
Growth and Physiological Responses of Chrysotila roscoffensis (Coccolithales) to Different Phosphorus Resources

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Strain
2.2. Experimental Design
2.3. Oxygen Production and the Chlorophyll Fluorescence Parameters
2.4. Activity of Cellular Alkaline Phosphatases (AP)
2.5. Cellular Calcification
2.6. Variations of Nitrate and Phosphate Concentration in the Water
2.7. Data Analysis
3. Results
3.1. Population Growth
3.2. Variations in the Chlorophyll Fluorescence Parameters and Oxygen Production
3.3. Algal Alkaline Phosphatase (AP) Activity Variations
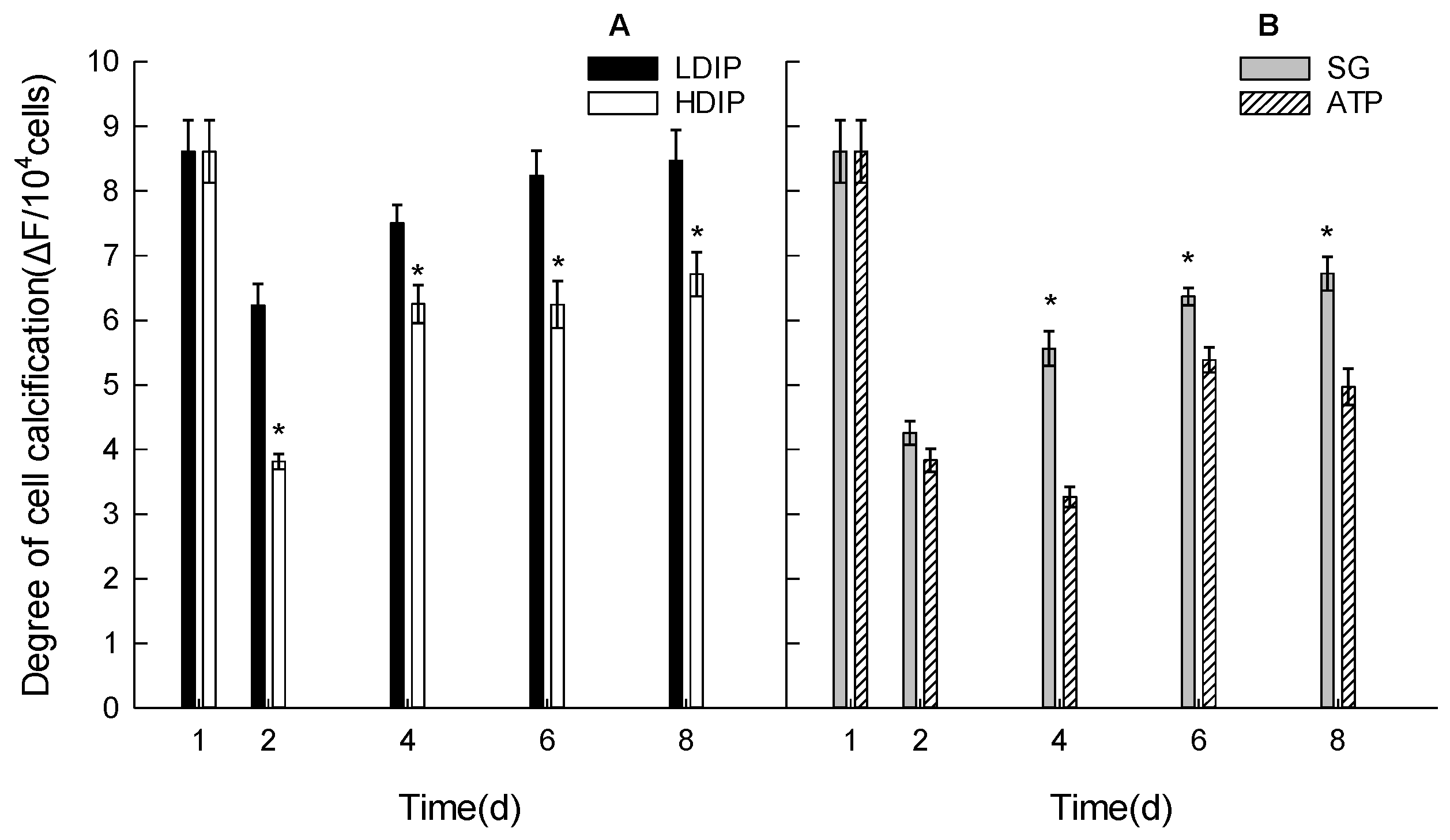
3.4. Extracellular Calcification Variations
3.5. Variation in Nitrate and Phosphate Concentrations and the N/P Ratio in the Water
4. Discussion
4.1. Population Growth
4.2. Nitrate and Phosphate Variations in the Water
4.3. AP Activity
4.4. Photosynthetic Activity and Cellular Calcification
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, S.J.; Litaker, R.W.; Sunda, W.G. Phosphorus physiological ecology and molecular mechanisms in marine phytoplankton. J. Phycol. 2016, 52, 10–36. [Google Scholar] [CrossRef] [PubMed]
- Cembella, A.D.; Antia, N.J.; Harrison, P.J. The utilization of inorganic and organic phosphorous compounds as nutrients by eukaryotic microalgae: A multidisciplinary perspective: Part I. Crit. Rev. Microbiol. 1982, 10, 317–391. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Nelson, C.R.; Karl, D.M. Phosphorus cycling in the North Pacific Subtropical Gyre using cosmogenic 32P and 33P. Limnol. Oceanogr. 2002, 47, 762–770. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.H.; Liang, Y.; Kang, W. Utilization of dissolved organic phosphorus by different groups of phytoplankton taxa. Harmful Algae 2011, 12, 113–118. [Google Scholar] [CrossRef]
- Ghyoot, C.; Gypens, N.; Flynn, K.J.; Lancelot, C. Modelling alkaline phosphatase activity in microalgae under orthophosphate limitation: The case of Phaeocystis globosa. J. Plankton Res. 2015, 37, 869–885. [Google Scholar] [CrossRef] [Green Version]
- Ammerman, J.W. Role of Ecto-phosphohydrolases in phosphorus regeneration in estuarine and coastal ecosystems. In Microbial Enzymes in Aquatic Environments; Springer: New York, NY, USA, 1991; pp. 165–186. [Google Scholar]
- Tiwari, B.; Singh, S.; Kaushik, M.S.; Mishra, A.K. Regulation of organophosphate metabolism in cyanobacteria. A review. Microbiology 2015, 84, 291–302. [Google Scholar] [CrossRef]
- Xie, E.; Su, Y.P.; Deng, S.Q.; Kontopyrgou, M.; Zhang, D.Y. Significant influence of phosphorus resources on the growth and alkaline phosphatase activities of Microcystis aeruginosa. Environ. Pollut. 2021, 268, 115807. [Google Scholar] [CrossRef]
- Xie, C.S.; Lu, R.J.; Huang, Y.; Wang, Q.; Xu, X.H. Effects of ions and phosphates on alkaline phosphatase activity in aerobic activated sludge system. Bioresour. Technol. 2010, 101, 3394–3399. [Google Scholar] [CrossRef]
- Willsky, G.R.; Bennett, R.L.; Malamy, M.H. Inorganic phosphate transport in Escherichia coli: Involvement of two genes which play a role in alkaline phosphatase regulation. J. Bacteriol. 1973, 113, 529–539. [Google Scholar] [CrossRef] [Green Version]
- Nausch, M. Alkaline phosphatase activities and the relationship to inorganic phosphate in the Pomeranian Bight (southern Baltic Sea). Aquat. Microb. Ecol. 1998, 16, 87–94. [Google Scholar] [CrossRef]
- Shi, X.L.; Qian, S.Q.; Kong, F.X.; Zhang, M.; Yu, Y. Differences in growth and alkaline phosphatase activity between Microcystis aeruginosa and Chlorella pyrenoidosa in response to media with different organic phosphorus. J. Limnol. 2011, 70, 21–25. [Google Scholar] [CrossRef]
- Bopp, L.; Resplandy, L.; Orr, J.C.; Doney, S.C.; Dunne, J.P.; Gehlen, M.; Halloran, P.; Heinze, C.; Ilyina, T.; Séférian, R.; et al. Multiple stressors of ocean ecosystems in the 21st century: Projections with CMIP5 models. Biogeosciences 2013, 10, 6225–6245. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Peng, R.H.; Yang, Y.; He, L.; Wang, W.Q.; Zheng, T.L.; Lin, G.H. Mariculture pond influence on mangrove areas in south China: Significantly larger nitrogen and phosphorus loadings from sediment wash-out than from tidal water exchange. Aquaculture 2014, 426, 204–212. [Google Scholar] [CrossRef]
- Deininger, A.; Frigstad, H. Reevaluating the role of organic matter sources for coastal eutrophication, oligotrophication, and ecosystem health. Front. Mar. Sci. 2019, 6, 210. [Google Scholar]
- Wang, Y.J.; Liu, D.Y.; Xiao, W.P.; Zhou, P.; Tian, C.G.; Zhang, C.S.; Du, J.Z.; Guo, H.; Wang, B.D. Coastal eutrophication in China: Trend, sources, and ecological effects. Harmful Algae 2021, 107, 102058. [Google Scholar] [CrossRef]
- Karl, D.M.; Björkman, K.M. Phosphorus cycle in seawater: Dissolved and particulate pool inventories and selected phosphorus fluxes. Methods Microbiol. 2001, 30, 239–270. [Google Scholar]
- Billen, G.; Garnier, J.; Rousseau, V. Nutrient fluxes and water quality in the drainage network of the Scheldt basin over the last 50 years. Hydrobiologia 2005, 540, 47–67. [Google Scholar] [CrossRef]
- Andersen, R.A.; Kim, J.I.; Tittley, I.; Yoon, H.S. A re-investigation of Chrysotila (Prymnesiophyceae) using material collected from the type locality. Phycologia 2014, 53, 463–473. [Google Scholar] [CrossRef]
- Fukuzaki, K.; Imai, I.; Fukushima, K.; Ishii, K.I.; Sawayama, S.; Yoshioka, T. Fluorescent characteristics of dissolved organic matter produced by bloom-forming coastal phytoplankton. J. Plankton Res. 2014, 36, 685–694. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.X.; Jiang, Y.; Liu, B.N.; Yan, X.J.; Zhang, W.D. The relationship between calcification and photosynthesis in the coccolithophorid Pleurochrysis carterae. Acta Ecol. Sin. 2012, 32, 38–43. [Google Scholar] [CrossRef]
- Reifel, K.M.; McCoy, M.P.; Tiffany, M.A.; Rocke, T.E.; Trees, C.C.; Barlow, S.B.; Faulkner, D.J.; Hurlbert, S.H. Pleurochrysis pseudoroscoffensis (Prymnesiophyceae) blooms on the surface of the Salton Sea, California. Hydrobiologia 2001, 466, 177–185. [Google Scholar] [CrossRef]
- Bautista-Chamizo, E.; De Orte, M.R.; DelValls, T.Á.; Riba, I. Simulating CO2 leakages from CCS to determine Zn toxicity using the marine microalgae Pleurochrysis roscoffensis. Chemosphere 2016, 144, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Maria, P.; Evgeniy, G.; Boris, S.; Nikita, Z.; Anna, M.; Alla, F.; Maxim, K.; Yevhen, M.; Ilia, Y.; Elena, G.; et al. Fucoxanthin production by heterokont microalgae. Algal Res. 2017, 24, 387–393. [Google Scholar]
- Moheimani, N.R.; Webb, J.P.; Borowitzka, M.A. Bioremediation and other potential applications of coccolithophorid algae: A review. Algal Res. 2012, 1, 120–133. [Google Scholar] [CrossRef]
- Young, J.R. Functions of coccoliths. In Coccolithophores; Winter, A., Siesser, W.G., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 63–82. [Google Scholar]
- De Vrind-de Jong, E.W.; Borman, A.H.; Thierry, R.; Westbroek, P.W.; Cruter, M.; Kanerling, J.P. Calcification in the coccolithophorids Emiliania huxleyi and Pleurochrysis carterae II. Biochem. Aspects. In Bioremediation in Lower Plants and Animals; Leadbeater, B.S.C., Ed.; Clarendon Press: Oxford, UK, 1986; pp. 205–217. [Google Scholar]
- Siesser, W.G. Historical background of coccolithophore studies. In Coccolithophores; Winter, A., Siesser, W.G., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 51–62. [Google Scholar]
- Rost, B.; Riebesell, U. Coccolithophores and the biological pump: Responses to environmental changes. In Coccolithophores; Springer: Berlin, 2004; pp. 99–125. [Google Scholar]
- Kang, P.P.; Xu, S.G. The impact of mariculture on nutrient dynamics and identification of the nitrate sources in coastal waters. Environ. Sci. Pollut. Res. 2016, 23, 1300–1311. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Springer: Boston, MA, USA, 1975; pp. 29–60. [Google Scholar]
- Harrison, P.J.; Waters, R.E.; Taylor, F.J.R. A broad spectrum artificial sea water medium for coastal and open ocean phytoplankton. J. Phycol. 1980, 16, 28–35. [Google Scholar] [CrossRef]
- Roháček, K. Chlorophyll fluorescence parameters: The definitions, photosynthetic meaning, and mutual relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Fitzgerald, G.P.; Nelson, T.C. Extractive and enzymatic analyses for limiting or surplus phosphorus in algae. J. Phycol. 1975, 11, 32–37. [Google Scholar] [CrossRef]
- Hadas, O.; Pinkas, R. Arylsulfatase and alkaline phosphatase (Apase) activity in sediments of Lake Kinneret, Israel. In The Interactions Between Sediments and Water; Springer: Dordrecht, The Netherlands, 1997; pp. 671–679. [Google Scholar]
- Xu, Y.; Wahlund, T.M.; Feng, L.; Shaked, Y.; Morel, F.M. A novel alkaline phosphatase in the coccolithophore Emiliania huxleyi (Prymnesiophyceae) and its regulation by phosphorus. J. Phycol. 2006, 42, 835–844. [Google Scholar] [CrossRef]
- Oviedo, A.M.; Langer, G.; Ziveri, P. Effect of phosphorus limitation on coccolith morphology and element ratios in Mediterranean strains of the coccolithophore Emiliania huxleyi. J. Exp. Mar. Biol. Ecol. 2014, 459, 105–113. [Google Scholar] [CrossRef]
- Godrijan, J.; Young, J.R.; Marić Pfannkuchen, D.; Precali, R.; Pfannkuchen, M. Coastal zones as important habitats of coccolithophores: A study of species diversity, succession, and life-cycle phases. Limnol. Oceanogr. 2018, 63, 1692–1710. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, Y.; Park, S.E.; Kim, T.; Kim, B.; Kang, D.J.; Rho, T. Impact of aquaculture on distribution of dissolved organic matter in coastal Jeju Island, Korea, based on absorption and fluorescence spectroscopy. Environ. Sci. Pollut. Res. 2022, 29, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Dyhrman, S.T.; Ruttenberg, K.C. Presence and regulation of alkaline phosphatase activity in eukaryotic phytoplankton from the coastal ocean: Implications for dissolved organic phosphorus remineralization. Limnol. Oceanogr. 2006, 51, 1381–1390. [Google Scholar] [CrossRef]
- Riegman, R.; Stolte, W.; Noordeloos, A.A.; Slezak, D. Nutrient uptake and alkaline phosphatase (EC 3:1:3:1) activity of Emiliania huxleyi (Prymnesiophyceae) during growth under N and P limitation in continuous cultures. J. Phycol. 2000, 36, 87–96. [Google Scholar] [CrossRef]
- Li, D.; Liu, R.Q.; Cui, X.Y.; He, M.L.; Zheng, S.Y.; Du, W.J.; Gao, M.; Wang, C.H. Co-culture of bacteria and microalgae for treatment of high concentration biogas slurry. J. Water Process Eng. 2021, 41, 102014. [Google Scholar] [CrossRef]
- Xin, C.H.; Addy, M.M.; Zhao, J.Y.; Cheng, Y.L.; Cheng, S.B.; Mu, D.Y.; Liu, Y.H.; Ding, R.J.; Chen, P.; Ruan, R. Comprehensive techno-economic analysis of wastewater-based algal biofuel production: A case study. Bioresour. Technol. 2016, 211, 584–593. [Google Scholar] [CrossRef] [Green Version]
- Stockner, J.G.; Shortreed, K.S. Response of Anabaena and Synechococcus to manipulation of nitrogen: Phosphorus ratios in a lake fertilization experiment. Limnol. Oceanogr. 1988, 33, 1348–1361. [Google Scholar] [CrossRef]
- Cuvin-Aralar, M.L.; Focken, U.; Becker, K.; Aralar, E.V. Effects of low nitrogen-phosphorus ratios in the phytoplankton community in Laguna de Bay, a shallow eutrophic lake in the Philippines. Aquat. Ecol. 2004, 38, 387–401. [Google Scholar] [CrossRef]
- Zhang, T.X.; Lu, X.R.; Yu, R.D.; Qin, M.Y.; Wei, C.; Hong, S.J. Response of extracellular and intracellular alkaline phosphatase in Microcystis aeruginosa to organic phosphorus. Environ. Sci. Pollut. Res. 2020, 27, 42304–42312. [Google Scholar] [CrossRef]
- Chiaudani, G.; Vighi, M. Multistep approach to identification of limiting nutrients in Northern Adriatic eutrophied coastal waters. Water Res. 1982, 16, 1161–1166. [Google Scholar] [CrossRef]
- Quisel, J.D.; Wykoff, D.D.; Grossman, A.R. Biochemical characterization of the extracellular phosphatases produced by phosphorus-deprived Chlamydomonas reinhardtii. Plant Physiol. 1996, 111, 839–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rengefors, K.; Pettersson, K.; Blenckner TM, A.D.; Anderson, D.M. Species-specific alkaline phosphatase activity in freshwater spring phytoplankton: Application of a novel method. J. Plankton Res. 2001, 23, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Cañavate, J.P.; Armada, I.; Hachero-Cruzado, I. Aspects of phosphorus physiology associated with phosphate-induced polar lipid remodeling in marine microalgae. J. Plant Physiol. 2017, 214, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Rose, C.; Axler, R.P. Uses of alkaline phosphatase activity in evaluating phytoplankton community phosphorus deficiency. Hydrobiologia 1997, 361, 145–156. [Google Scholar] [CrossRef]
- Jamet, D.; Amblard, C.; Devaux, J. Size-fractionated alkaline phosphatase activity in the Hypereutrophic Villerest Reservoir (Roanne, France). Water Environ. Res. 2001, 73, 132–141. [Google Scholar] [CrossRef]
- Jin, X.B.; Liu, C.L.; Poulton, A.J.; Dai, M.H.; Guo, X.H. Coccolithophore responses to environmental variability in the South China Sea: Species composition and calcite content. Biogeosciences 2016, 13, 4843–4861. [Google Scholar] [CrossRef] [Green Version]
- Azam, F.; Hodson, R.E. Dissolved ATP in the sea and its utilization by marine bacteria. Nature 1977, 267, 696–698. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Šupraha, L.; Gerecht, A.C.; Probert, I.; Henderiks, J. Eco-physiological adaptation shapes the response of calcifying algae to nutrient limitation. Sci. Rep. 2015, 5, 16499. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Xiao, P.; Liu, B.; Han, J.; Li, Y.; Zhou, C. Growth and Physiological Responses of Chrysotila roscoffensis (Coccolithales) to Different Phosphorus Resources. J. Mar. Sci. Eng. 2022, 10, 1860. https://doi.org/10.3390/jmse10121860
Zhou L, Xiao P, Liu B, Han J, Li Y, Zhou C. Growth and Physiological Responses of Chrysotila roscoffensis (Coccolithales) to Different Phosphorus Resources. Journal of Marine Science and Engineering. 2022; 10(12):1860. https://doi.org/10.3390/jmse10121860
Chicago/Turabian StyleZhou, Li, Peng Xiao, Baoning Liu, Jichang Han, Yahe Li, and Chengxu Zhou. 2022. "Growth and Physiological Responses of Chrysotila roscoffensis (Coccolithales) to Different Phosphorus Resources" Journal of Marine Science and Engineering 10, no. 12: 1860. https://doi.org/10.3390/jmse10121860
APA StyleZhou, L., Xiao, P., Liu, B., Han, J., Li, Y., & Zhou, C. (2022). Growth and Physiological Responses of Chrysotila roscoffensis (Coccolithales) to Different Phosphorus Resources. Journal of Marine Science and Engineering, 10(12), 1860. https://doi.org/10.3390/jmse10121860