
Distribution and Identification of Ulva aragoensis (Ulvaceae, Chlorophyta), a Constituent Species of Green Tides in the Southern Yellow Sea, Based on Molecular Data


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
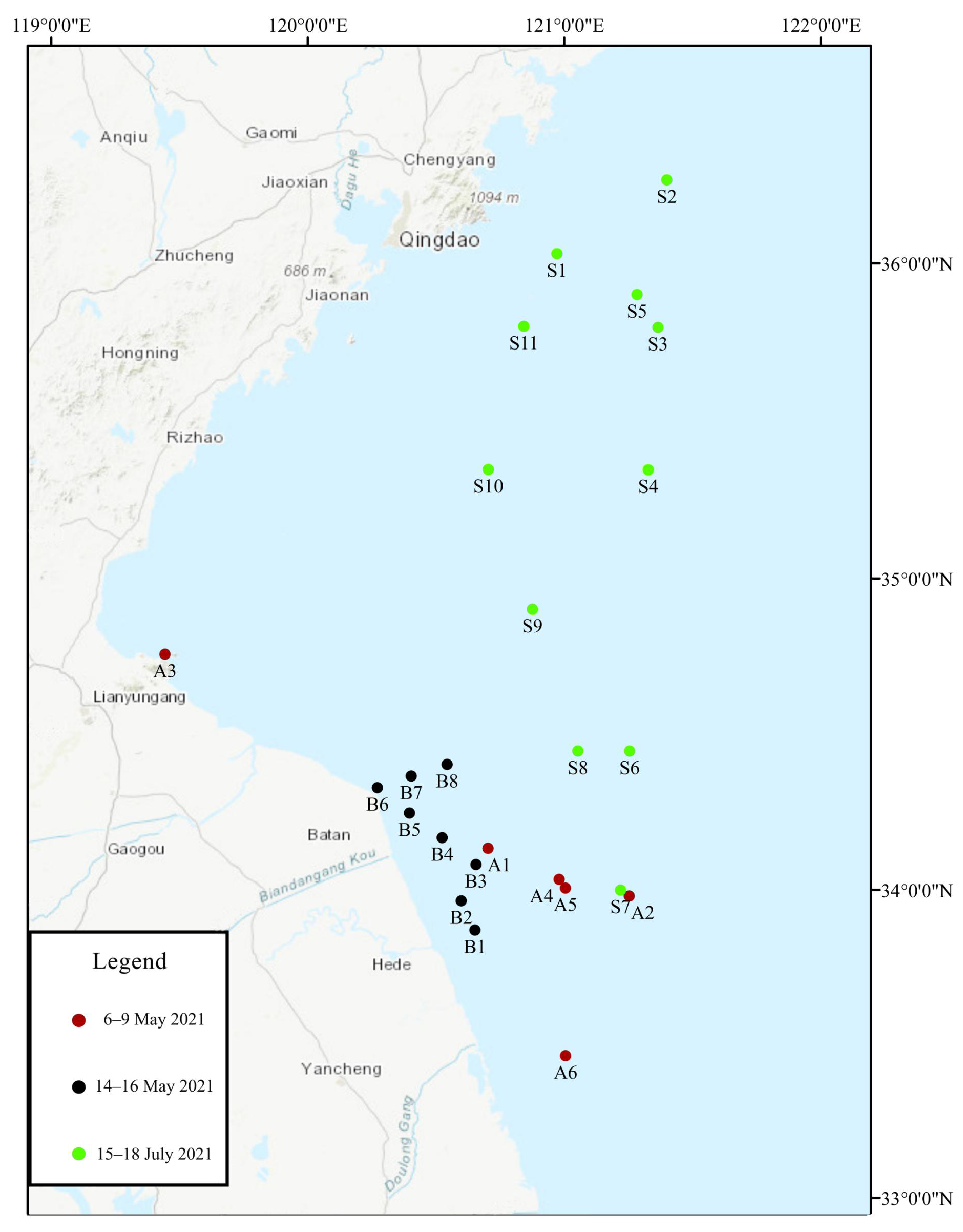
2.1. Sampling Point Setting and Sampling Process
2.2. Micro-Propagule Cultivation and Pretreatment of Green Macroalgae Samples
2.3. Identification of U. aragoensis
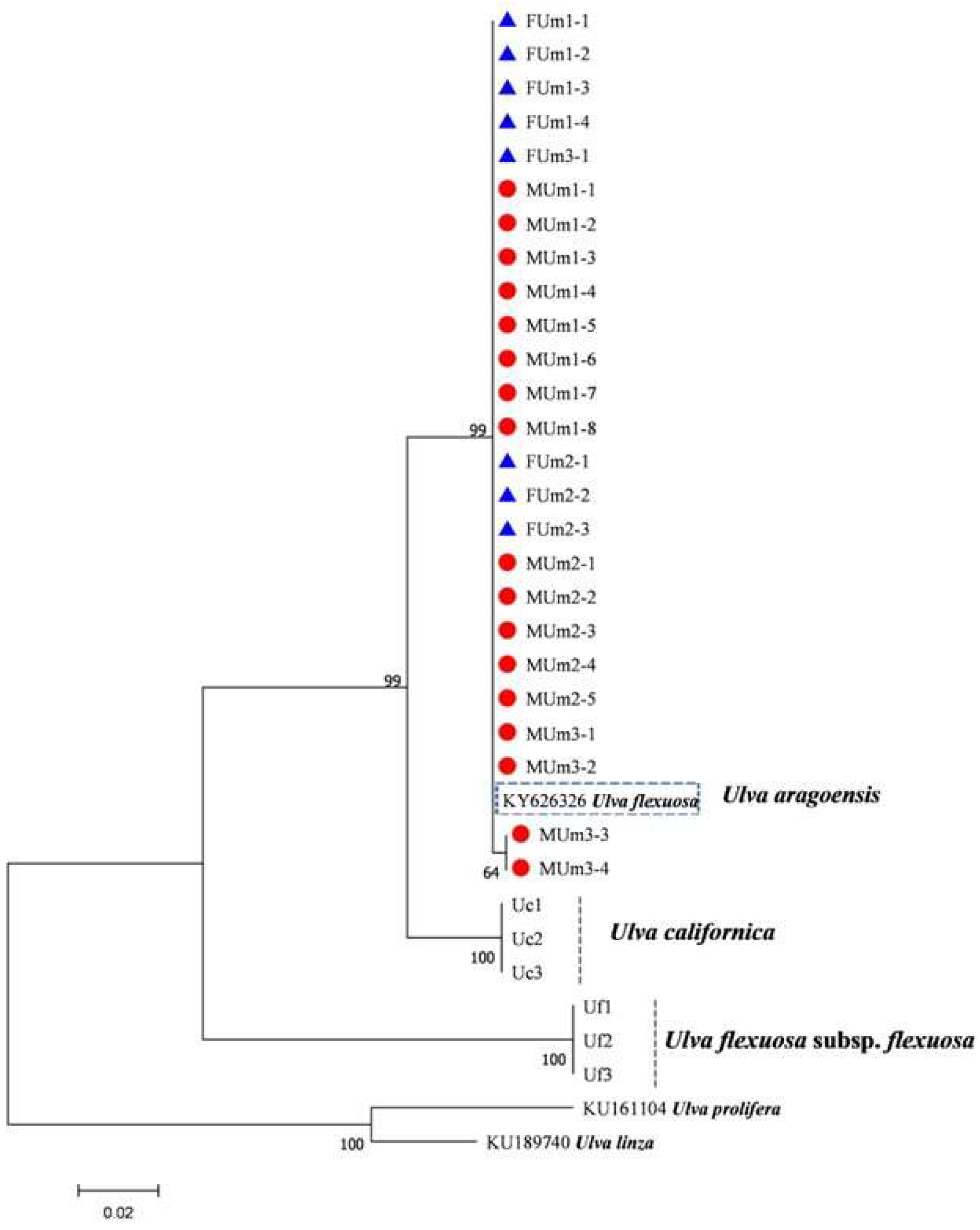
2.4. Sequencing of Multiple DNA Barcodes and Phylogenetic Tree Construction of U. aragoensis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence | Direction | Reference |
|---|---|---|---|
| ITS-F | 5′-TCGTAACAAGGTTTCCGTAGG-3′ | Forward | [36] |
| ITS-R | 5′-TTCCTTCCGCTTATTGATATGC-3′ | Reverse | [36] |
| tufA-F | 5′-GGNGCNGCNCAAATGGAYGG-3′ | Forward | [37] |
| tufA-R | 5′-CCTTCNCGAATMGCRAAWCGC-3′ | Reverse | [37] |
| 18S-F | 5′-GGAGGATTAGGGTCCGATTCC-3′ | Forward | [38] |
| 18S-R | 5′-CTTCCGTCAATTCCTTTAAG-3′ | Reverse | [38] |
| LSU-F | 5′-AMAAGTACCRYGAGGGAAAG-3′ | Forward | [15] |
| LSU-R | 5′-GCACTAATCATTCGCTTTACC-3′ | Reverse | [15] |
| psbA-F | 5′-YTHTAYCCWATHTGGGAAGC-3′ | Forward | [39] |
| psbA-R | 5′-GGGAAGTTRTGNGCRTTRCG-3′ | Reverse | [39] |
| rbcL-F | 5′-ATGTCACCACAAACAGAAACTAAAGC-3′ | Forward | [40] |
| rbcL-R | 5′-AATTCAAATTTAATTTCTTTCC-3′ | Reverse | [40] |
| rps2-trnL-F | 5′-AAAATCAAAATCTAGTAAACCAGGC-3′ | Forward | [41] |
| rps2-trnL-R | 5′-GCTAGCGATTCTTAACGCGATTGGG-3′ | Reverse | [41] |
| Target | PCR Reaction Profile |
|---|---|
| ITS | 94 °C for 5 min, followed by 30 cycles of 94 °C for 40 s, 55 °C for 40 s, and 65 °C for 70 s, with the final step at 65 °C for 10 min |
| tufA | 94 °C for 2 min, followed by 35 cycles of 94 °C for 1 min, 50 °C for 30 s, and 72 °C for 1 min, with the final step at 72 °C for 10 min |
| 18S | 95 °C for 2 min, followed by 35 cycles of 95 °C for 1 min, 50 °C for 1 min, and 72 °C for 4 min, with the final step at 72 °C for 6 min |
| LSU | 94 °C for 5 min, followed by 38 cycles of 94 °C for 30 s, 50 °C for 30 s, and 72 °C for 1 min, with the final step at 72 °C for 7 min |
| psbA | 94 °C for 10 min, followed by 30 cycles of 94 °C for 1 min, 57 °C for 1 min, and 72 °C for 50 s, with the final step at 72 °C for 10 min |
| rbcL | 94 °C for 3 min, followed by 35 cycles of 94 °C for 1 min, 45 °C for 2 min, and 65 °C for 3 min, with the final step at 72 °C for 10 min |
| rps2-trnL | 94 °C for 5 min, followed by 30 cycles of 94 °C for 30 s, 52 °C for 40 s, and 72 °C for 40 s, with the final step at 72 °C for 5 min |
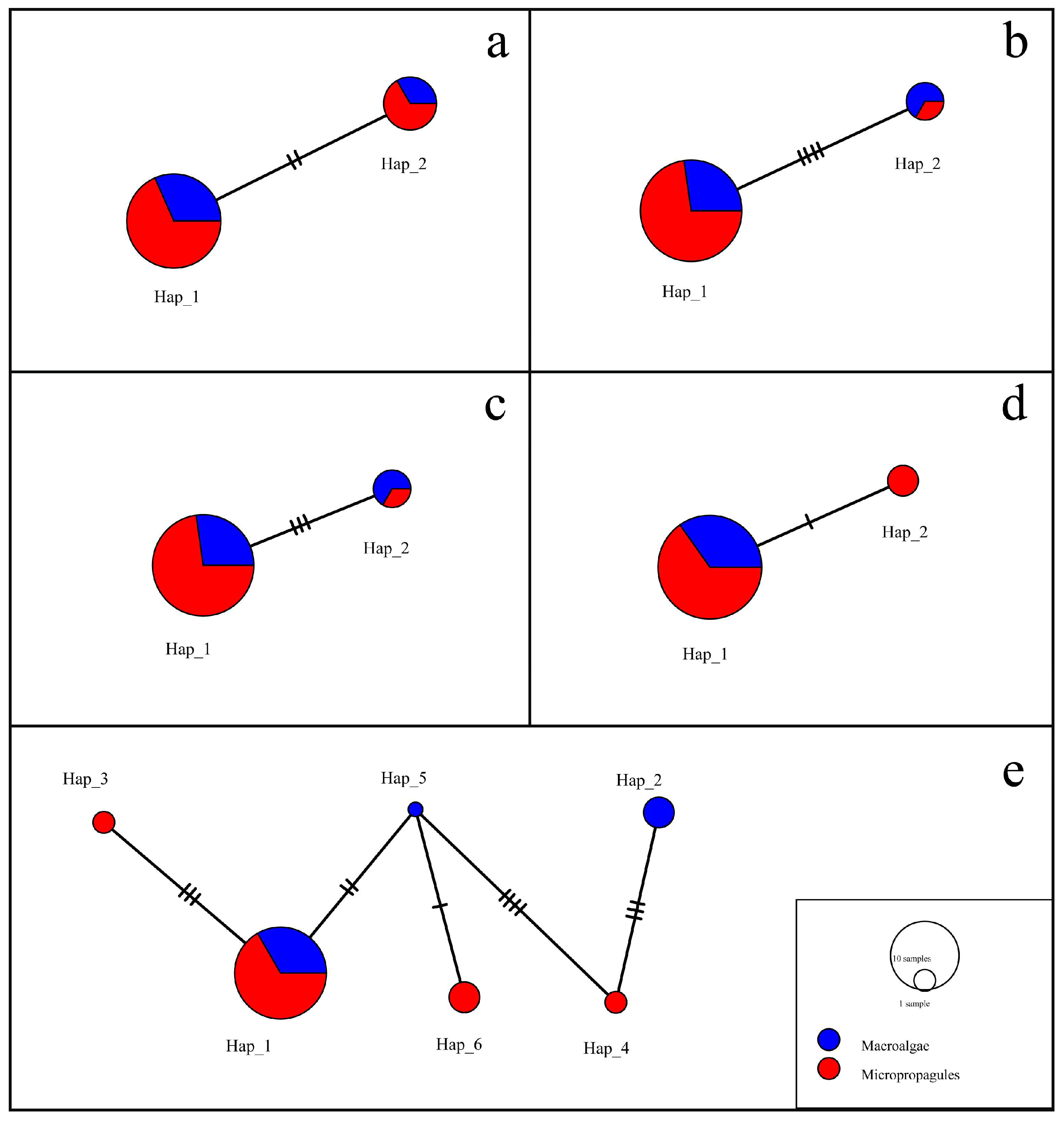
2.5. Haplotype Analysis of U. aragoensis
3. Results
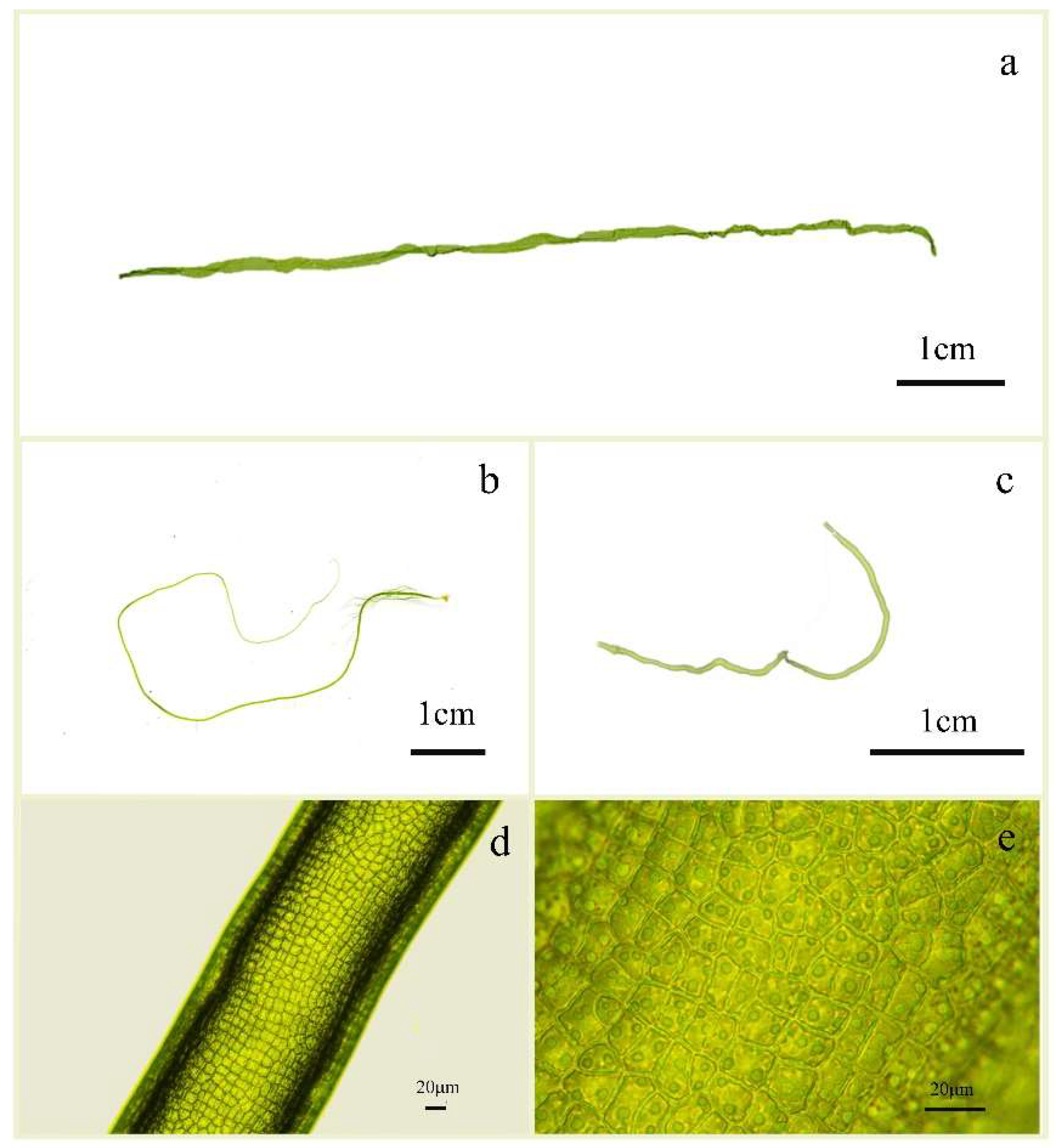
3.1. Morphological Characteristics of U. aragoensis
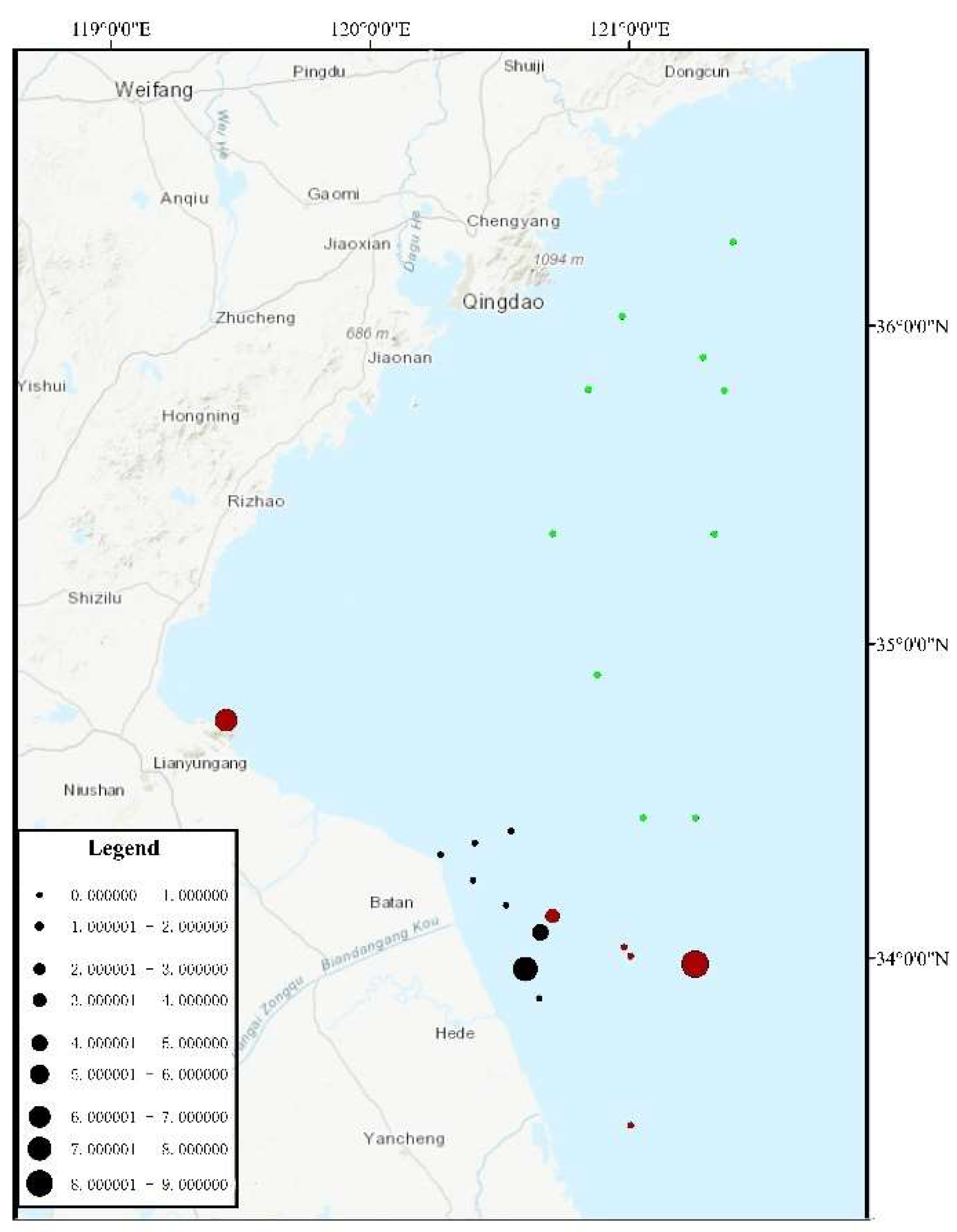
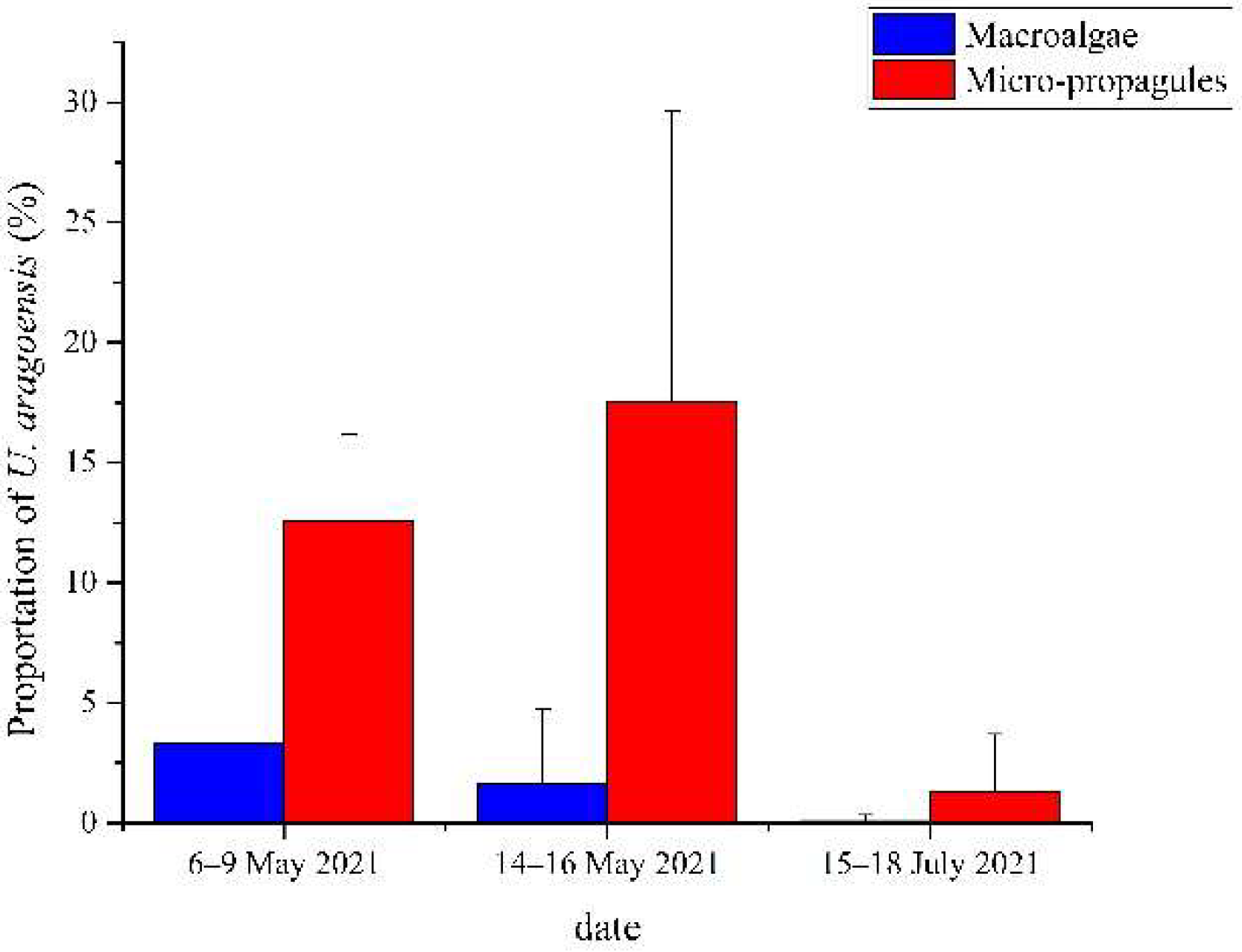
3.2. Proportion of U. aragoensis at Each Site
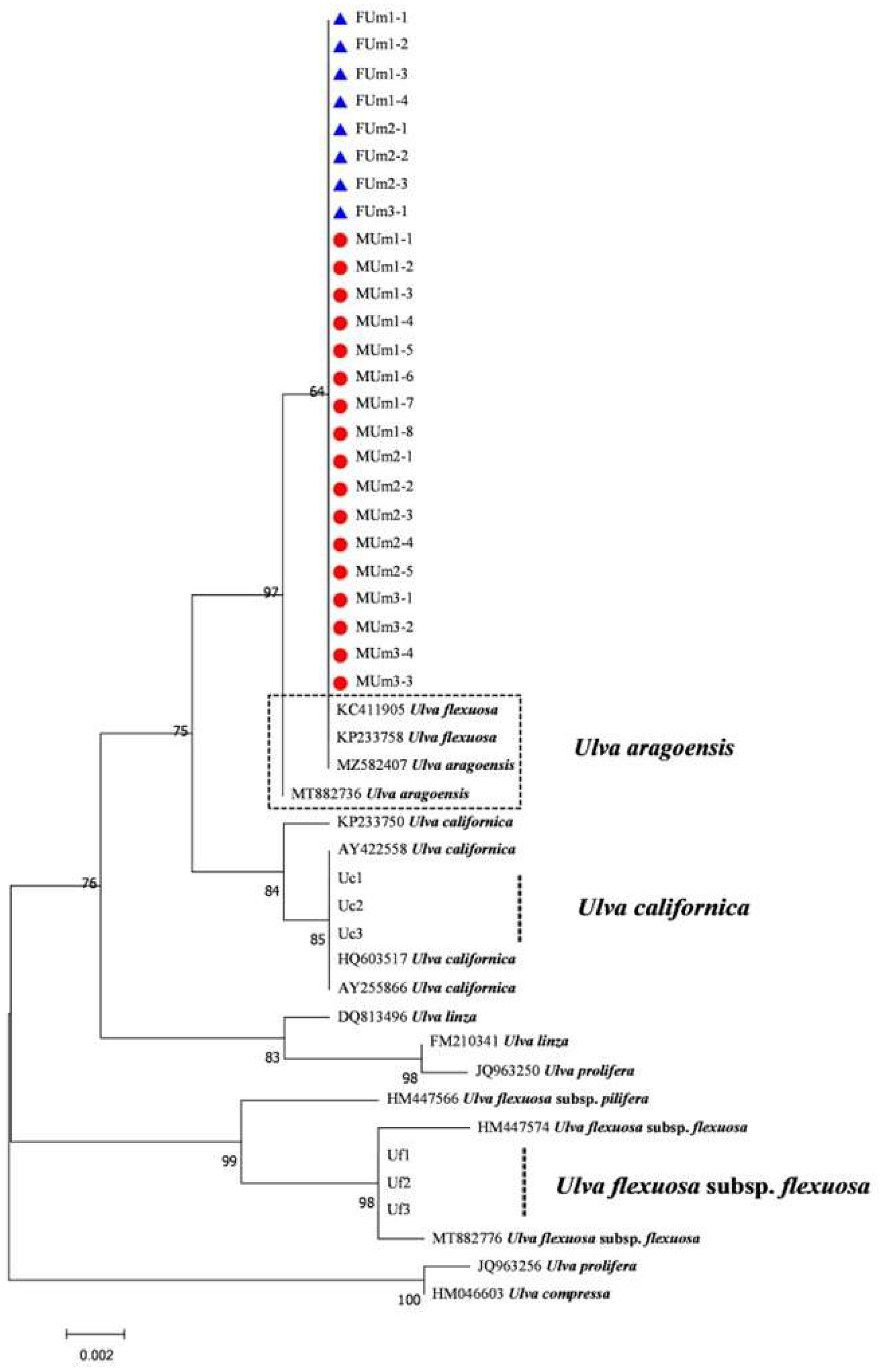
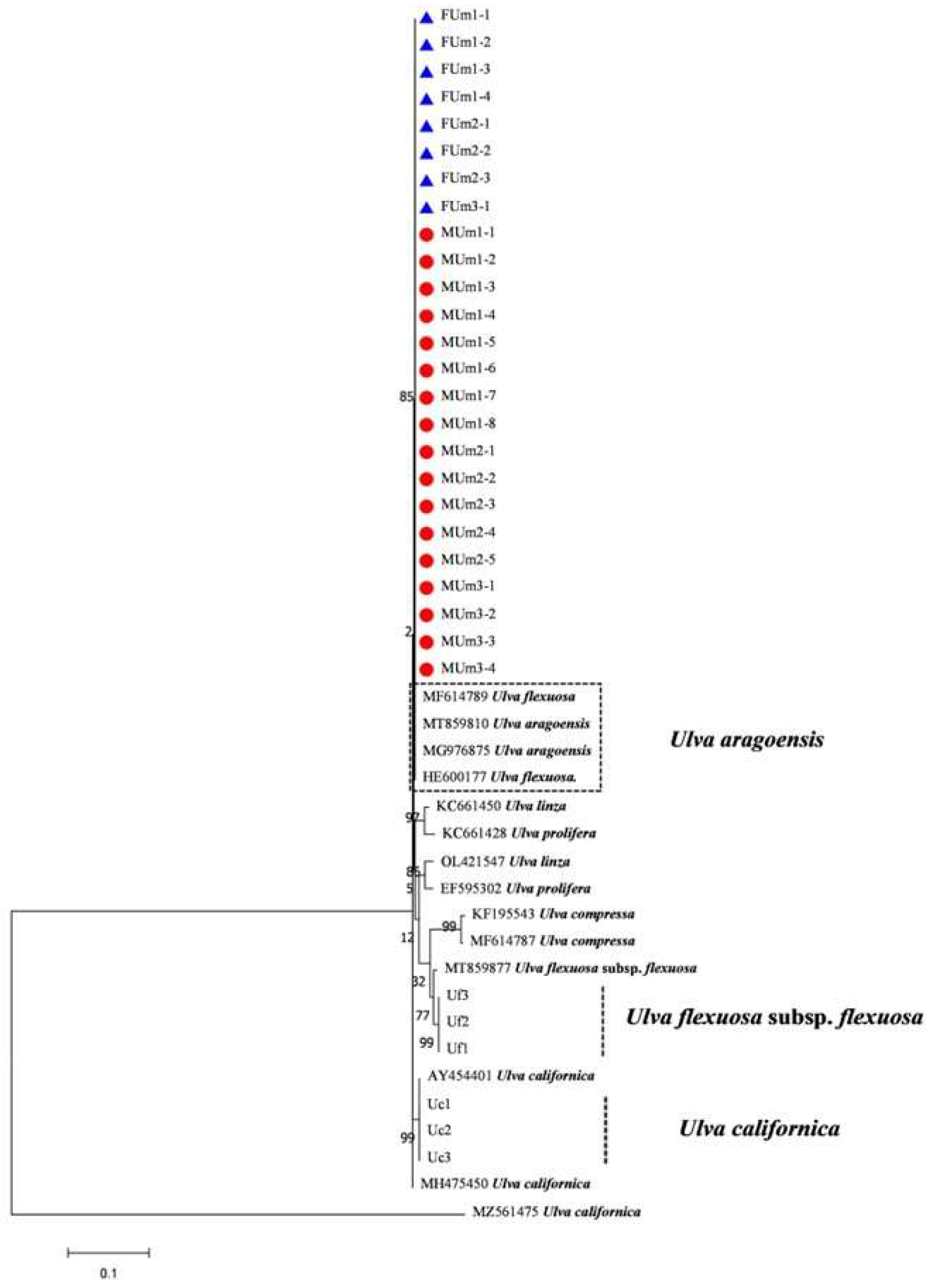
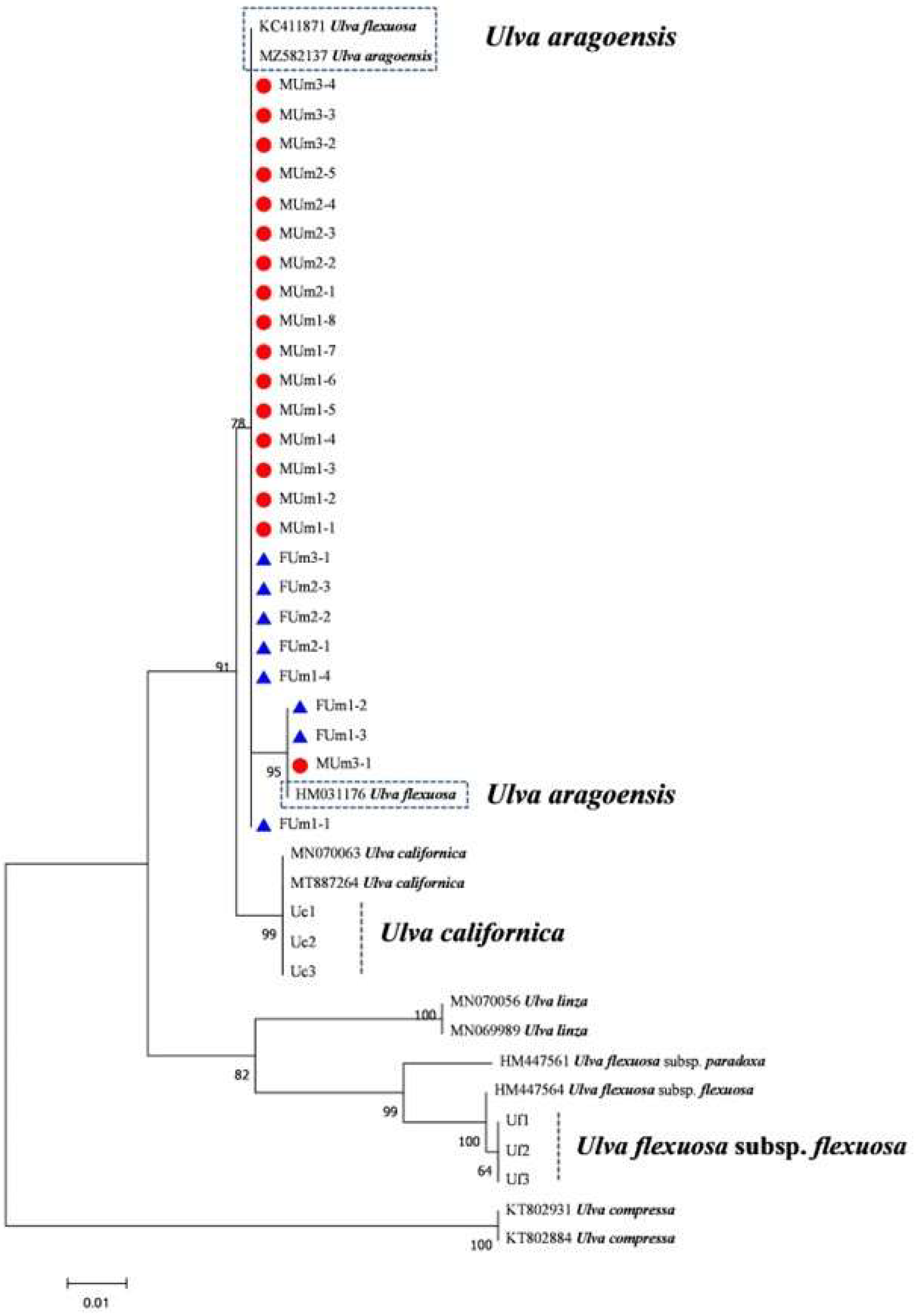
3.3. Phylogenetic Tree Results of Each DNA Barcode
3.4. Analysis of Haplotype Relationship between Macroalgae and Micro-Propagules
4. Discussion
4.1. Ulva Aragoensis Is a Constituent Species of the Early Green Tide in the SYS
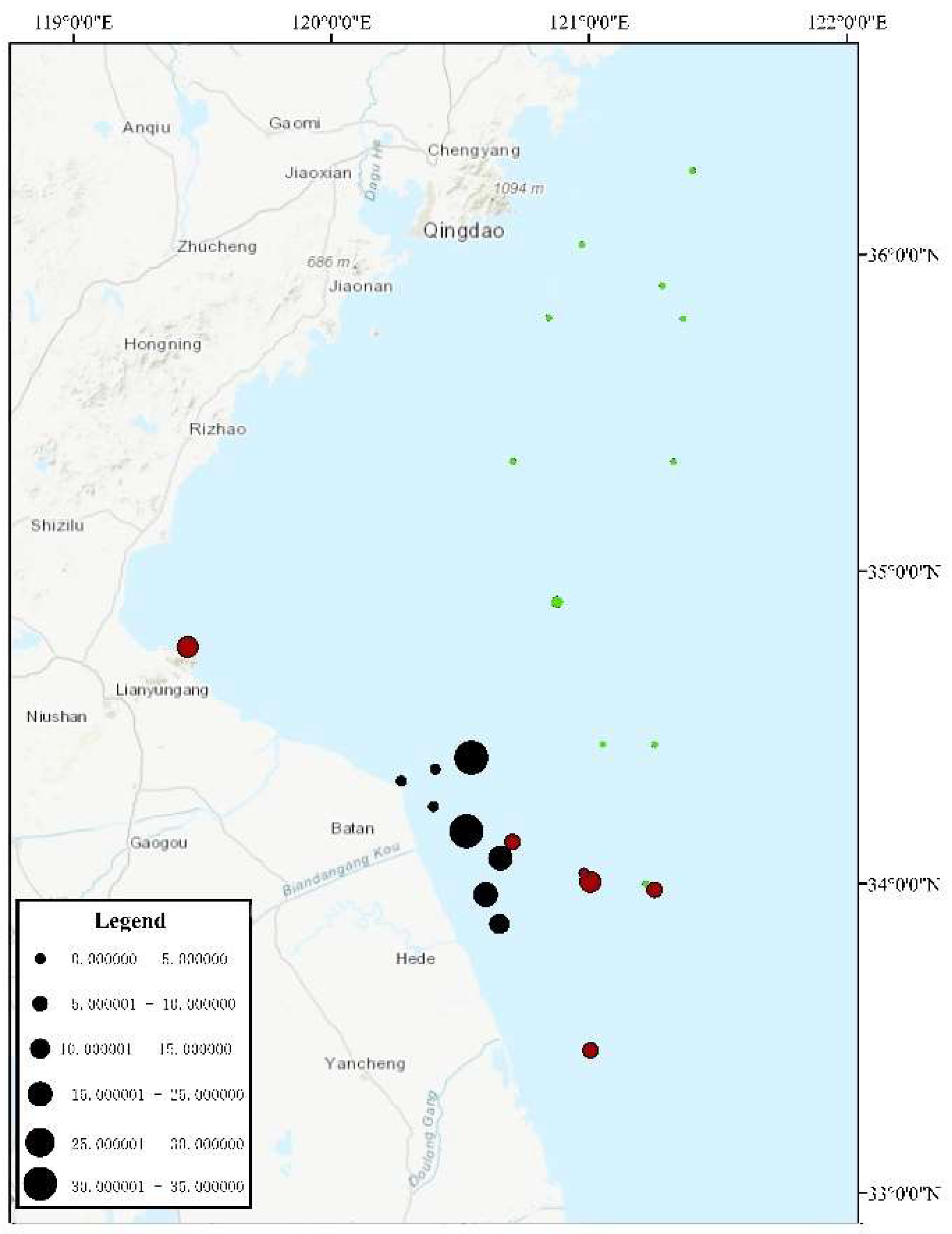
4.2. The Proportion of U. aragoensis Decreased as the Green Tide Moved Northward
4.3. Appropriate Molecular Markers for Green Tide Macroalgae Identification
4.4. The Micro-Propagule U. aragoensis and the Floating Green Tide U. aragoensis Are Related
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taylor, R.; Fletcher, R.L.; Raven, J.A. Preliminary Studies on the Growth of Selected ‘Green Tide’ Algae in Laboratory Culture: Effects of Irradiance, Temperature, Salinity and Nutrients on Growth Rate. Bot. Mar. 2001, 44, 327–336. [Google Scholar] [CrossRef]
- Ye, N.H.; Zhang, X.W.; Mao, Y.Z.; Liang, C.W.; Xu, D.; Zou, J.; Zhuang, Z.M.; Wang, Q.Y. ‘Green Tides’ are Overwhelming the Coastline of Our Blue Planet: Taking the World’s Largest Example. Ecol. Res. 2011, 26, 477–485. [Google Scholar] [CrossRef]
- Schreyers, L.; Van Emmerik, T.; Biermann, L.; Le Lay, Y.F. Spotting Green Tides over Brittany from Space: Three Decades of Monitoring with Landsat Imagery. Remote Sens. 2021, 13, 1308. [Google Scholar] [CrossRef]
- Lagourgue, L.; Gobin, S.; Brisset, M.; Vandenberghe, S.; Bonneville, C.; Jauffrais, T.; Van Wynsberge, S.; Payri, C.E. Ten New Species of Ulva (Ulvophyceae, Chlorophyta) Discovered in New Caledonia: Genetic and Morphological Diversity, and Bloom Potential. Eur. J. Phycol. 2022, 57, 458–478. [Google Scholar] [CrossRef]
- Song, W.; Han, H.B.; Wang, Z.L.; Li, Y. Molecular Identification of the Macroalgae That Cause Green Tides in the Bohai Sea, China. Aquat. Bot. 2019, 156, 38–46. [Google Scholar] [CrossRef]
- Wang, J.F.; Li, N.; Jiang, P.; Boo, S.M.; Lee, W.J.; Cui, Y.L.; Lin, H.Z.; Zhao, J.; Liu, Z.Y.; Qin, S. Ulva and Enteromorpha (Ulvaceae, Chlorophyta) from Two Sides of the Yellow Sea: Analysis of Nuclear rDNA ITS and Plastid rbcL Sequence Data. Chin. J. Oceanol. Limnol. 2010, 28, 762–768. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.H.; Shi, J.T.; Gao, S.; Huo, Y.Z.; Cui, J.J.; Shen, H.; Liu, G.Y.; He, P.M. Annual Patterns of Macroalgal Blooms in the Yellow Sea during 2007–2017. PLoS ONE 2019, 14, e0210460. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Jiang, P.; Liu, Z.Y.; Wei, W.; Lin, H.Z.; Li, F.C.; Wang, J.F.; Qin, S. The Yellow Sea Green Tides Were Dominated by One Species, Ulva (Enteromorpha) prolifera, from 2007 to 2011. Chin. Sci. Bull. 2013, 58, 2298–2302. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Y.; Huo, Y.Z.; Zhang, J.H.; Cui, J.J.; Wang, Y.; Yang, L.L.; Zhou, Q.Y.; Lu, Y.W.; Yu, K.F.; He, P.M. Variations of Dominant Free-Floating Ulva Species in the Source Area for the World’s Largest Macroalgal Blooms, China: Differences of Ecological Tolerance. Harmful Algae 2018, 74, 58–66. [Google Scholar] [CrossRef]
- Han, W.; Chen, L.P.; Zhang, J.H.; Tian, X.L.; Hua, L.; He, Q.; Huo, Y.Z.; Yu, K.F.; Shi, D.J.; Ma, J.H.; et al. Seasonal Variation of Dominant Free-Floating and Attached Ulva Species in Rudong Coastal Area, China. Harmful Algae 2013, 28, 46–54. [Google Scholar] [CrossRef]
- Huo, Y.Z.; Zhang, J.H.; Chen, L.P.; Hu, M.; Yu, K.F.; Chen, Q.F.; He, Q.; He, P.M. Green Algae Blooms Caused by Ulva prolifera in the Southern Yellow Sea: Identification of the Original Bloom Location and Evaluation of Biological Processes Occurring during the Early Northward Floating Period. Limnol. Oceanogr. 2013, 58, 2206–2218. [Google Scholar] [CrossRef]
- Liu, F.; Pang, S.J.; Xu, N.; Shan, T.F.; Sun, S.; Hu, X.A.; Yang, J.Q. Ulva Diversity in the Yellow Sea during the Large-Scale Green Algal Blooms in 2008–2009. Phycol. Res. 2010, 58, 270–279. [Google Scholar] [CrossRef]
- Gao, G.; Zhong, Z.H.; Zhou, X.H.; Xu, J.T. Changes in Morphological Plasticity of Ulva prolifera under Different Environmental Conditions: A Laboratory Experiment. Harmful Algae 2016, 59, 51–58. [Google Scholar] [CrossRef]
- Leliaert, F.; Zhang, X.W.; Ye, N.H.; Malta, E.; Engelen, A.H.; Mineur, F.; Verbruggen, H.; De Clerck, O. Research Note: Identity of the Qingdao Algal Bloom. Phycol. Res. 2009, 57, 147–151. [Google Scholar] [CrossRef]
- Saunders, G.W.; Kucera, H. An Evaluation of rbcL, tufA, UPA, LSU and ITS as DNA Barcode Markers for the Marine Green Macroalgae. Cryptogam. Algol. 2010, 31, 487–528. [Google Scholar]
- Shimada, S.; Yokoyama, N.; Arai, S.; Hiraoka, M. Phylogeography of the Genus Ulva (Ulvophyceae, Chlorophyta), with Special Reference to the Japanese Freshwater and Brackish Taxa. J. Appl. Phycol. 2008, 20, 979–989. [Google Scholar] [CrossRef]
- Liu, J.L.; Zhao, X.H.; Kang, X.Y.; Zhuang, M.M.; Ding, X.W.; Zhao, L.J.; Wen, Q.L.; Zhu, Y.; Gu, K.; Bao, Q.J.; et al. Good News: We Can Identify Ulva Species Erupted in the Yellow Sea More Easily and Cheaply Now. Conserv. Genet. Resour. 2020, 12, 447–449. [Google Scholar] [CrossRef]
- Kang, J.H.; Jang, J.E.; Kim, J.H.; Byeon, S.Y.; Kim, S.; Choi, S.K.; Kang, Y.H.; Park, S.R.; Lee, H.J. Species Composition, Diversity, and Distribution of the Genus Ulva along the Coast of Jeju Island, Korea Based on Molecular Phylogenetic Analysis. PLoS ONE 2019, 14, e0219958. [Google Scholar] [CrossRef] [Green Version]
- Hiraoka, M.; Ichihara, K.; Zhu, W.R.; Shimada, S.; Oka, N.; Cui, J.J.; Tsubaki, S.; He, P.M. Examination of Species Delimitation of Ambiguous DNA-Based Ulva (Ulvophyceae, Chlorophyta) Clades by Culturing and Hybridisation. Phycologia 2017, 56, 517–532. [Google Scholar] [CrossRef]
- Hiraoka, M.; Shimada, S.; Uenosono, M.; Masuda, M. A New Green-Tide-Forming Alga, Ulva ohnoi Hiraoka et Shimada sp. nov. (Ulvales, Ulvophyceae) from Japan. Phycol. Res. 2004, 52, 17–29. [Google Scholar] [CrossRef]
- Ichihara, K.; Arai, S.; Uchimura, M.; Fay, E.J.; Ebata, H.; Hiraoka, M.; Shimada, S. New Species of Freshwater Ulva, Ulva limnetica (Ulvales, Ulvophyceae) from the Ryukyu Islands, Japan. Phycol. Res. 2009, 57, 94–103. [Google Scholar] [CrossRef]
- Cui, J.J.; Monotilla, A.P.; Zhu, W.R.; Takano, Y.; Shimada, S.; Ichihara, K.; Matsui, T.; He, P.M.; Hiraoka, M. Taxonomic Reassessment of Ulva prolifera (Ulvophyceae, Chlorophyta) Based on Specimens from the Type Locality and Yellow Sea Green Tides. Phycologia 2018, 57, 692–704. [Google Scholar] [CrossRef]
- Krupnik, N.; Rinkevich, B.; Paz, G.; Douek, J.; Lewinsohn, E.; Israel, A.; Carmel, N.; Mineur, F.; Maggs, C.A. Native, Invasive and Cryptogenic Ulva Species from the Israeli Mediterranean Sea: Risk and Potential. Mediterr. Mar. Sci. 2018, 19, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Shimada, S.; Hiraoka, M.; Nabata, S.; Iima, M.; Masuda, M. Molecular Phylogenetic Analyses of the Japanese Ulva and Enteromorpha (Ulvales, Ulvophyceae), with Special Reference to the Free-Floating Ulva. Phycol. Res. 2003, 51, 99–108. [Google Scholar] [CrossRef]
- Hayden, H.S.; Blomster, J.; Maggs, C.A.; Silva, P.C.; Stanhope, M.J.; Waaland, J.R. Linnaeus Was Right All along: Ulva and Enteromorpha Are Not Distinct Genera. Eur. J. Phycol. 2003, 38, 277–294. [Google Scholar] [CrossRef]
- Melton, J.T.; Lopez-Bautista, J.M. Diversity of the Green Macroalgal Genus Ulva (Ulvophyceae, Chlorophyta) from the East and Gulf Coast of the United States Based on Molecular Data. J. Phycol. 2021, 57, 551–568. [Google Scholar] [CrossRef]
- Liu, D.Y.; Keesing, J.K.; Xing, Q.U.; Shi, P. World’s Largest Macroalgal Bloom Caused by Expansion of Seaweed Aquaculture in China. Mar. Pollut. Bull. 2009, 58, 888–895. [Google Scholar] [CrossRef]
- Huo, Y.Z.; Han, H.B.; Hua, L.; Wei, Z.L.; Yu, K.F.; Shi, H.H.; Kim, J.K.; Yarish, C.; He, P.M. Tracing the Origin of Green Macroalgal Blooms Based on the Large Scale Spatio-Temporal Distribution of Ulva Microscopic Propagules and Settled Mature Ulva Vegetative Thalli in Coastal Regions of the Yellow Sea, China. Harmful Algae 2016, 59, 91–99. [Google Scholar] [CrossRef]
- Miao, X.X.; Xiao, J.; Xu, Q.Z.; Fan, S.L.; Wang, Z.L.; Wang, X.; Zhang, X.L. Distribution and Species Diversity of the Floating Green Macroalgae and Micro-Propagules in the Subei Shoal Southwestern Yellow Sea. PeerJ 2020, 8, e10538. [Google Scholar] [CrossRef]
- Song, W.; Li, Y.; Fang, S.; Wang, Z.L.; Xiao, J.; Li, R.X.; Fu, M.Z.; Zhu, M.Y.; Zhang, X.L. Temporal and Spatial Distributions of Green Algae Micro-Propagules in the Coastal Waters of the Subei Shoal, China. Estuar. Coast. Shelf Sci. 2015, 163, 29–35. [Google Scholar] [CrossRef]
- Huo, Y.Z.; Hua, L.; Wu, H.L.; Zhang, J.H.; Cui, J.J.; Huang, X.W.; Yu, K.F.; Shi, H.H.; He, P.M.; Ding, D.W. Abundance and Distribution of Ulva Microscopic Propagules Associated with a Green Tide in the Southern Coast of the Yellow Sea. Harmful Algae 2014, 39, 357–364. [Google Scholar] [CrossRef]
- Coat, G.; Dion, P.; Noailles, M.C.; De-Reviers, B.; Fontaine, J.M.; Berger-Perrot, Y.; Loiseaux-De, G.S. Ulva armoricana (Ulvales, Chlorophyta) from the Coasts of Brittany (France). I. Morphological Identification. Eur. J. Phycol. 1998, 33, 81–86. [Google Scholar] [CrossRef]
- Tian, X.L.; Huo, Y.Z.; Chen, L.P.; He, J.H.; Zhang, J.H.; Jia, R.; Liu, H.; Wang, J.H.; Xu, R.; Yang, J.Q.; et al. Molecular Detection and Analysis of Green Seaweeds from Rudong Coasts in Jiangsu Province. Chin. Sci. Bull. 2011, 56, 309–317. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X Windows Interface: Flexible Strategies for Multiple Sequence Alignment Aided by Quality Analysis Tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Leskinen, E.; Pamilo, P. Evolution of the ITS Sequences of Ribosomal DNA in Enteromorpha (Chlorophyceae). Hereditas 1997, 126, 17–23. [Google Scholar] [CrossRef]
- Fama, P.; Wysor, B.; Kooistra, W.; Zuccarello, G.C. Molecular Phylogeny of the Genus Caulerpa (Caulerpales, Chlorophyta) Inferred from Chloroplast tufA Gene. J. Phycol. 2002, 38, 1040–1050. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.H.; Shen, S.D.; Chen, W.Z.; Li, H.H. Phylogenetic Analyses of Four Species of Ulva and Monostroma grevillei Using ITS, rbcL and 18s rDNA Sequence Data. Chin. J. Oceanol. Limnol. 2013, 31, 97–105. [Google Scholar] [CrossRef]
- Wang, J.F.; Jiang, P.; Cui, Y.L.; Li, N.; Wang, M.Q.; Lin, H.Z.; He, P.M.; Qin, S. Molecular Analysis of Green-Tide-Forming Macroalgae in the Yellow Sea. Aquat. Bot. 2010, 93, 25–31. [Google Scholar] [CrossRef]
- Manhart, J.R. Phylogenetic Analysis of Green Plant rbcL Sequences. Mol. Phylogenet. Evol. 1994, 3, 114–127. [Google Scholar] [CrossRef]
- Liu, J.L.; Tong, Y.C.; Xia, J.; Sun, Y.Q.; Zhao, X.H.; Sun, J.Y.; Zhao, S.; Zhuang, M.M.; Zhang, J.H.; He, P.M. Ulva Macroalgae within Local Aquaculture Ponds along the Estuary of Dagu River, Jiaozhou Bay, Qingdao. Mar. Pollut. Bull. 2021, 174, 113243. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A Software for Comprehensive Analysis of DNA Polymorphism Data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Leigh, J.W.; Bryant, D. Popart: Full-Feature Software for Haplotype Network Construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Huo, Y.Z.; Han, H.B.; Shi, H.H.; Wu, H.L.; Zhang, J.H.; Yu, K.F.; Xu, R.; Liu, C.C.; Zhang, Z.L.; Liu, K.F.; et al. Changes to the Biomass and Species Composition of Ulva sp. on Porphyra Aquaculture Rafts, along the Coastal Radial Sandbank of the Southern Yellow Sea. Mar. Pollut. Bull. 2015, 93, 210–216. [Google Scholar] [CrossRef]
- Sun, Y.Q.; Yao, L.L.; Liu, J.L.; Tong, Y.C.; Xia, J.; Zhao, X.H.; Zhao, S.; Fu, M.L.; Zhuang, M.M.; He, P.M.; et al. Prevention Strategies for Green Tides at Source in the Southern Yellow Sea. Mar. Pollut. Bull. 2022, 178, 113646. [Google Scholar] [CrossRef]
- Xia, Z.Y.; Yuan, H.Q.; Liu, J.L.; Sun, Y.Q.; Tong, Y.C.; Zhao, S.; Xia, J.; Li, S.; Hu, M.J.; Cao, J.X.; et al. A Review of Physical, Chemical, and Biological Green Tide Prevention Methods in the Southern Yellow Sea. Mar. Pollut. Bull. 2022, 180, 113772. [Google Scholar] [CrossRef]
- Wang, Z.L.; Xiao, J.; Fan, S.L.; Li, Y.; Liu, X.Q.; Liu, D.Y. Who Made the World’s Largest Green Tide in China?—An Integrated Study on the Initiation and Early Development of the Green Tide in Yellow Sea. Limnol. Oceanogr. 2015, 60, 1105–1117. [Google Scholar] [CrossRef]
- Geng, H.X.; Yan, T.; Zhou, M.J.; Liu, Q. Comparative Study of the Germination of Ulva prolifera Gametes on Various Substrates. Estuar. Coast. Shelf Sci. 2015, 163, 89–95. [Google Scholar] [CrossRef]
- Ma, Y.F.; Wong, K.P.; Tsou, J.Y.; Zhang, Y.Z. Investigating Spatial Distribution of Green-Tide in the Yellow Sea in 2021 Using Combined Optical and SAR Images. J. Mar. Sci. Eng. 2022, 10, 127. [Google Scholar] [CrossRef]
- Wang, B.; Wu, L.; Zhao, N.; Liu, T.R.; Hirose, N. Summer Wind Effects on Coastal Upwelling in the Southwestern Yellow Sea. J. Mar. Sci. Eng. 2021, 9, 1021. [Google Scholar] [CrossRef]
- Song, M.J.; Kong, F.Z.; Li, Y.F.; Zhao, J.; Yu, R.C.; Zhou, M.J.; Jiang, P.; Yan, T. A Massive Green Tide in the Yellow Sea in 2021: Field Investigation and Analysis. Int. J. Environ. Res. Public Health 2022, 19, 11753. [Google Scholar] [CrossRef]













| Sample No. | Station | Type |
|---|---|---|
| FUm1-1 | A1 | Macroalgae |
| FUm1-2 | A1 | Macroalgae |
| FUm1-3 | A2 | Macroalgae |
| FUm1-4 | A3 | Macroalgae |
| FUm2-1 | B1 | Macroalgae |
| FUm2-2 | B2 | Macroalgae |
| FUm2-3 | B3 | Macroalgae |
| FUm3-1 | S9 | Macroalgae |
| MUm1-1 | A1 | Micro-propagules |
| MUm1-2 | A1 | Micro-propagules |
| MUm1-3 | A1 | Micro-propagules |
| MUm1-4 | A3 | Micro-propagules |
| MUm1-5 | A5 | Micro-propagules |
| MUm1-6 | A5 | Micro-propagules |
| MUm1-7 | A5 | Micro-propagules |
| MUm1-8 | A6 | Micro-propagules |
| MUm2-1 | B4 | Micro-propagules |
| MUm2-2 | B5 | Micro-propagules |
| MUm2-3 | B5 | Micro-propagules |
| MUm2-4 | B7 | Micro-propagules |
| MUm2-5 | B7 | Micro-propagules |
| MUm3-1 | S7 | Micro-propagules |
| MUm3-2 | S8 | Micro-propagules |
| MUm3-3 | S9 | Micro-propagules |
| MUm3-4 | S9 | Micro-propagules |
| DNA Makers | rbcL | tufA | rps2-trnL |
|---|---|---|---|
| Accession in NCBI | KX579943 Ulva flexuosa | HE600177 Ulva flexuosa | KY626326 Ulva flexuosa |
| KP233758 Ulva flexuosa | MH475451 Ulva flexuosa | KX455878 Ulva flexuosa | |
| KC411905 Ulva flexuosa | OL604529 Ulva flexuosa | MH013470 Ulva flexuosa | |
| KC411904 Ulva flexuosa | MH475452 Ulva flexuosa | ||
| KC411892 Ulva flexuosa | |||
| KP233759 Ulva flexuosa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, Y.; Xia, L.; Liu, J.; Zhao, S.; Sun, Y.; Wu, T.; Xia, Z.; Li, S.; Cao, J.; Zhang, J. Distribution and Identification of Ulva aragoensis (Ulvaceae, Chlorophyta), a Constituent Species of Green Tides in the Southern Yellow Sea, Based on Molecular Data. J. Mar. Sci. Eng. 2022, 10, 1767. https://doi.org/10.3390/jmse10111767
Tong Y, Xia L, Liu J, Zhao S, Sun Y, Wu T, Xia Z, Li S, Cao J, Zhang J. Distribution and Identification of Ulva aragoensis (Ulvaceae, Chlorophyta), a Constituent Species of Green Tides in the Southern Yellow Sea, Based on Molecular Data. Journal of Marine Science and Engineering. 2022; 10(11):1767. https://doi.org/10.3390/jmse10111767
Chicago/Turabian StyleTong, Yichao, Lihua Xia, Jinlin Liu, Shuang Zhao, Yuqing Sun, Tingjian Wu, Zhangyi Xia, Shuang Li, Jiaxing Cao, and Jianheng Zhang. 2022. "Distribution and Identification of Ulva aragoensis (Ulvaceae, Chlorophyta), a Constituent Species of Green Tides in the Southern Yellow Sea, Based on Molecular Data" Journal of Marine Science and Engineering 10, no. 11: 1767. https://doi.org/10.3390/jmse10111767
APA StyleTong, Y., Xia, L., Liu, J., Zhao, S., Sun, Y., Wu, T., Xia, Z., Li, S., Cao, J., & Zhang, J. (2022). Distribution and Identification of Ulva aragoensis (Ulvaceae, Chlorophyta), a Constituent Species of Green Tides in the Southern Yellow Sea, Based on Molecular Data. Journal of Marine Science and Engineering, 10(11), 1767. https://doi.org/10.3390/jmse10111767