
Skin Culturable Microbiota in Farmed European Seabass (Dicentrarchuslabrax) in Two Aquacultures with and without Antibiotic Use

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
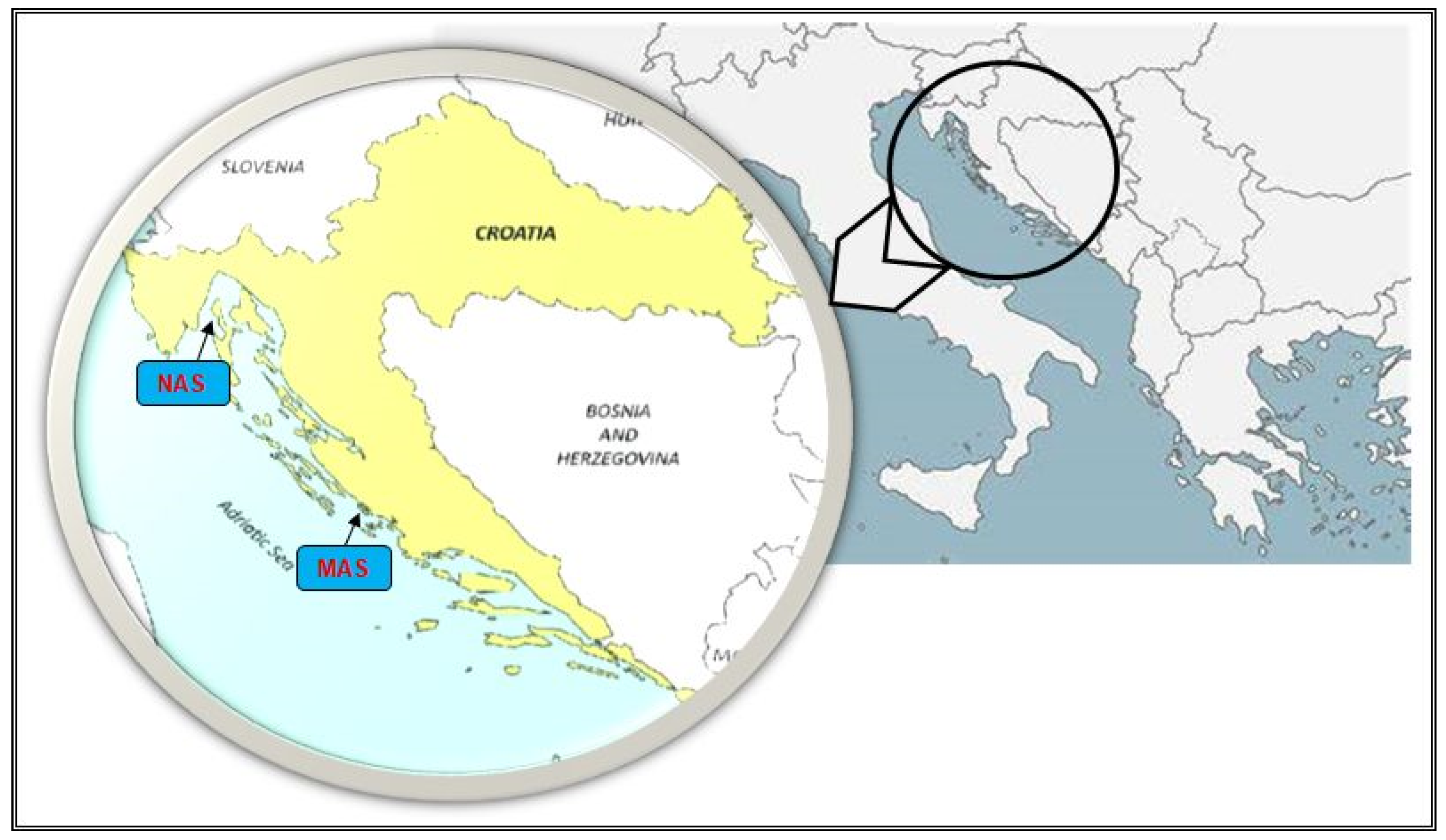
2.1. Sampling
2.2. Physicochemical and Microbiological Analysis of Seawater and Sediment
2.3. Number of Heterotrophic Bacteria and Vibrio Count
2.4. DNA Isolation and PCR Amplification of Partial 16S rRNA Gene
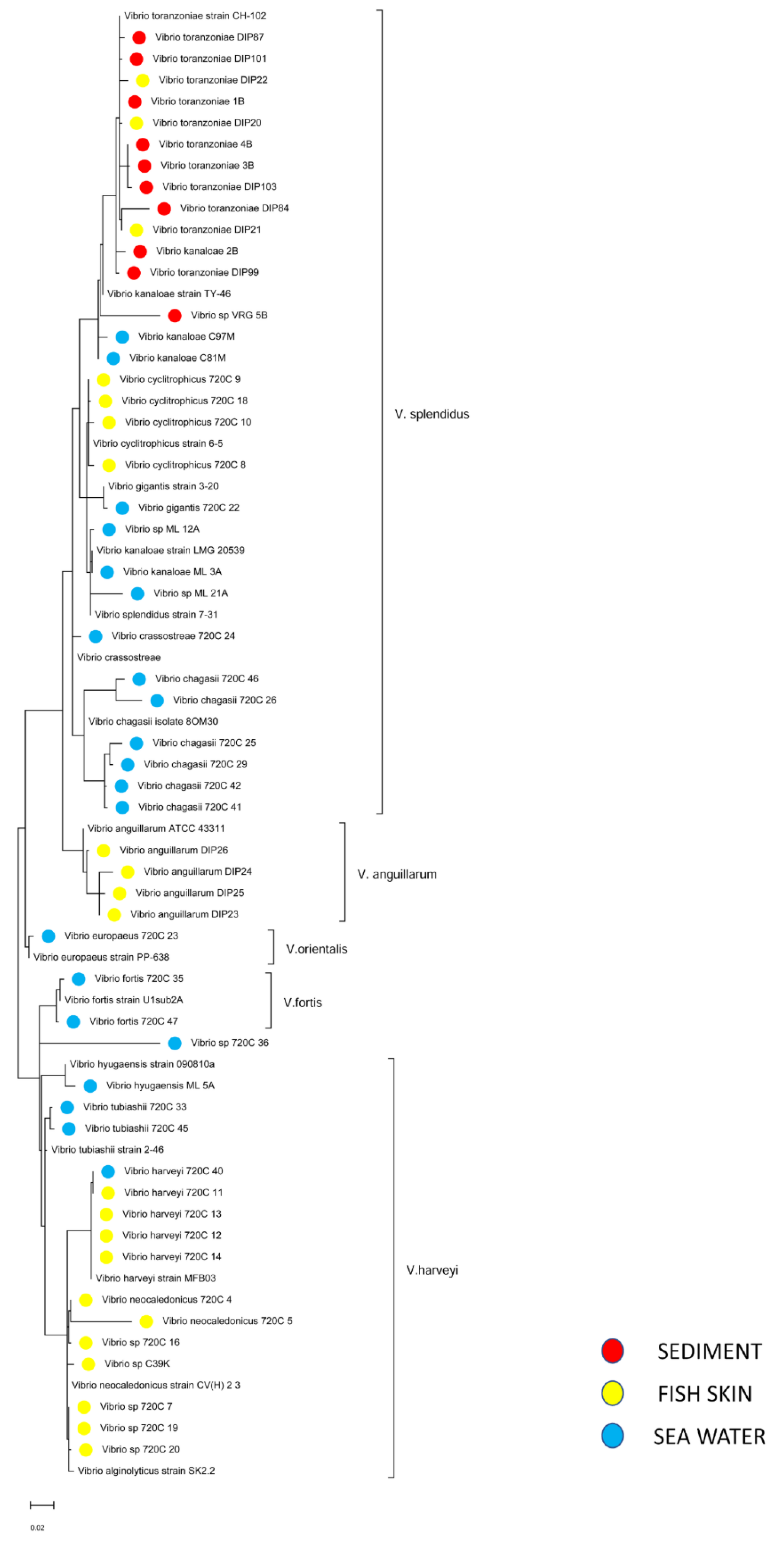
2.5. Sequencing and Phylogenetic Analysis of Vibrio sp.
2.6. Antimicrobial Susceptibility
2.7. Statistical Analysis
3. Results
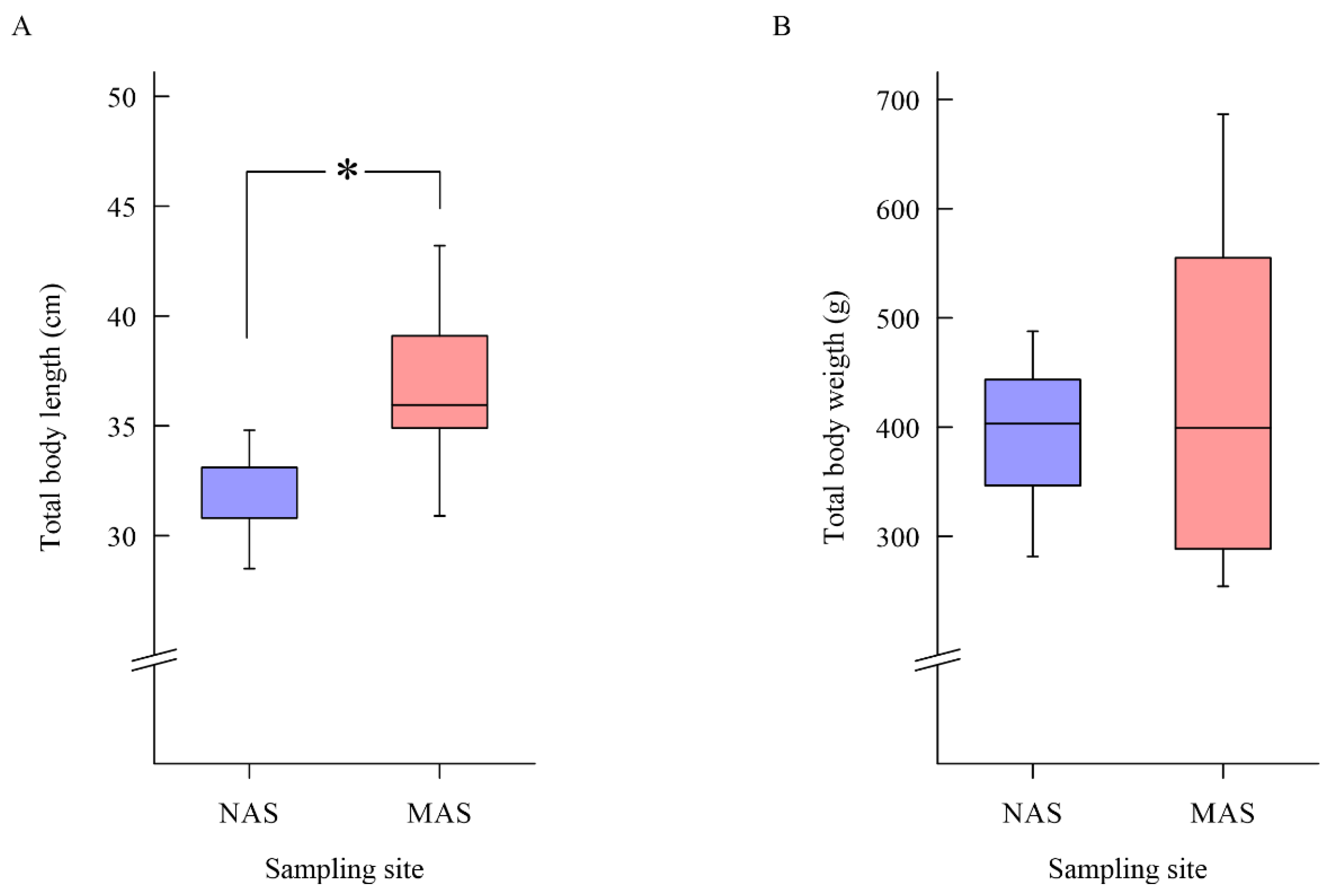
3.1. Results of Sea Bass Health Examination
3.2. Physicochemical and Microbiological Analysis of Seawater
3.3. Number of Heterotrophic Bacteria and Vibrio Count
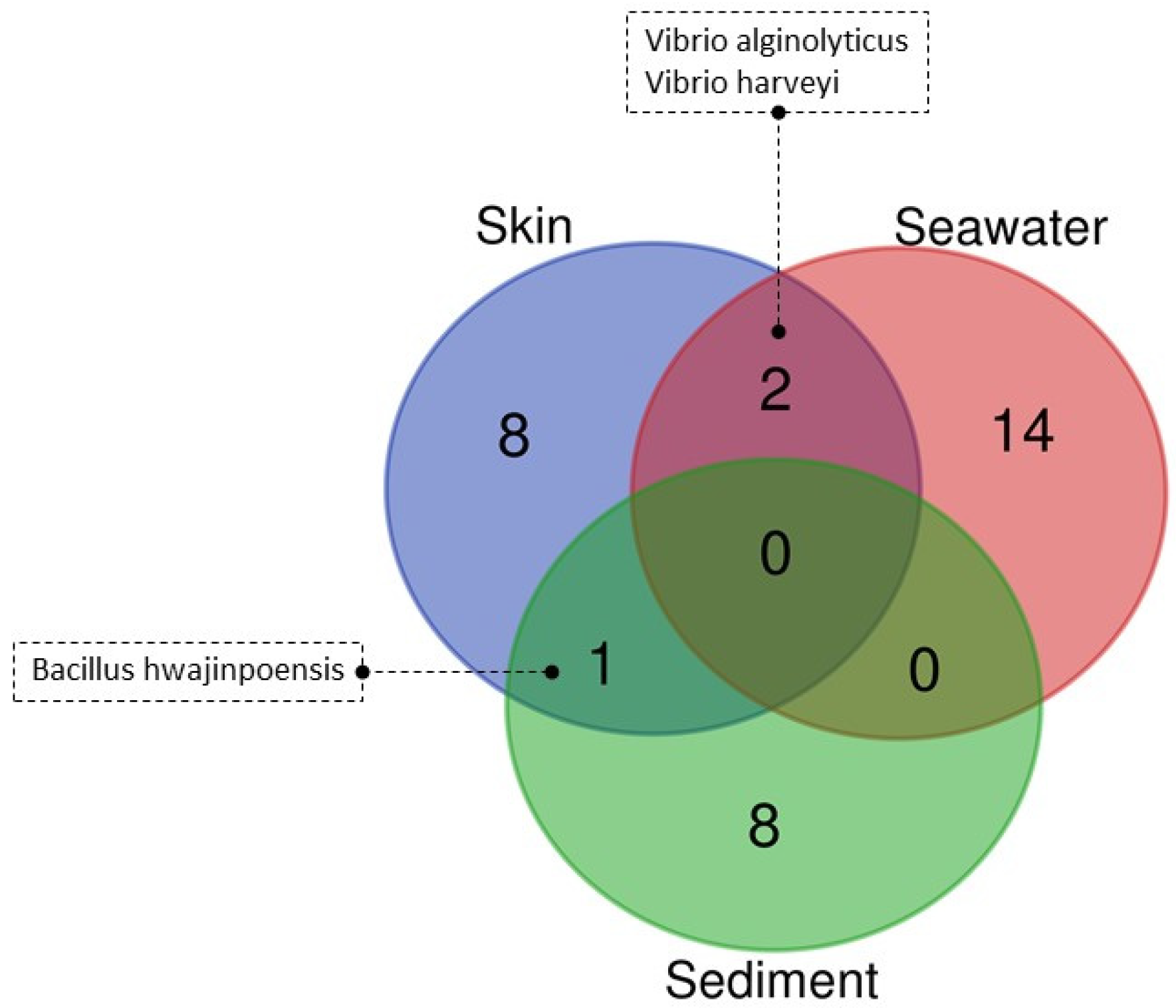
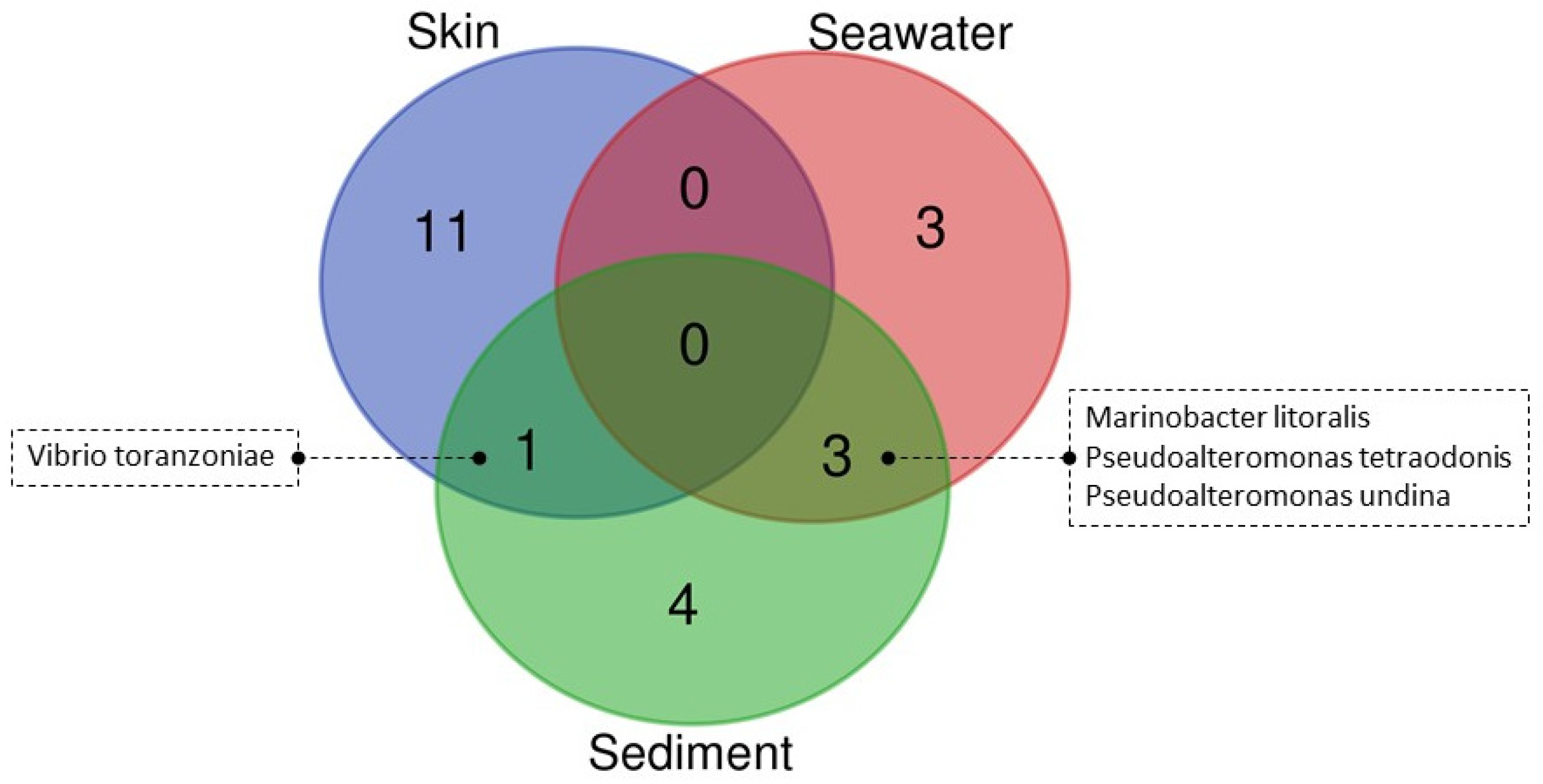
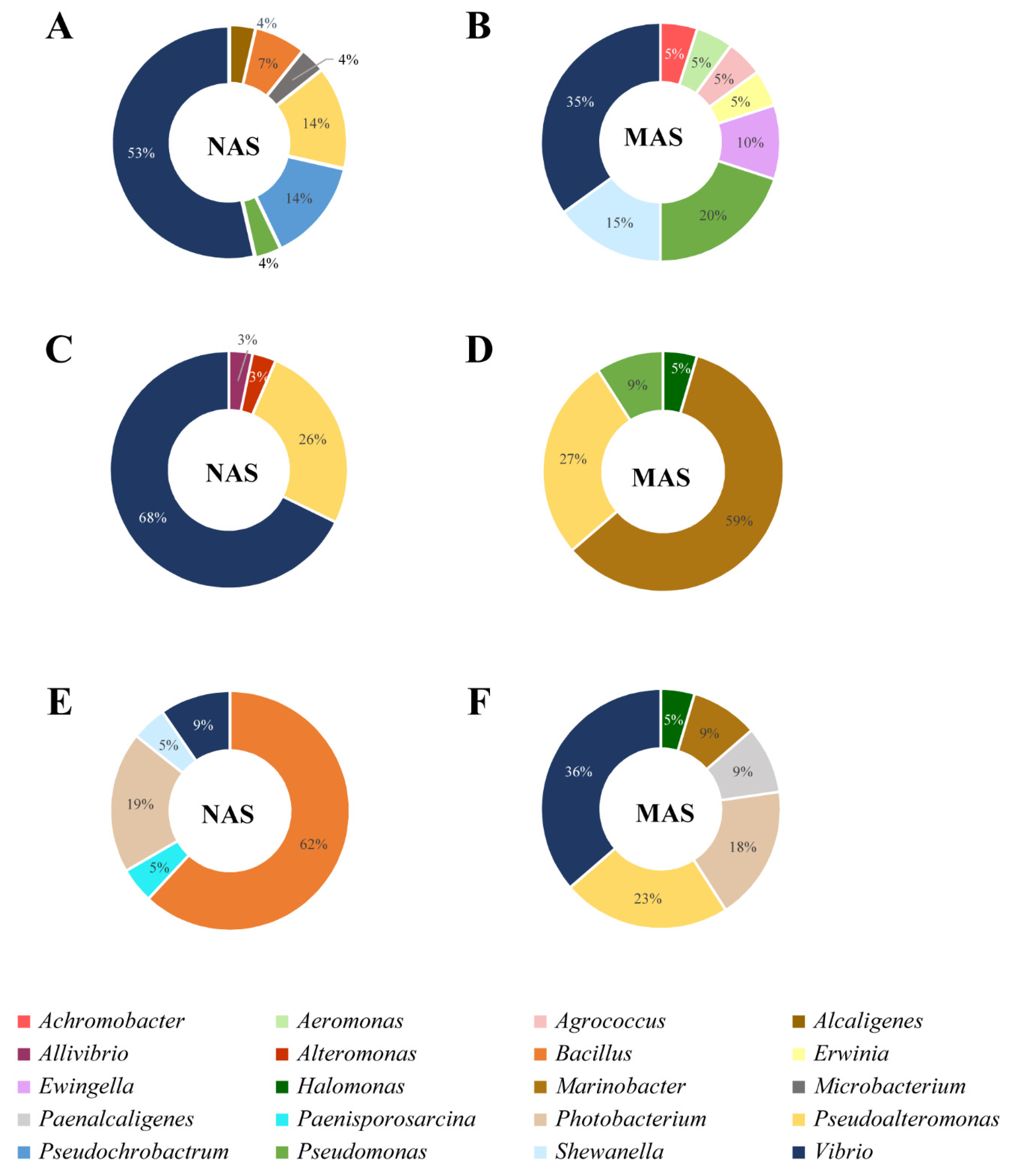
3.4. Culturable Microbiota
3.5. Antimicrobial Resistance of Bacterial Isolates
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Esteban, M.A. An Overview of the Immunological Defenses in Fish Skin. Int. Sch. Res. Netw. 2012, 2012, 853470. [Google Scholar] [CrossRef] [Green Version]
- FAO. 2021. Available online: https://www.fao.org/fishery/countrysector/naso_croatia/en (accessed on 8 November 2021).
- Camara-Ruiz, M.; Cerezo, I.M.; Guardiola, F.A.; Garcia-Beltran, J.M.; Balebona, M.C.; Morinigo, M.A.; Esteban, M.A. Alteration of the Immune Response and the Microbiota of the Skin during a Natural Infection by Vibrio harveyi in European Seabass (Dicentrarchus labrax). Microorganisms 2021, 9, 964. [Google Scholar] [CrossRef] [PubMed]
- Haenen, O.L.M.; Fouz, B.; Amaro, C.; Isern, M.M.; Mikkelsen, H.; Zrncic, S.; Travers, M.A.; Renault, T.; Wardle, R.; Hellström, A.; et al. Vibriosis in aquaculture. In Proceedings of the 16th EAFP Conference, Tampere, Finland, 4 September 2013; pp. 138–148. [Google Scholar]
- Veić, T. Characterization of Vibrio Species Isolated from European Sea Bass (Dicentrarhus Labrax, Linnaeus, 1758) Farmed on the Eastern Adriatic Sea. Master’s Thesis, University of Zagreb, Zagreb, Croatia, 2016. [Google Scholar]
- Rosado, D.; Xavier, R.; Severino, R.; Tavares, F.; Cable, J.; Perez-Losada, M. Effects of disease, antibiotic treatment and recovery trajectory on the microbiome of farmed seabass (Dicentrarchus labrax). Sci. Rep. 2019, 9, 18946. [Google Scholar] [CrossRef] [PubMed]
- Rosado, D.; Perez-Losada, M.; Severino, R.; Cable, J.; Xavier, R. Characterization of the skin and gill microbiomes of the farmed seabass (Dicentrarchus labrax) and seabream (Sparus aurata). Aquaculture 2018, 500, 57–64. [Google Scholar] [CrossRef]
- Bruijn, I.D.; Liu, Y.; Wiegertjes, G.F.; Raaijmakers, J.M. Exploring fish microbial communities to mitigate emerging diseases in aquaculture. FEMS Microbiol. Ecol. 2018, 94, 1–12. [Google Scholar] [CrossRef]
- Reverter, M.; Tapissier-Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Biological and Ecological Roles of External Fish Mucus: A Review. Fishes 2018, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Gatesoupe, F.-J.; Huelvan, C.; Le Bayon, N.; Le Delliou, H.; Madec, L.; Mouchel, O.; Quazuguel, P.; Mazurais, D.; Zambonino-Infante, J.-L. The highly variable microbiota associated to intestinal mucosa correlates with growth and hypoxia resistance of sea bass, Dicentrarchus labrax, submitted to different nutritional histories. BMC Microbiol. 2016, 16, 266. [Google Scholar] [CrossRef] [Green Version]
- Perez-Pascual, D.; Estelle, J.; Dutto, G.; Rodde, C.; Bernardet, J.-F.; Marchand, Y.; Duchaud, E.; Przybyla, C.; Ghigo, J.-M. Growth Performance and adaptability of European sea bass (Dicentrarchus labrax) gut microbiota to alternative diets free of fish products. Microorganisms 2020, 8, 1346. [Google Scholar] [CrossRef]
- Silvi, S.; Nardi, M.; Sulpizio, R.; Orpianesi, C.; Caggiano, M.; Carnevali, O.; Cresci, A. Effect of the addition of Lactobacillus delbrueckii subsp. delbrueckii on the gut microbiota composition and contribution to the well-being of European sea bass (Dicentrarchus labrax, L.). Microb. Ecol. Health Dis. 2008, 20, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Roquigny, R.; Mougin, J.; Le Bris, C.; Bonnin-Jusserand, M.; Doyen, P.; Grard, T. Characterization of the marine aquaculture microbiome: A seasonal survey in a seabass farm. Aquaculture 2021, 531, 735987. [Google Scholar] [CrossRef]
- Kapetanovic, D.; Gavrilovic, A.; Jug-Dujakovic, J.; Smrzlic, I.V.; Kazazic, S.; Bojanic-Rasovic, M.; Kolda, A.; Pesic, A.; Peric, L.; Zunic, J.; et al. Assessment of microbial sea water quality and health status of farmed European seabass (Dicentrarchus labrax) in Eastern Adriatic sea (Montenegro and Croatia). Stud. Mar. 2019, 32, 52–64. [Google Scholar] [CrossRef]
- Rosado, D.; Pérez-Losada, M.; Pereira, A.; Severino, R.; Xavier, R. Effects of aging on the skin and gill microbiota of farmed seabass and seabream. Anim. Microbiome 2021, 3, 10. [Google Scholar] [CrossRef]
- Pepi, M.; Focardi, S. Antibiotic-Resistant Bacteria in Aquaculture and Climate Change: A Challenge for Health in the Mediterranean Area. Int. J. Environ. Res. Public Health 2021, 18, 5723. [Google Scholar] [CrossRef] [PubMed]
- Damir, K.; Irena, V.S.; Damir, V.; Emin, T. Occurrence, characterization and antimicrobial susceptibility of Vibrio alginolyticus in the Eastern Adriatic sea. Mar. Pollut. Bull. 2013, 75, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.H.; Blitchington, R.B.; Greene, R.C. Amplification of bacterial 16S ribosomal DNA with polymerase chain reaction. J. Clin. Microbiol. 1990, 28, 1942–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Philipose, K.K.; Sharma, S.R.; Loka, J.; Damodaran, D.; Rao, G.S.; Vaidya, N.G.; Sonali, S.M.; Sadhu, N.; Dube, P. Observations on variations in physico-chemical water parameters of marine fish cage farm off Karwar. Indian J. Fish. 2012, 59, 83–88. [Google Scholar]
- Kapetanovic, D.; Smrzlic, I.V.; Valic, D.; Teskeredzic, Z.; Teskeredzic, E. Culturable microbiota associated with farmed Atlantic bluefin tuna (Thunnus thynnus). Aquat. Living Resour. 2017, 30, 30. [Google Scholar] [CrossRef] [Green Version]
- Matijevic, S.; Kuspilic, G.; Morovic, M.; Grbec, B.; Bogner, D.; Skejic, S.; Veza, J. Physical and chemical properties of the water column and sediments at sea bass/sea bream farm in the middle Adriatic (Maslinova bay). Acta Adriat. 2009, 50, 59–76. [Google Scholar]
- Roux, F.L.; Wegner, K.M.; Baker-Austin, C.; Vezzulli, L.; Osorio, C.R.; Amaro, C.; Ritchie, J.M.; Defoirdt, T.; Destoumieux-Garzón, D.; Blokesch, M.; et al. The emergence of Vibrio pathogens in Europe: Ecology, evolution, and pathogenesis (Paris, 11-12th March 2015). Front. Microbiol. 2015, 6, 830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatsos, I.N. Standardizing the microbiota of fish used in research. Lab. Anim. 2017, 51, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, A.; Rückert, C.; Blom, J.; Busche, T.; Reichert, J.; Schubert, P.; Goesmann, A.; Kalinowski, J.; Wilke, T.; Kämpfer, P.; et al. High diversity of Vibrio spp. associated with different ecological niches in a marine aquaria system and description of Vibrio aquimaris sp. nov. Syst. Appl. Microbiol. 2020, 43, 126123. [Google Scholar] [CrossRef] [PubMed]
- Onianwah, I.F.; Stanley, H.O.; Oyakhire, M. Microorganisms in Aquaculture Development. Glob. Adv. Res. J. Microbiol. 2018, 7, 127–131. [Google Scholar]
- Čož-Rakovac, R.; Strunjak-Perović, I.; Topić Popović, N.; Hacmenjak, M.; Šimpraga, B.; Teskeredžić, E. Health status of wild and cultured sea bass in the northern Adriatic Sea. Vet. Med. 2002, 47, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Romero, J.; Feijoo, C.G.; Navarrete, P. Antibiotics in Aquaculture—Use, Abuse and Alternatives. In Health and Environment in Aquaculture; Carvalho, E., Ed.; InTech: London, UK, 2012; ISBN 978-953-51-0497-1. Available online: http://www.intechopen.com/books/health-and-environment-in-aquaculture/antibioticsin-aquaculture-use-abuse-and-alternatives-23/12/2021 (accessed on 23 December 2021).
- Pujalte, M.J.; Sitjà-Bobadilla, A.; Macián, M.C.; Álvarez-Pellitero, P.; Garay, E. Occurrence and virulence of Pseudoalteromonas spp. in cultured gilthead sea bream (Sparus aurata L.) and European sea bass (Dicentrarchus labrax L.). Molecular and phenotypic characterisation of P. undina strain U58. Aquaculture 2007, 271, 47–53. [Google Scholar] [CrossRef]
- Hamza, F.; Kumar, A.R.; Zinjarde, S. Efficacy of cell free supernatant from Bacillus licheniformis in protecting Artemia salina against Vibrio alginolyticus and Pseudomonas gessardii. Microb. Pathog. 2018, 116, 335–344. [Google Scholar] [CrossRef]
- Vendramin, N.; Zrncic, S.; Padros, F.; Oraic, D.; Le Breton, A.; Zarza, C.; Olesen, N.J. Fish health in Mediterranean Aquaculture, past mistakes and future challenges. Bull. Eur. Assoc. Fish Pathol. 2016, 36, 38–45. [Google Scholar]
- Lasa, A.; Avendaño-Herrera, R.; Estrada, J.M.; Romalde, J.L. Isolation and identification of Vibrio toranzoniae associated with diseased red conger eel (Genypterus chilensis) farmed in Chile. Vet. Microbiol. 2015, 179, 327–331. [Google Scholar] [CrossRef]
- Zorrilla, I.; Chabrillón, M.; Arijo, S.; Dıaz-Rosales, P.; Martınez-Manzanares, E.; Balebona, M.C.; Morinigo, M.A. Bacteria recovered from diseased cultured gilthead sea bream (Sparus aurata L.) in southwestern Spain. Aquaculture 2003, 218, 11–20. [Google Scholar] [CrossRef]
- Kang, C.H.; Kim, Y.G.; Oh, S.J.; Mok, J.S.; Cho, M.H.; So, J.S. Antibiotic resistance of Vibrio harveyi isolated from seawater in Korea. Mar. Pollut. Bull. 2014, 86, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Parin, U.; Erbaş, G.; Savasan, S.; Yuksel, H.T.; Gurpinar, S.; Kirkan, S. Antimicrobial resistance of Vibrio (Listonella) anguillarum isolated from rainbow trouts (Oncorhynchus mykiss). Indian J. Anim. Res. 2019, 53, 1522–1525. [Google Scholar] [CrossRef]
- Kummerer, K. Resistance in the environment. J. Antimicrob. Chemother. 2004, 54, 311–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelossi, E.; Vezzulli, L.; Milano, A.; Branzoni, M.; Fabiano, M.; Riccardi, G.; Banat, I.M. Antibiotic resistance of benthic bacteria in fish-farm and control sediments of the Western Mediterranean. Aquaculture 2003, 219, 83–97. [Google Scholar] [CrossRef]
- Baquero, F.; Martinez, J.-L.; Canton, R. Antibiotics and antibiotic resistance in water environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Kostelic, A.; Artukovic, B.; Beck, R.; Benic, M.; Cergolj, M.; Stokovic, I.; Barac, Z. Diseases of sheep on Croatian islands. In Proceedings of the XVI. Congress of the Mediterranean Federation for Health and Production of Ruminants: FeMeSPRum, Zadar, Croatia, 26 April 2008; pp. 227–232. [Google Scholar]
- Official Gazette 21/2011 Ordinance on Pharmacologically Active Substances and Their Classification in Relation to the Maximum Permitted Levels of Residues in Food of Animal Origin. Available online: https://narodne-novine.nn.hr/clanci/sluzbeni/2005_03_29_510.html (accessed on 23 December 2021).
- Done, H.Y.; Venkatesan, A.K.; Halden, R.U. Does the Recent Growth of Aquaculture Create Antibiotic Resistance Threats Different from those Associated with Land Animal Production in Agriculture? AAPS J. 2015, 17, 513–524, Erratum in: AAPS J. 2016, 18, 1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth (m) | Secchi (m) | Sal. (ppt) | Cond. (µS/cm) | TDS (mg/L) | Temp. (°C) | pH | DO (mg/L) | DO (%) |
|---|---|---|---|---|---|---|---|---|
| NAS | 28 m | |||||||
| 0.5 m | 38.25 | 51.5 | 37.3 | 22.7 | 8.35 | 7.01 | 96.4 | |
| 6 m | 38.26 | 51.5 | 37.3 | 21.7 | 8.36 | 6.94 | 95.4 | |
| 12 m | 38.27 | 51.5 | 37.3 | 19.7 | 8.37 | 6.93 | 95.3 | |
| 0.5 m above the bottom | 38.43 | 46.8 | 37.5 | 15.1 | 8.23 | 6.62 | 83.7 | |
| MAS | 20 m | |||||||
| 0.5 m | 37.3 | 56.2 | 28.1 | 23.5 | 8.12 | 8.48 | 99.2 | |
| 6 m | 37.1 | 56.0 | 28.0 | 23.0 | 8.13 | 8.07 | 93.4 | |
| 12 m | 37.2 | 56.3 | 28.0 | 22.5 | 8.13 | 8.35 | 95.5 | |
| 0.5 m above the bottom | 37.2 | 56.2 | 28.1 | 20.1 | 8.16 | 8.02 | 87.6 |
| NAS | MAS | |||||
|---|---|---|---|---|---|---|
| Sample Type | TC (MPN/100 mL) | EC (MPN/100 mL) | EN (MPN/100 mL) | TC (MPN/100 mL) | EC (MPN/100 mL) | EN (MPN/100 mL) |
| Seawater | ||||||
| 0.5 m | 487.0 | <10.0 | <10.0 | 88.0 | 25.5 | 10.0 |
| 6 m | 588.0 | <10.0 | <10.0 | 81.5 | 10.0 | < 10.0 |
| 12 m | 1034.0 | <10.0 | <10.0 | 20.0 | 10.0 | < 10.0 |
| 0.5 m above the bottom | 10.0 | <10.0 | <10.0 | 20.5 | < 10.0 | < 10.0 |
| Sediment | <10.0 | <10.0 | <10.0 | 15.0 | < 10.0 | 46.5 |
| NAS | MAS | |||||
|---|---|---|---|---|---|---|
| Skin | Water | Sediment | Skin | Water | Sediment | |
| HPC (CFU/mL) | 69.1 ± 21.7 | 278.75 ± 196.3 | 60 ± 14.1 | 36.2 ± 20.0 | 480 ± 281.9 | 120 ± 11.3 |
| Vibrio (CFU/mL) | 38.1 ± 35.2 | 14 ± 3.6 | 3 ± 0 | 5 ± 7.1 | 55 ± 51.1 | 88 ± 4.2 |
| Sample Type | Skin (n = 28) | Seawater (n = 32) | Sediment (n = 21) | ||||
|---|---|---|---|---|---|---|---|
| Species | Percent Identity % | No. | % | No. | % | No. | % |
| Alcaligenes faecalis | 97.9 | 1 | 3.6 | ||||
| Aliivibrio finisterrensis | 99.0 | 1 | 3.1 | ||||
| Alteromonas macleodii | 99.6 | 2 | 6.3 | ||||
| Bacillus aquimaris | 99.7–99.8 | 7 | 33.3 | ||||
| Bacillus horikoshii | 99.6 | 1 | 4.8 | ||||
| Bacillus hwajinpoensis | 99.4–100.0 | 2 | 7.1 | 3 | 14.3 | ||
| Bacillus idriensis | 99.8 | 1 | 4.8 | ||||
| Bacillus tianshenii | 99.7 | 1 | 4.8 | ||||
| Microbacterium oxydans | 99.6 | 1 | 3.6 | ||||
| Paenisporosarcina quisquiliarum | 99.5 | 1 | 4.8 | ||||
| Photobacterium aphoticum | 99.0–99.9 | 4 | 19.0 | ||||
| Pseudoalteromonas arabiensis | 99.2–99.5 | 3 | 10.7 | ||||
| Pseudoalteromonas hodoensis | 99.5 | 1 | 3.1 | ||||
| Pseudoalteromonas phenolica | 99.8 | 1 | 3.1 | ||||
| Pseudoalteromonas shioyasakiensis | 99.4–99.7 | 3 | 9.4 | ||||
| Pseudoalteromonas tetraodonis | 99.7–99.9 | 3 | 9.4 | ||||
| Pseudoalteromonas undina | 100.0 | 1 | 3.6 | ||||
| Pseudochrobactrum saccharolyticum | 98.9–99.5 | 4 | 14.3 | ||||
| Pseudomonas zhaodongensis | 99.4 | 1 | 3.6 | ||||
| Shewanella marinintestina | 99.9 | 1 | 4.8 | ||||
| Vibrio alginolyticus | 99.6–100.0 | 5 | 17.9 | 2 | 6.3 | ||
| Vibrio chagasii | 98.2–98.8 | 6 | 18.8 | ||||
| Vibrio crassostreae | 99.1 | 1 | 3.1 | ||||
| Vibrio cyclitrophicus | 99.4–100.0 | 4 | 14.3 | ||||
| Vibrio europaeus | 99.7 | 1 | 3.1 | ||||
| Vibrio fortis | 99.1–99.3 | 2 | 6.3 | ||||
| Vibrio gigantis | 99.6 | 1 | 3.1 | ||||
| Vibrio harveyi | 99.7–100.0 | 4 | 14.3 | 1 | 3.1 | ||
| Vibrio hyugaensis | 99.7 | 1 | 3.1 | ||||
| Vibrio kanaloae | 99.2–100.0 | 3 | 9.4 | ||||
| Vibrio neocaledonicus | 99.8 | 2 | 7.1 | ||||
| Vibrio toranzoniae | 98.5–99.9 | 2 | 9.5 | ||||
| Vibrio tubiashii | 96.1–99.3 | 3 | 9.4 | ||||
| Sample Type | Skin (n = 20) | Seawater (n = 22) | Sediment (n = 22) | ||||
|---|---|---|---|---|---|---|---|
| Species | Percent Identity % | No. | % | No. | % | No. | % |
| Achromobacter spanius | 98.7 | 1 | 5.0 | ||||
| Aeromonas molluscorum | 99.7 | 1 | 5.0 | ||||
| Agrococcus sp. | 98.9 | 1 | 5.0 | ||||
| Erwinia billingiae | 99.6 | 1 | 5.0 | ||||
| Ewingella americana | 99.3–99.4 | 2 | 10.0 | ||||
| Halomonas aquamarina | 99.5 | 1 | 4.5 | ||||
| Halomonas boliviensis | 99.8 | 1 | 4.5 | ||||
| Marinobacter litoralis | 99.4–100.0 | 13 | 59.1 | 2 | 9.1 | ||
| Paenalcaligenes suwonensis | 99.6–99.7 | 2 | 9.1 | ||||
| Photobacterium lutimaris | 98.4–98.8 | 4 | 18.2 | ||||
| Pseudoalteromonas tetraodonis | 99.9 | 2 | 9.1 | 4 | 18.2 | ||
| Pseudoalteromonas undina | 99.5–99.8 | 4 | 18.2 | 1 | 4.5 | ||
| Pseudomonas azotoformans | 99.8 | 1 | 5.0 | ||||
| Pseudomonas gessardii | 99.8 | 1 | 5.0 | ||||
| Pseudomonas kribbensis | 99.9 | 1 | 4.5 | ||||
| Pseudomonas poae | 98.8 | 1 | 5.0 | ||||
| Pseudomonas sp. DSM 28142 | 99.9 | 1 | 5.0 | ||||
| Pseudomonas zhaodongensis | 99.7 | 1 | 4.5 | ||||
| Shewanella arctica | 99.0–99.5 | 3 | 15.0 | ||||
| Vibrio anguillarum | 98.8–99.7 | 4 | 20.0 | ||||
| Vibrio kanaloae | 96.3–99.5 | 2 | 9.1 | ||||
| Vibrio toranzoniae | 99.4–100.0 | 3 | 15.0 | 6 | 27.3 | ||
| NAS | MAS | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Antibiotic | Skin Swabs (N = 28) | Seawater (N = 32) | Sediment (N = 21) | Skin Swabs (N = 20) | Seawater (N = 22) | Sediment (N = 22) | ||||||
| No. of Isolates | % | No. of Isolates | % | No. of Isolates | % | No. of Isolates | % | No. of Isolates | % | No. of Isolates | % | |
| Enrofloxacin | 11 | 39.3 | 10 | 31.3 | 13 | 61.9 | 10 | 50.0 | 3 | 13.6 | 2 | 9.1 |
| Florfenicol | 8 | 28.6 | 3 | 9.4 | 9 | 42.9 | 13 | 65.0 | 3 | 13.6 | 3 | 13.6 |
| Gentamicin | 4 | 14.3 | 6 | 18.8 | 4 | 19.0 | 4 | 20.0 | 1 | 4.5 | - | - |
| Ampicillin | 16 | 57.1 | 16 | 50.0 | 6 | 28.6 | 20 | 100.0 | 3 | 13.6 | 7 | 31.8 |
| Erythromycin | 11 | 39.3 | 12 | 37.5 | 9 | 42.9 | 15 | 75.0 | 3 | 13.6 | 6 | 27.3 |
| Oxytetracycline | 9 | 32.1 | 1 | 3.1 | 6 | 28.6 | 7 | 35.0 | 3 | 13.6 | 2 | 9.1 |
| Sulfamethoxazole/Trimethoprim | 6 | 21.4 | 3 | 9.4 | 3 | 14.3 | 11 | 55.0 | 6 | 27.3 | 2 | 9.1 |
| Vancomycin | 24 | 85.7 | 27 | 84.4 | 9 | 42.9 | 20 | 100.0 | 20 | 90.9 | 19 | 86.4 |
| Flumequine | 6 | 21.4 | 4 | 12.5 | 11 | 52.4 | 8 | 40.0 | 4 | 18.2 | 4 | 18.2 |
| Imipenem | 5 | 17.9 | 2 | 6.3 | 6 | 28.6 | 13 | 65.0 | 1 | 4.5 | 2 | 9.1 |
| Ciprofloxacin | 9 | 32.1 | 6 | 18.8 | 10 | 47.6 | 6 | 30.0 | 4 | 18.2 | 4 | 18.2 |
| Streptomycin | 11 | 39.3 | 13 | 40.6 | 6 | 28.6 | 11 | 55.0 | 3 | 13.6 | 2 | 9.1 |
| Chloramphenicol | 4 | 14.3 | 1 | 3.1 | 3 | 14.3 | 8 | 40.0 | 2 | 9.1 | 1 | 4.5 |
| V. alginolyticus (N = 7) | V. toranzoniae (N = 11) | V. anguillarum (N = 4) | V. chagasii (N = 6) | V. harveyi (N = 5) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Antibiotic | No. of Isolates | % | No. of Isolates | % | No. of Isolates | % | No. of Isolates | % | No. of Isolates | % |
| Enrofloxacin | 0 | 2 | 18 | 4 | 100 | 2 | 40 | 2 | 40 | |
| Florfenicol | 0 | 0 | 3 | 75 | 0 | 2 | 40 | |||
| Gentamicin | 0 | 0 | 2 | 50 | 0 | 0 | ||||
| Ampicillin | 7 | 100 | 4 | 36 | 4 | 100 | 6 | 100 | 5 | 100 |
| Erythromycin | 1 | 14 | 2 | 18 | 4 | 100 | 4 | 67 | 5 | 100 |
| Oxytetracycline | 1 | 14 | 0 | 1 | 25 | 0 | 0 | |||
| Sulfamethoxazole/Trimethoprim | 0 | 1 | 9 | 1 | 25 | 0 | 0 | |||
| Vancomycin | 7 | 100 | 8 | 73 | 4 | 100 | 6 | 100 | 5 | 100 |
| Flumequine | 0 | 3 | 27 | 3 | 75 | 1 | 17 | 1 | 20 | |
| Imipenem | 1 | 14 | 4 | 36 | 3 | 75 | 0 | 1 | 20 | |
| Ciprofloxacin | 1 | 14 | 2 | 18 | 2 | 50 | 1 | 17 | 3 | 60 |
| Streptomycin | 1 | 14 | 1 | 9 | 4 | 100 | 5 | 83 | 2 | |
| Chloramphenicol | 0 | 2 | 18 | 1 | 25 | 0 | 0 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramljak, A.; Vardić Smrzlić, I.; Kapetanović, D.; Barac, F.; Kolda, A.; Perić, L.; Balenović, I.; Klanjšček, T.; Gavrilović, A. Skin Culturable Microbiota in Farmed European Seabass (Dicentrarchuslabrax) in Two Aquacultures with and without Antibiotic Use. J. Mar. Sci. Eng. 2022, 10, 303. https://doi.org/10.3390/jmse10030303
Ramljak A, Vardić Smrzlić I, Kapetanović D, Barac F, Kolda A, Perić L, Balenović I, Klanjšček T, Gavrilović A. Skin Culturable Microbiota in Farmed European Seabass (Dicentrarchuslabrax) in Two Aquacultures with and without Antibiotic Use. Journal of Marine Science and Engineering. 2022; 10(3):303. https://doi.org/10.3390/jmse10030303
Chicago/Turabian StyleRamljak, Ana, Irena Vardić Smrzlić, Damir Kapetanović, Fran Barac, Anamarija Kolda, Lorena Perić, Ivana Balenović, Tin Klanjšček, and Ana Gavrilović. 2022. "Skin Culturable Microbiota in Farmed European Seabass (Dicentrarchuslabrax) in Two Aquacultures with and without Antibiotic Use" Journal of Marine Science and Engineering 10, no. 3: 303. https://doi.org/10.3390/jmse10030303
APA StyleRamljak, A., Vardić Smrzlić, I., Kapetanović, D., Barac, F., Kolda, A., Perić, L., Balenović, I., Klanjšček, T., & Gavrilović, A. (2022). Skin Culturable Microbiota in Farmed European Seabass (Dicentrarchuslabrax) in Two Aquacultures with and without Antibiotic Use. Journal of Marine Science and Engineering, 10(3), 303. https://doi.org/10.3390/jmse10030303