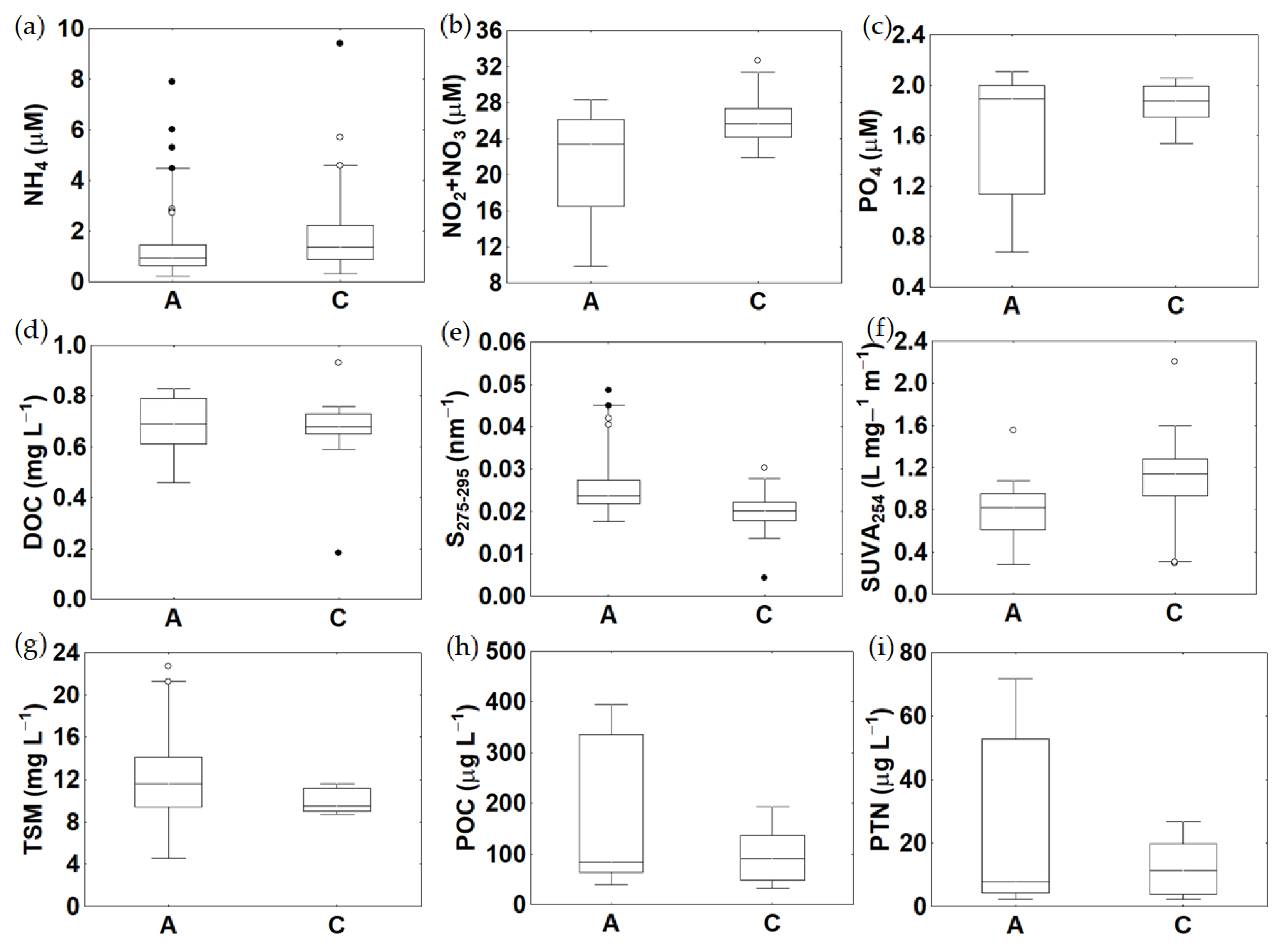
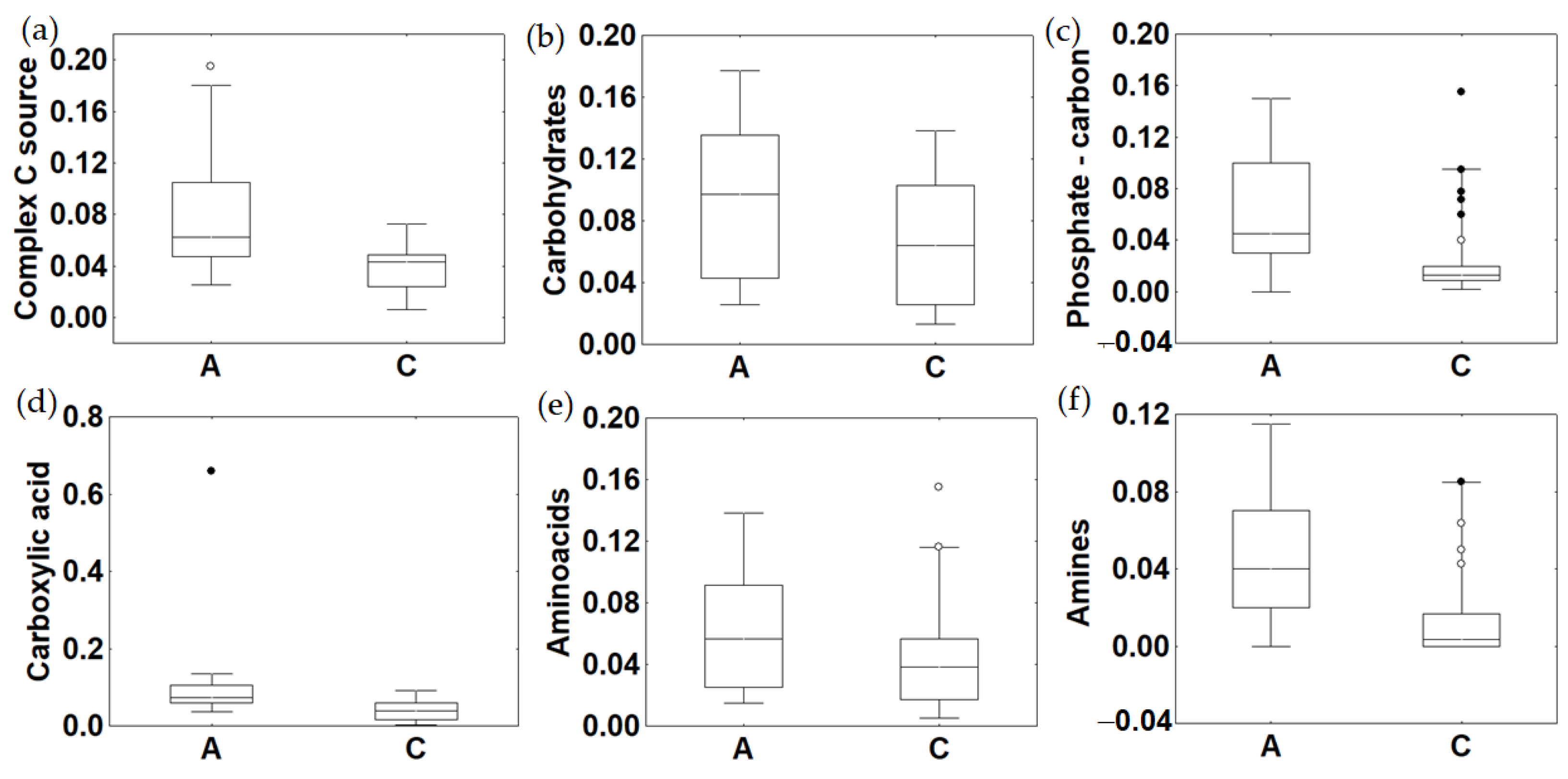
3.3. Trophic Variables
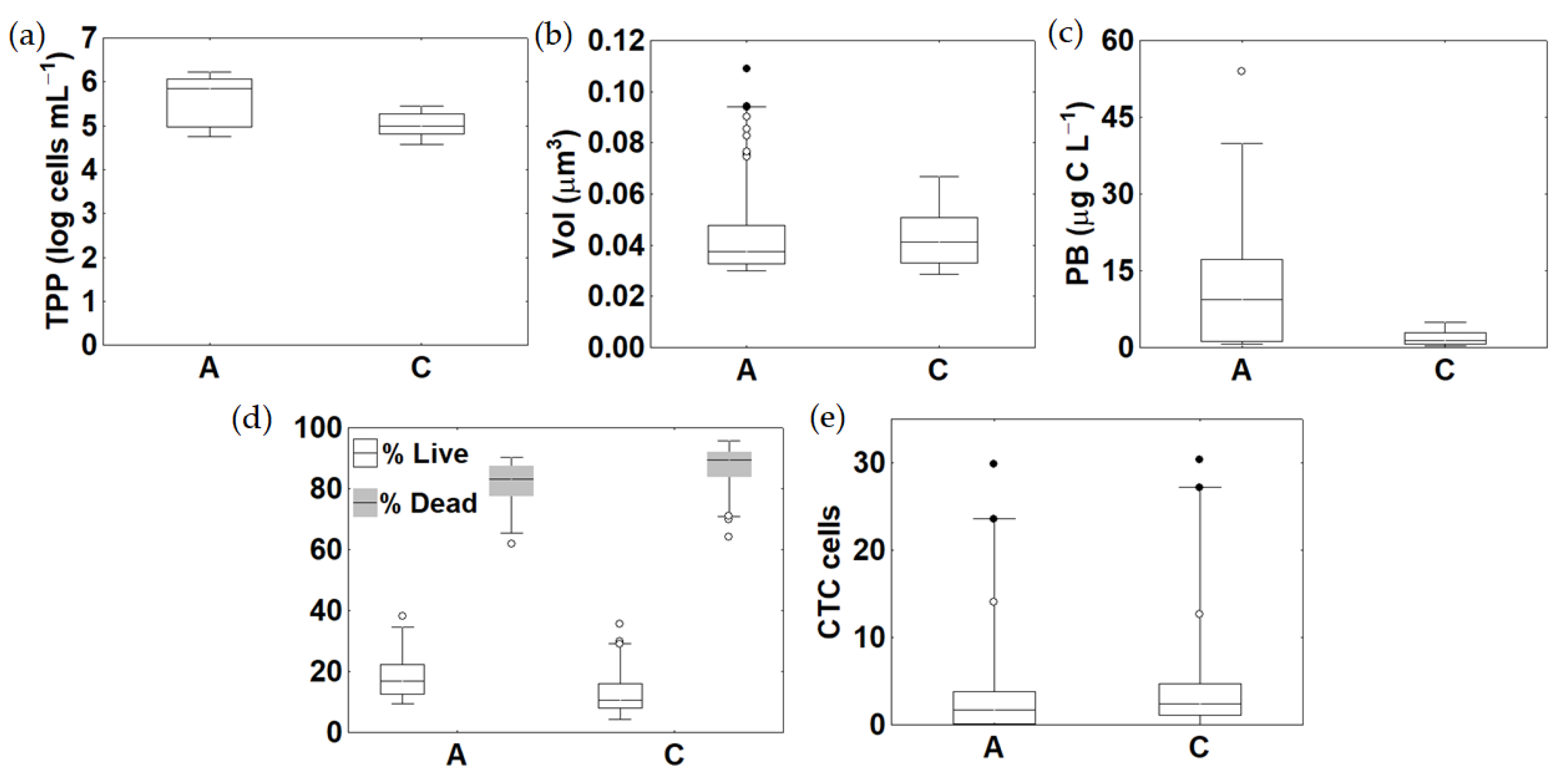
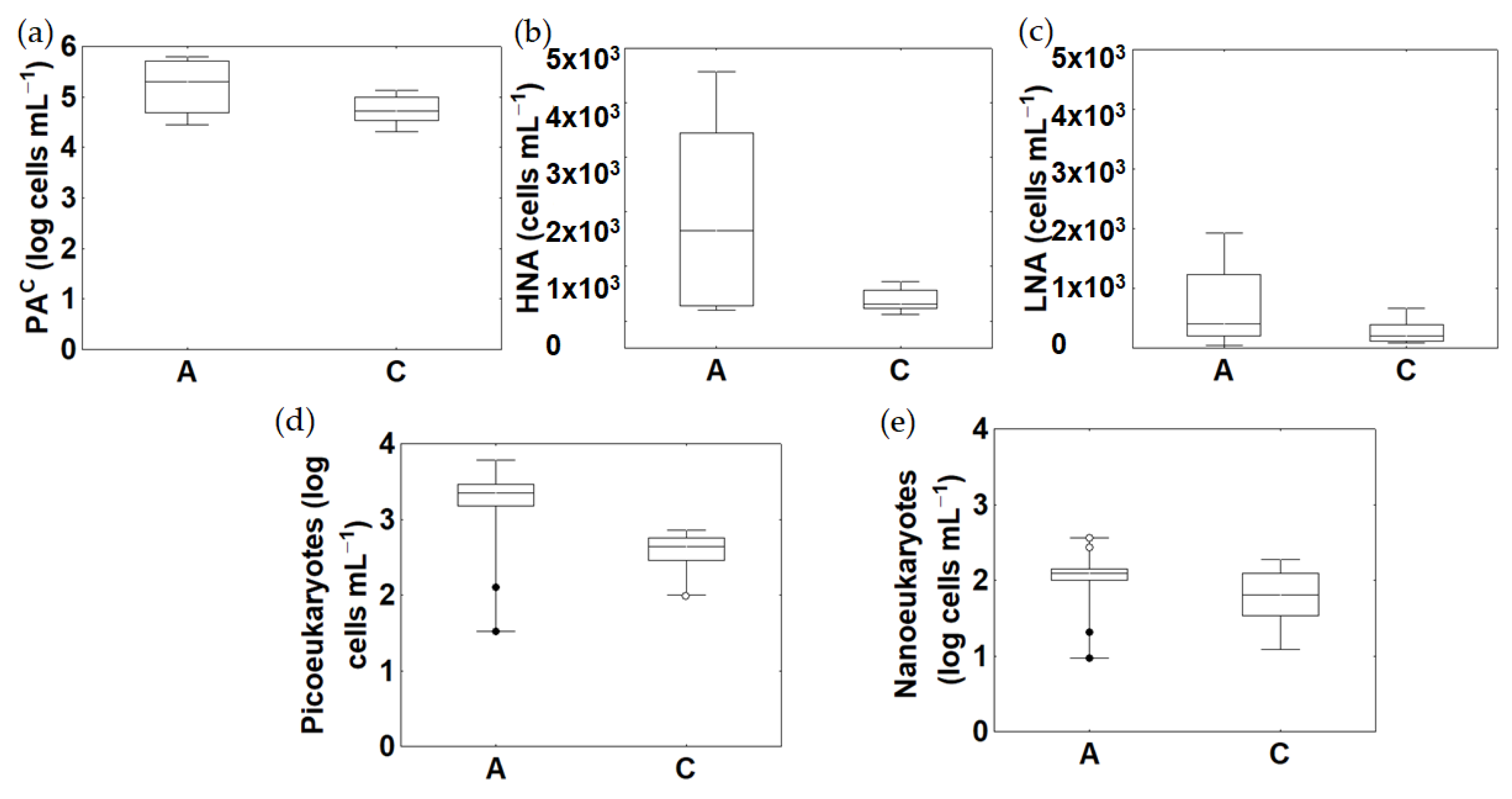
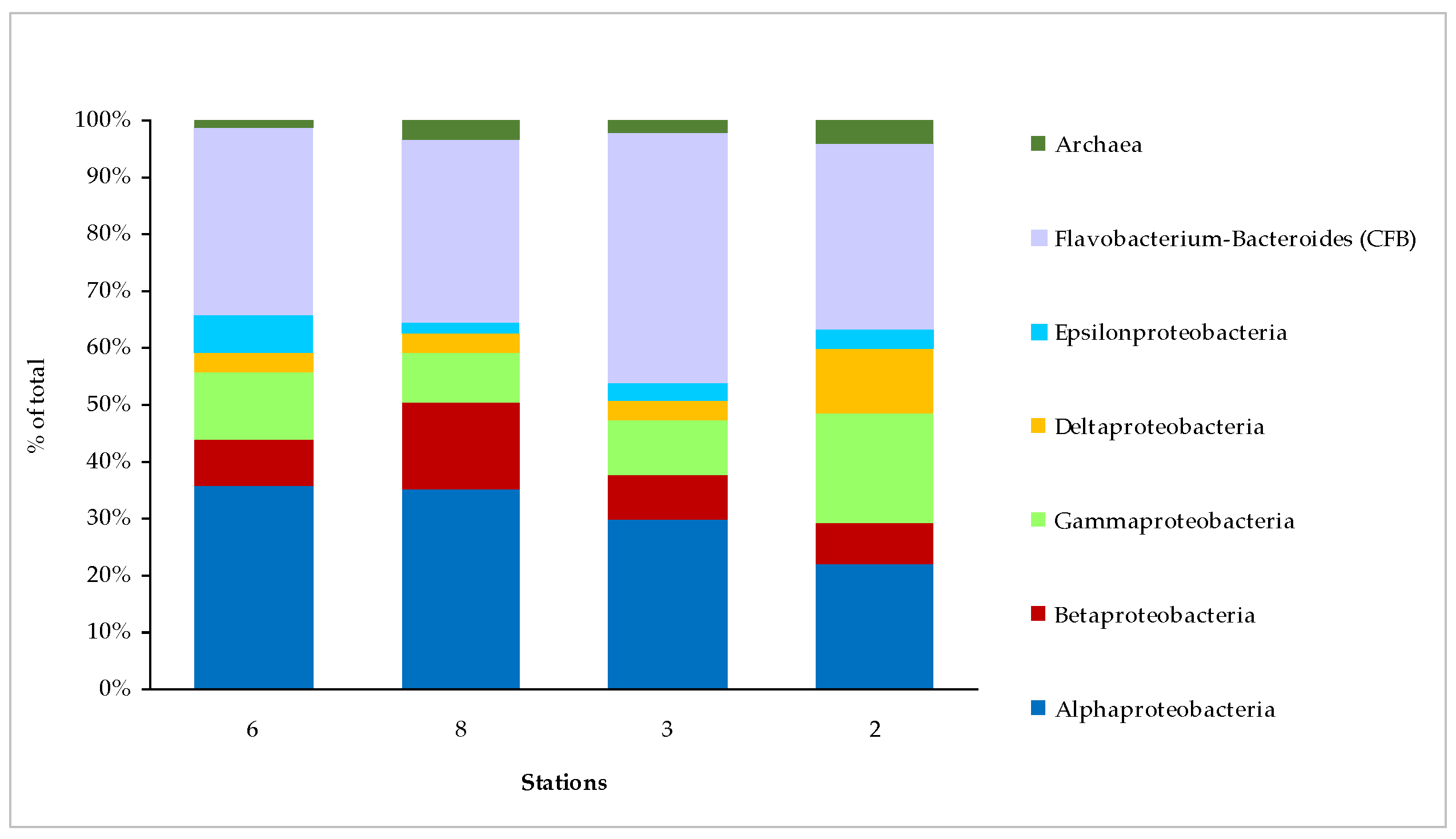
3.4. Microbial Abundance
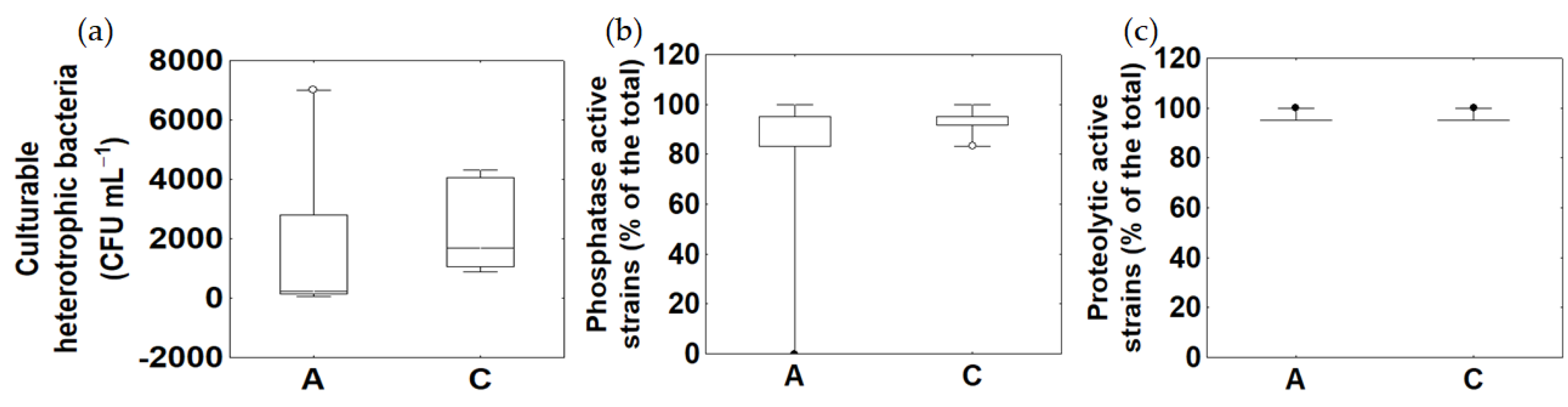
3.5. Microbial Metabolism
4. Discussion
4.1. Spatial Variability of Environmental and Microbial Parameters
4.2. Vertical Profiles of Abiotic and Biotic Variables
4.3. Major Drivers Affecting Environmental and Microbial Abundance, Structure and Metabolic Patterns
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Henley, S.F.; Cavan, E.L.; Fawcett, S.E.; Kerr, R.; Monteiro, T.; Sherrell, R.M.; Bowie, A.R.; Boyd, P.W.; Barnes, D.K.A.; Schloss, I.R.; et al. Changing Biogeochemistry of the Southern Ocean and Its Ecosystem Implications. Front. Mar. Sci. 2000, 7, 581. [Google Scholar] [CrossRef]
- Catalano, G.; Budillon, G.; La Ferla, R.; Povero, P.; Ravaioli, M.; Saggiomo, V.; Accornero, A.; Azzaro, M.; Carrada, G.C.; Giglio, F.; et al. A global budget of carbon and nitrogen in the Ross Sea (Southern Ocean). In Carbon and Nutrient Fluxes in Continental Margins: A Global Synthesis; Liu, K.K., Atkinson, L., Quinones, R., Talaue-McManus, L., Eds.; Global Change, The IGBP Series; Springer: Berlin, Germany, 2006; pp. 303–318. [Google Scholar]
- Smith, W.O., Jr.; Ainley, D.G.; Arrigo, K.R.; Dinniman, M.S. The oceanography and ecology of the Ross Sea. Ann. Rev. Mar. Sci. 2014, 6, 469–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrigo, K.R.; van Dijken, G.L. Annual changes in sea-ice, chlorophyll a, and primary production in the Ross Sea, Antarctica. Deep Sea Res. Part II 2004, 51, 117–138. [Google Scholar] [CrossRef]
- Smith, W.O., Jr. Primary productivity measurements in the Ross Sea, Antarctica: A regional synthesis. Earth Syst. Sci. Data 2022, 14, 2737–2747. [Google Scholar] [CrossRef]
- Silvano, A.; Foppert, A.; Rintoul, S.R.; Holland, P.R.; Tamura, T.; Kimura, N.; Castagno, P.; Falco, P.; Budillon, G.; Haumann, F.A. Recent recovery of Antarctic Bottom Water formation in the Ross Sea driven by climate anomalies. Nat. Geosci. 2020, 13, 780–786. [Google Scholar] [CrossRef]
- Castagno, P.; Capozzi, V.; DiTullio, G.R.; Falco, P.; Fusco, G.; Rintoul, S.R.; Spezie, G.; Budillon, G. Rebound of shelf water salinity in the Ross Sea. Nat. Commun. 2019, 10, 5441. [Google Scholar] [CrossRef] [Green Version]
- Brooks, C.M.; Bloom, E.; Kavanagh, A.; Nocito, E.S.; Watters, G.M.; Weller, J. The Ross Sea, Antarctica: A highly protected MPA in international waters. Mar. Policy 2021, 134, 104795. [Google Scholar] [CrossRef]
- Legendre, L.; Rivkin, R.B.; Weinbauer, M.G.; Guidi, L.; Uitz, J. The microbial carbon pump concept: Potential biogeochemical significance in the globally changing ocean. Progr. Oceanogr. 2015, 134, 432–450. [Google Scholar] [CrossRef] [Green Version]
- Misic, C.; Harriague, A.C.; Mangoni, O.; Cotroneo, Y.; Aulicino, G.; Castagno, P. Different responses of the trophic features of particulate organic matter to summer constraints in the Ross Sea. J. Mar. Syst. 2017, 166, 132–143. [Google Scholar] [CrossRef]
- Misic, C.; Povero, P.; Fabiano, M. Ectoenzymatic ratios in relation to particulate organic matter distribution (Ross Sea, Antarctica). Microb. Ecol. 2002, 44, 224–234. [Google Scholar] [CrossRef]
- Misic, C.; Povero, P.; Fabiano, M. Relationship between ecto-enzymatic activity and organic substrates availability (Ross Sea, Antarctica): An experimental approach. Polar Biol. 1998, 20, 367–376. [Google Scholar] [CrossRef]
- Ducklow, H.; Carlson, C.; Church, M.; Kirchman, D.; Smith, D.; Steward, G. The seasonal development of the bacterioplankton bloom in the Ross Sea, Antarctica, 1994–1997. Deep Sea Res. Part II 2001, 48, 4199–4221. [Google Scholar] [CrossRef]
- Monticelli, L.S.; La Ferla, R.; Maimone, G. Dynamics of bacterioplankton activities after a summer phytoplankton bloom period in Terra Nova Bay. Antarct. Sci. 2003, 15, 85–93. [Google Scholar] [CrossRef]
- Celussi, M.; Bergamasco, A.; Cataletto, B.; Umani, S.F.; Del Negro, P. Water masses’ bacterial community structure and microbial activities in the Ross Sea, Antarctica. Antarct. Sci. 2010, 22, 361–370. [Google Scholar] [CrossRef]
- Celussi, M.; Cataletto, B.; Fonda Umani, S.; Del Negro, P. Depth profiles of bacterioplankton assemblages and their activities in the Ross Sea. Deep Sea Res. Part I 2009, 56, 2193–2205. [Google Scholar] [CrossRef]
- Mangoni, O.; Saggiomo, V.; Bolinesi, F.; Margiotta, F.; Budillon, G.; Cotroneo, Y.; Misic, C.; Rivaro, P.; Saggiomo, M. Phytoplankton blooms during austral summer in the Ross Sea, Antarctica: Driving factors and trophic implications. PLoS ONE 2017, 12, e0176033. [Google Scholar] [CrossRef] [Green Version]
- Bolinesi, F.; Saggiomo, M.; Ardini, F.; Castagno, P.; Cordone, A.; Fusco, G.; Rivaro, P.; Saggiomo, V.; Mangoni, O. Spatial-Related Community Structure and Dynamics in Phytoplankton of the Ross Sea, Antarctica. Front. Mar. Sci. 2020, 7, 574963. [Google Scholar] [CrossRef]
- Zaccone, R.; Misic, C.; Azzaro, F.; Azzaro, M.; Maimone, G.; Mangoni, O.; Fusco, G.; Rappazzo, A.C.; La Ferla, R. Regulation of Microbial Activity Rates by Organic Matter in the Ross Sea during the Austral Summer 2017. Microorganisms 2020, 8, 1273. [Google Scholar] [CrossRef]
- Hoppe, H.G.; Arnosti, C.; Herndl, G. Ecological significance of bacterial enzymes in the marine environment. In Enzymes in the Environment: Activity, Ecology and Applications; Burns, R.G., Dick, R.P., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 85–125. [Google Scholar] [CrossRef]
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef]
- Azam, F.; Smith, D.C.; Steward, G.F.; Hagström, Å. Bacteria-organic matter coupling and its significance for oceanic carbon cycling. Microb. Ecol. 1993, 28, 167–179. [Google Scholar] [CrossRef]
- Cho, B.; Azam, F. Major role of bacteria in biogeochemical fluxes in the ocean's interior. Nature 1988, 332, 441–443. [Google Scholar] [CrossRef]
- D’Sa, E.J.; Kim, H.-C.; Ha, S.-Y.; Joshi, I. Ross Sea Dissolved Organic Matter Optical Properties During an Austral Summer: Biophysical Influences. Front. Mar. Sci. 2021, 8, 749096. [Google Scholar] [CrossRef]
- Azzaro, M.; Packard, T.T.; Monticelli, L.S.; Maimone, G.; Rappazzo, A.C.; Azzaro, F.; Grilli, F.; Crisafi, E.; La Ferla, R. Microbial metabolic rates in the Ross Sea: The ABIOCLEAR Project. Nat. Conserv. 2019, 34, 441–475. [Google Scholar] [CrossRef] [Green Version]
- Kohut, J.T.; Kustka, A.B.; Hiscock, M.R.; Lam, P.J.; Measures, C.; Milligan, A.; White, A.; Carvalho, F.; Hatta, M.; Jones, B.M.; et al. Mesoscale variability of the summer bloom over the northern Ross Sea shelf: A tale of two banks. J. Mar. Syst. 2017, 166, 50–60. [Google Scholar] [CrossRef]
- Monien, D.; Monien, P.; Brünjes, R.; Widmer, T.; Kappenberg, A.; Silva Busso, A.A.; Schnetger, B.; Brumsack, H. Meltwater as a source of potentially bioavailable iron to Antarctica waters. Antarctic Sci. 2017, 29, 277–291. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.; Russell, G.; Rind, D.; Stone, P.; Lacis, A.; Lebedeff, S.; Ruedy, R.; Travis, L. Efficient three-dimensional global models for climate studies: Models I and II. Mon. Weather Rev. 1983, 111, 609–662. [Google Scholar] [CrossRef]
- Caruso, G.; Azzaro, M.; Caroppo, C.; Decembrini, F.; Monticelli, L.S.; Leonardi, M.; Maimone, M.; Zaccone, R.; La Ferla, R. Microbial community and its potential as descriptor of environmental status. ICES J. Mar. Sci. 2016, 73, 2174–2177. [Google Scholar] [CrossRef] [Green Version]
- Iseki, K.; MacDonald, R.W.; Carmack, E. Distribution of particulate matter in the Southeastern Beaufort Sea in late summer. Proc. NIPR Symp. Polar Biol. 1987, 1, 35–46. [Google Scholar]
- Specchiulli, A.; Cilenti, L.; D'Adamo, R.; Fabbrocini, A.; Guo, W.; Huang, L.; Lugliè, A.; Padedda, B.M.; Scirocco, T.; Magni, P. Dissolved organic matter dymics in Mediterranean lagoons: The relationship between DOC and CDOM. Mar. Chem. 2018, 202, 37–48. [Google Scholar] [CrossRef]
- Specchiulli, A.; Bignami, F.; Marini, M.; Fabbrocini, A.; Scirocco, T.; Campanelli, A.; Penna, P.; Santucci, A.; D'Adamo, R. The role of forcing agents on biogeochemical variability along the southwestern Adriatic coast: The Gulf of Manfredonia case study. Estuar. Coast. Shelf Sci. 2016, 183, 136–149. [Google Scholar] [CrossRef]
- Porter, K.G.; Feig, Y.S. The use of DAPI for identifying and counting aquatic microflora. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- La Ferla, R.; Maimone, G.; Azzaro, M.; Conversano, F.; Brunet, C.; Cabral, A.S.; Paranhos, R. Vertical distribution of the prokaryotic cell size in the Mediterranean Sea. Helgol. Mar. Res. 2012, 66, 635–650. [Google Scholar] [CrossRef] [Green Version]
- La Ferla, R.; Azzaro, M.; Michaud, L.; Caruso, G.; Lo Giudice, A.; Paranhos, R.; Cabral, A.S.; Conte, A.; Cosenza, A.; Maimone, G.; et al. Prokaryotic abundance and activity in permafrost of the Northern Victoria Land and upper Victoria Valley (Antarctica). Microb. Ecol. 2017, 74, 402–415. [Google Scholar] [CrossRef]
- Teira, E.; Reinthaler, T.; Pernthaler, A. Combining catalysed reporter deposition-fluorescence in situ hybridization and microautoradiography to detect substrate utilization by bacteria and Archaea in the deep ocean. Appl. Environ. Microbiol. 2004, 70, 4411–4414. [Google Scholar] [CrossRef] [Green Version]
- Pernthaler, A.; Pernthaler, J.; Amann, R. Sensitive multi-color fluorescence in situ hybridization for the identification of environmental microorganisms. In Molecular Microbial Ecology Manual, 2nd ed.; Kowalchuk, G., Bruijn, F.J., Head, I.M., Akkermans, A.D., Elsas, J.D., Eds.; Kluwer Academic Press: Dordrecht, The Netherlands, 2004; Volume 11, pp. 711–726. [Google Scholar]
- Wallner, G.; Amann, R.; Beisker, W. Optimizing fluorescent in situ hybridization of suspended cells with rRNA-targeted oligonucleotide probes for the flow cytometric identification of microorganisms. Cytometry 1993, 14, 136–143. [Google Scholar] [CrossRef]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [CrossRef] [Green Version]
- Daims, H.; Brühl, A.; Amann, R.; Schleifer, K.H.; Wagner, M. The domain-specific probe EUB338 is insufficient for the detection of all Bacteria: Development and evaluation of a more comprehensive probe set. Syst. Appl. Microbiol. 1999, 22, 434–444. [Google Scholar] [CrossRef]
- Neef, A. Application of In Situ single-cell Identification of Bacteria for Population Analysis in Complex Microbial Communities. Ph.D. Thesis, Technical University Munich, Munich, Germany, 1997. [Google Scholar]
- Manz, W.; Amann, R.; Ludwig, W.; Wagner, M.; Schleifer, K.-H. Phylogenetic oligodeoxynucleotide probes for the major subclasses of proteobacteria: Problems and solutions. Syst. Appl. Microbiol. 1992, 15, 593–600. [Google Scholar] [CrossRef]
- Loy, A.; Lehner, A.; Lee, N.; Adamczyk, J.; Meier, H.; Ernst, J.; Schleifer, K.H.; Wagner, M. Oligonucleotide microarray for 16S rRNA gene-based detection of all recognized lineages of sulfate reducing prokaryotes in the environment. Appl. Environ. Microbiol. 2002, 68, 5064–5081. [Google Scholar] [CrossRef] [Green Version]
- Loy, A.; Maixner, F.; Wagner, M.; Horn, M. ProbeBase: An online resource for rRNA-targeted oligonucleotide probes. Nucleic Acids Res. 2007, 35, 800–804. [Google Scholar] [CrossRef]
- Manz, W.; Amann, R.; Ludwig, W.; Vancanneyt, M.; Schleifer, K.H. Application of a suite of 16S rRNA specific oligonucleotide probes designed to investigate bacteria of the phylum Cytophaga-Flavobacter-Bacteroides in the natural environment. Microbiology 1996, 142, 1097–1106. [Google Scholar] [CrossRef] [Green Version]
- Stahl, D.A.; Amann, R. Development and application of nucleic acid probes. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrand, E., Goodfellow, M., Eds.; Wiley: Chichester, UK, 1991; pp. 205–248. [Google Scholar]
- Gasol, J.M.; del Giorgio, P.A. Using flow cytometry for counting natural planktonic bacteria and understanding the structure of planktonic bacterial communities. Sci. Mar. 2000, 64, 197–224. [Google Scholar] [CrossRef] [Green Version]
- Brussaard, C.P.D. Viral control of phytoplankton population—A review. J. Eukaryot. Microbiol. 2004, 51, 125–138. [Google Scholar] [CrossRef]
- Marie, D.; Simon, N.; Vaulot, D. Phytoplankton cell counting by flow cytometry. In Algal Culturing Techniques; Andersen, R.A., Ed.; Physiological Society of America: Oxford, UK, 2005; pp. 253–267. [Google Scholar]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [Green Version]
- Garland, J.L. Analytical approaches to the characterization of samples of microbial communities using patterns of potential C source utilization. Soil Biol. Biochem. 1996, 28, 213–221. [Google Scholar] [CrossRef]
- Sala, M.M.; Arin, L.; Balagué, V.; Felipe, J.; Guadayol, O.; Vaqué, D. Functional diversity of bacterioplankton assemblages in Western Antarctic seawaters during late spring. Mar. Ecol. Progr. Ser. 2005, 292, 13–21. [Google Scholar] [CrossRef]
- Caruso, G.; Dell'Acqua, O.; Caruso, R.; Azzaro, M. Phenotypic characterization of bacterial isolates from marine waters and plastisphere communities of the Ross Sea (Antarctica). J. Clin. Microbiol. Biochem. Technol. 2022, 8, 001–009. [Google Scholar] [CrossRef]
- Clarke, K.R.; Green, R.H. Statistical design and analysis for a “biological effects” study. Mar. Ecol. Prog. Ser. 1988, 46, 213–226. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An. Approach to Statistical Analysis and Interpretation; Primer-E Ltd.: Plymouth, UK, 2001. [Google Scholar]
- Cavicchioli, R. Microbial ecology of Antarctic aquatic systems. Nat. Rev. Microbiol. 2015, 13, 691–706. [Google Scholar] [CrossRef]
- Rivaro, P.; Abelmoschi, M.L.; Grotti, M.; Ianni, C.; Magi, E.; Margiotta, F.; Massolo, S.; Saggiomo, V. Combined effects of hydrographic structure and iron and copper availability on the phytoplankton growth in Terra Nova Bay Polynya (Ross Sea, Antarctica). Deep Sea Res. I 2012, 62, 97–110. [Google Scholar] [CrossRef]
- Carlson, C.A.; Hansell, D.A.; Peltzer, E.T.; Smith, W.O., Jr. Stocks and dynamics of dissolved and particulate organic matter in the southern Ross Sea, Antarctica. Deep Sea Res. Part II Top. Stud. Oceanogr. 2000, 47, 3201–3225. [Google Scholar] [CrossRef]
- Schultze, S.; Weishaar Andersen, T.; Hessen, D.O.; Ruus, A.; Borgå, K.; Poste, A.E. Land-cover, climate and fjord morphology drive differences in organic matter and nutrient dynamics in two contrasting northern river-fjord systems. Estuar. Coast. Shelf Sci. 2022, 270, 107831. [Google Scholar] [CrossRef]
- Weishaar, J.L.; Aiken, G.R.; Bergamaschi, B.A.; Fram, M.S.; Fujii, R.; Mopper, K. Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ. Sci. Technol. 2003, 37, 4702–4708. [Google Scholar] [CrossRef] [PubMed]
- Peeters, K.; Hodgson, D.A.; Convey, P.; Willems, A. Culturable diversity of heterotrophic bacteria in Forlidas Pond (Pensacola Mountains) and Lundström Lake (Shackleton Range), Antarctica. Microb. Ecol. 2011, 62, 399–413. [Google Scholar] [CrossRef]
- Michaud, L.; Caruso, C.; Mangano, S.; Interdonato, F.; Bruni, V.; Lo Giudice, A. Predominance of Flavobacterium, Pseudomonas, and Polaromonas within the prokaryotic community of freshwater shallow lakes in the northern Victoria Land, East Antarctica. FEMS Microbiol. Ecol. 2012, 82, 391–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Ferla, R.; Maimone, G.; Lo Giudice, A.; Azzaro, F.; Cosenza, A.; Azzaro, M. Cell size and other phenotypic traits of prokaryotic cells in pelagic areas of the Ross Sea (Antarctica). Hydrobiologia 2015, 761, 181–194. [Google Scholar] [CrossRef]
- Gilichinsky, D.; Vishnivetskaya, T.; Petrova, M. Bacteria in permafrost. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Schinner, F., Marx, J.C., Gerday, C., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2008; pp. 83–102. [Google Scholar]
- Zaccone, R.; Caruso, G. Microbial hydrolysis of polysaccharides and organic phosphates in the Northern Adriatic Sea. Chem. Ecol. 2002, 18, 85–94. [Google Scholar] [CrossRef]
- Bauer, J.E.; Bianchi, T.S. Dissolved Organic Carbon Cycling and Transformation. In Treatise on Estuarine and Coastal Science; Wolanski, E., Mc Lusky, D.S., Eds.; Academic Press: Waltham, MA, USA, 2011; Volume 5, pp. 7–67. [Google Scholar]
- Piontek, J.; Sperling, M.; Nothing, E.M.; Engel, A. Regulation of bacterioplankton activity in Fram Strait (Arctic Ocean) during early summer: The role of organic matter supply and temperature. J. Mar. Syst. 2014, 132, 83–94. [Google Scholar] [CrossRef]
- Bowman, J.P.; McCammon, S.A.; Dann, A.L. Biogeographic and quantitative analyses of abundant uncultivated gamma-proteobacterial clades from marine sediment. Microb. Ecol. 2005, 49, 451–460. [Google Scholar] [CrossRef]
- Ravenschlag, K.; Sahm, K.; Amann, R. Quantitative molecular analysis of the microbial community in marine arctic sediments (Svalbard). Appl. Environ. Microbiol. 2001, 67, 387–395. [Google Scholar] [CrossRef]
- Pearce, D.A.; Hodgson, D.A.; Thorne, M.A.S.; Burns, G.; Cockell, C.S. Preliminary analysis of life within a former subglacial lake sediment in Antarctica. Diversity 2013, 5, 680–702. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A area | |||
| Stations | Lat (S) | Long (E) | Max. depth (m) |
| 2 | 75°35.1282′ | 165°28.4682′ | 842 |
| 3 | 75°31.6676′ | 165°43.1940′ | 789 |
| 6 | 75°12.3186′ | 163°33.0294′ | 1108 |
| 8 | 75°20.9871′ | 164°41.00′ | 683 |
| 11 | 75°07.0896′ | 164°09.8970′ | 971 |
| 12 | 75°04.3128′ | 163°42.2874′ | 867 |
| 14 | 74°55′39.598″ | 163°59′46.799″ | 344 |
| 15 | 74°42.414′ | 164°13.5088′ | 498 |
| 18 | 75°10.426′ | 165°2.3732′ | 1054 |
| 19 | 75°00.2847′ | 165°07.5264′ | 925 |
| 20 | 74°47.456′ | 165°23.582′ | 662 |
| 21 | 74°52.4730′ | 166°34.7040′ | 886 |
| 22 | 75°04.5138′ | 166°24.2760′ | 855 |
| 23 | 75°14.1960′ | 166°10.9332′ | 852 |
| C area | |||
| Stations | Lat (S) | Long (E) | Max. depth (m) |
| 63 | 72°12.7692′ | 172°00.1914′ | 415 |
| 64 | 72°16.9812′ | 172°30.0396′ | 516 |
| 65 | 72°22.2870′ | 172°59.6796′ | 513 |
| 66 | 72°27.0273′ | 173°29.9220′ | 480 |
| 67 | 72°31.2120′ | 174°00.1422′ | 430 |
| 68 | 72°35.5641′ | 174°30.0630′ | 406 |
| 74 | 71°48.3646′ | 173°35.2806′ | 1969 |
| 75 | 71°53.5158′ | 174°04.4184′ | 1870 |
| 76 | 71°58.0998′ | 174°34.5156′ | 1786 |
| 77 | 72°02.4300′ | 175°04.7070′ | 1623 |
| Probes | Target Group | Probe Sequence (5′-3′) | References |
|---|---|---|---|
| EUB338I | Most, but not all bacteria | GCTGCCTCCCGTAGGAGT | [39] |
| EUB338II | Planctomycetes | GCAGCCACCCGTAGGTGT | [40] |
| EUB338III | Verrucomicrobiales | GCTGCCACCCGTAGGTGT | [40] |
| ALF968 | Alphaproteobacteria | GGTAAGGTTCTGCGCGTT | [41] |
| BET42a | Betaproteobacteria | GCCTTCCCACTTCGTTT | [42] |
| GAM42a | Gammaproteobacteria | GCCTTCCCACATCGTTT | [42] |
| DELTA495a | Most Deltaproteobacteria | AGTTAGCCGGTGCTTCCT | [43] |
| DELTA495b | Deltaproteobacteria | AGTTAGCCGGCGCTTCCT | [43] |
| DELTA495c | Deltaproteobacteria | AATTAGCCGGTGCTTCCT | [43] |
| EPSY914 | Epsilonproteobacteria | GGTCCCCGTCTATTCCTT | [44] |
| CF319a | Bacteroidetes | TGGTCCGTGTCTCAGTAC | [45] |
| ARCH915 | Archaea | GTGCTCCCCCGCCAATTCCT | [46] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azzaro, M.; Specchiulli, A.; Maimone, G.; Azzaro, F.; Lo Giudice, A.; Papale, M.; La Ferla, R.; Paranhos, R.; Souza Cabral, A.; Rappazzo, A.C.; et al. Trophic and Microbial Patterns in the Ross Sea Area (Antarctica): Spatial Variability during the Summer Season. J. Mar. Sci. Eng. 2022, 10, 1666. https://doi.org/10.3390/jmse10111666
Azzaro M, Specchiulli A, Maimone G, Azzaro F, Lo Giudice A, Papale M, La Ferla R, Paranhos R, Souza Cabral A, Rappazzo AC, et al. Trophic and Microbial Patterns in the Ross Sea Area (Antarctica): Spatial Variability during the Summer Season. Journal of Marine Science and Engineering. 2022; 10(11):1666. https://doi.org/10.3390/jmse10111666
Chicago/Turabian StyleAzzaro, Maurizio, Antonietta Specchiulli, Giovanna Maimone, Filippo Azzaro, Angelina Lo Giudice, Maria Papale, Rosabruna La Ferla, Rodolfo Paranhos, Anderson Souza Cabral, Alessandro Ciro Rappazzo, and et al. 2022. "Trophic and Microbial Patterns in the Ross Sea Area (Antarctica): Spatial Variability during the Summer Season" Journal of Marine Science and Engineering 10, no. 11: 1666. https://doi.org/10.3390/jmse10111666
APA StyleAzzaro, M., Specchiulli, A., Maimone, G., Azzaro, F., Lo Giudice, A., Papale, M., La Ferla, R., Paranhos, R., Souza Cabral, A., Rappazzo, A. C., Renzi, M., Castagno, P., Falco, P., Rivaro, P., & Caruso, G. (2022). Trophic and Microbial Patterns in the Ross Sea Area (Antarctica): Spatial Variability during the Summer Season. Journal of Marine Science and Engineering, 10(11), 1666. https://doi.org/10.3390/jmse10111666