
Comparative Transcriptomics of Gonads Reveals the Molecular Mechanisms Underlying Gonadal Development in Giant Freshwater Prawns (Macrobrachium rosenbergii)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. RNA Extraction and Transcriptome Sequencing
2.3. De novo Assembly and Gene Function Annotation
2.4. Analysis of DEGs and Functional Enrichment
2.5. Quantitative Real-Time PCR Validation
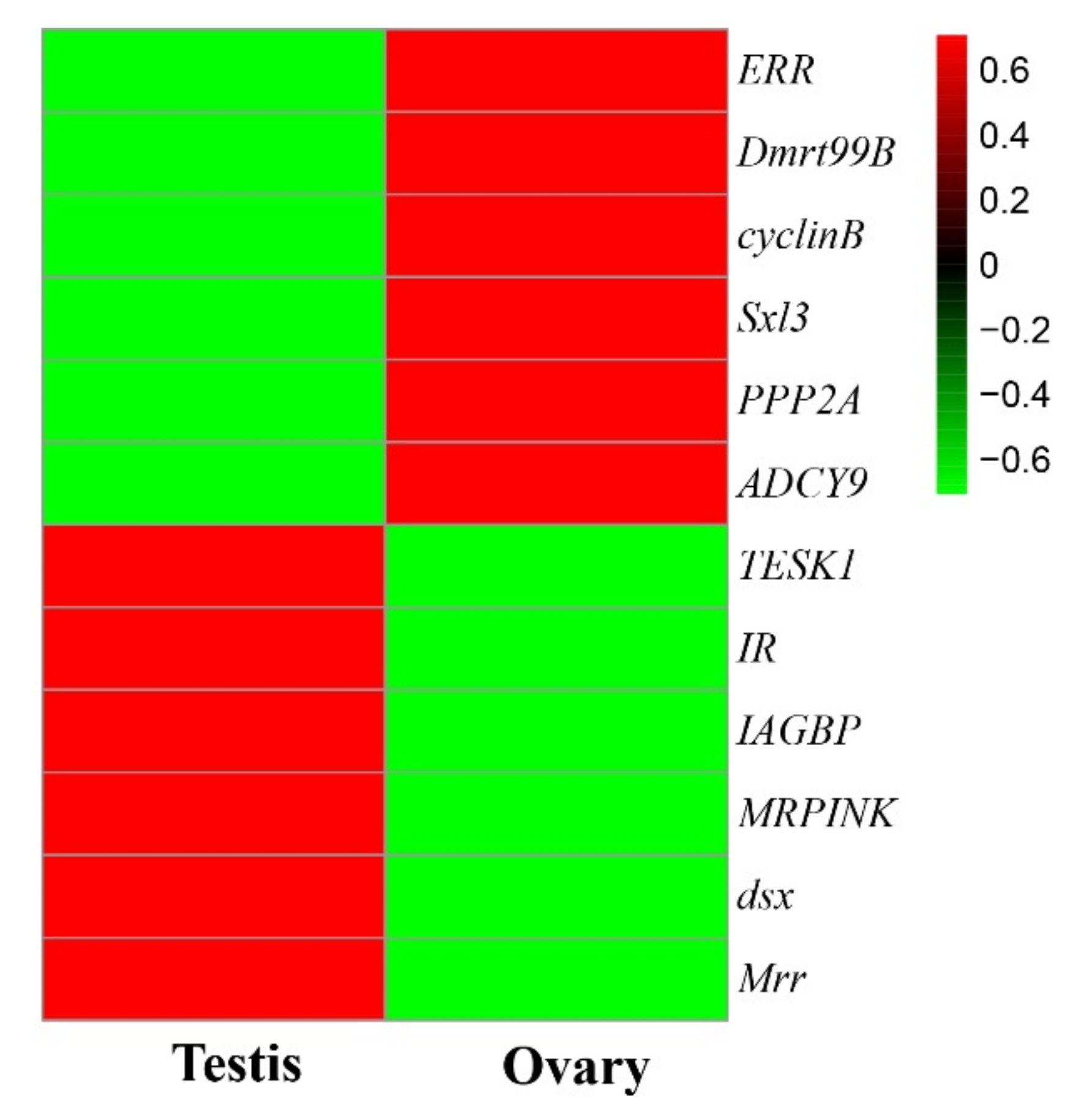
2.6. Screening of Genes Relating to Gonadal Development
2.7. Statistical Analysis
3. Results
3.1. Transcriptome Sequencing and De Novo Assembly
3.2. Functional Annotation and Classification of the Transcriptome
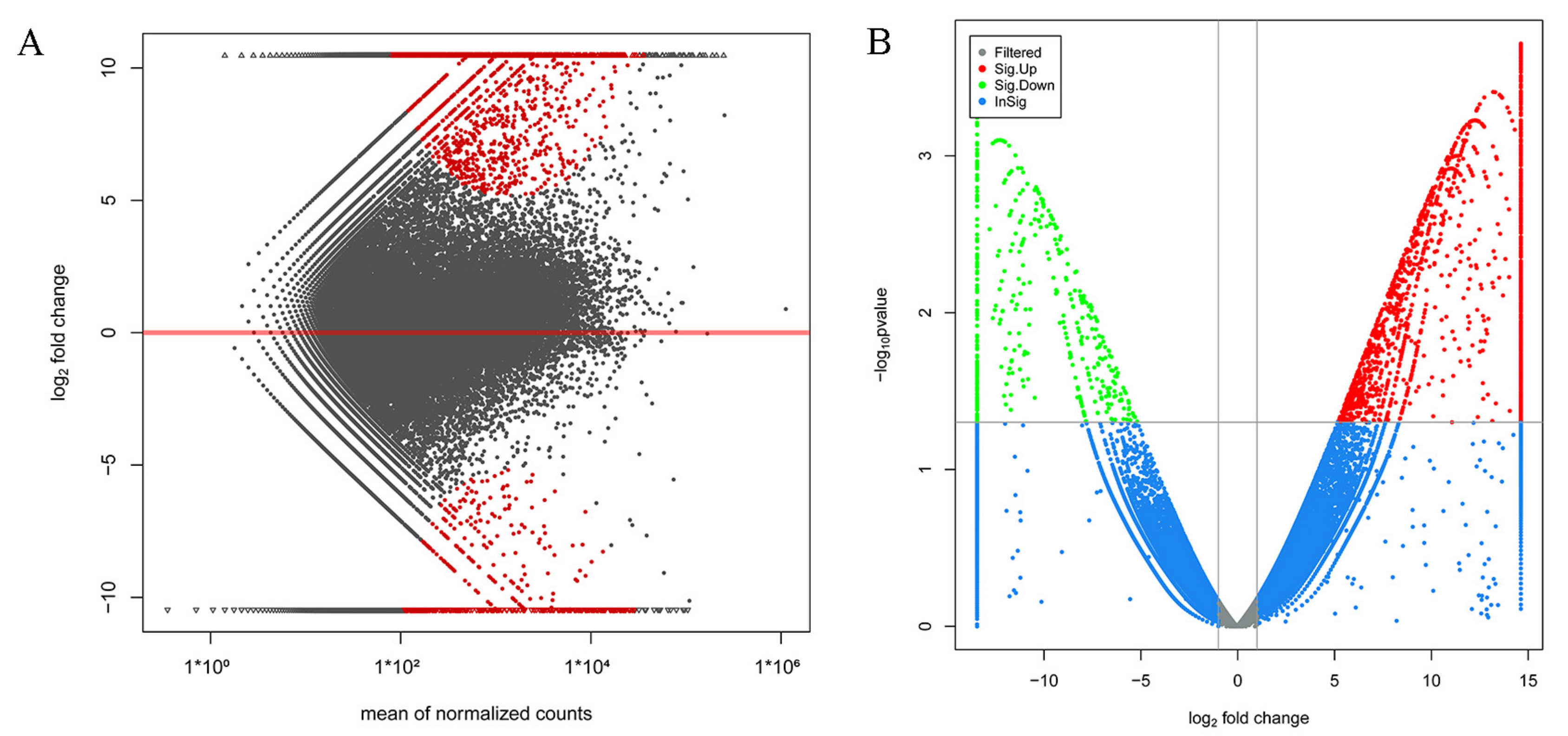
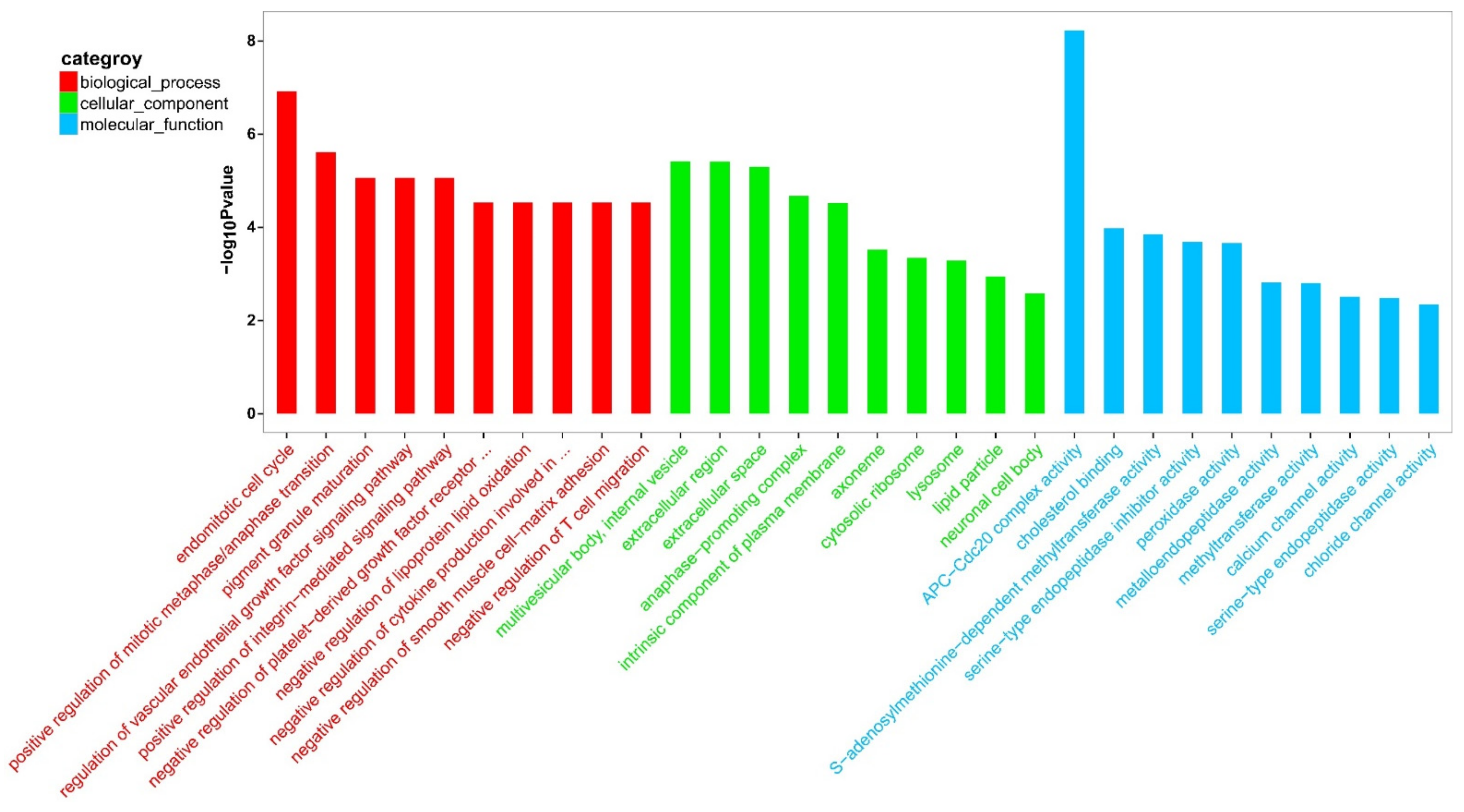
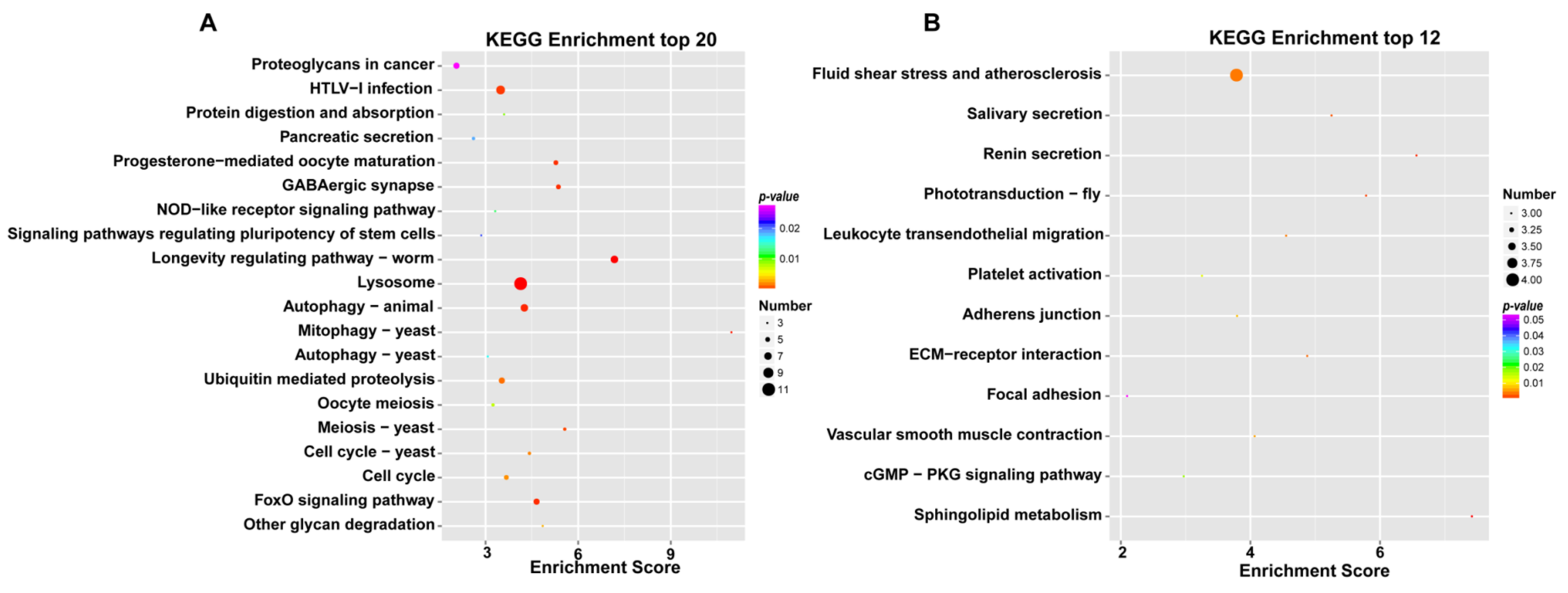
3.3. Analysis and Functional Enrichment of DEGs
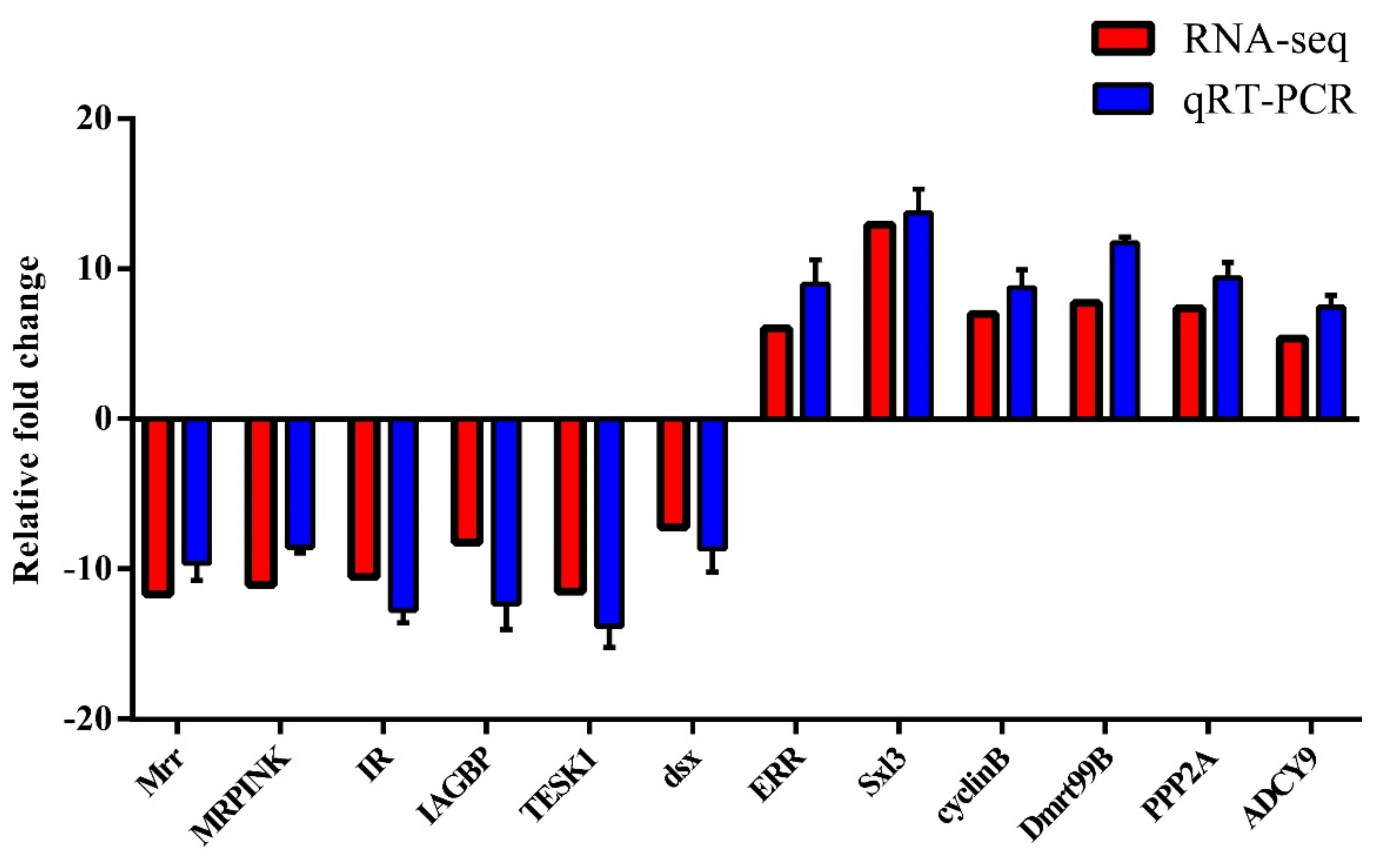
3.4. qRT-PCR Validation of DEGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- New, M. Freshwater prawn farming: Global status, recent research and a glance at the future. Aquac. Res. 2005, 36, 210–230. [Google Scholar] [CrossRef]
- Sagi, A.; Aflalo, E. The androgenic gland and monosex culture of freshwater prawn Macrobrachium rosenbergii (De Man): A biotechnological perspective. Aquac. Res. 2005, 36, 231–237. [Google Scholar] [CrossRef]
- Nair, C.M.; Salin, K.R.; Raju, M.; Sebastian, M. Economic analysis of monosex culture of giant freshwater prawn (Macrobrachium rosenbergii De Man): A case study. Aquac. Res. 2006, 37, 949–954. [Google Scholar] [CrossRef]
- Waiho, K.; Shi, X.; Fazhan, H.; Li, S.; Zhang, Y.; Zheng, H.; Liu, W.; Fang, S.; Ikhwanuddin, M.; Ma, H. High-Density Genetic Linkage Maps Provide Novel Insights Into ZW/ZZ Sex Determination System and Growth Performance in Mud Crab (Scylla paramamosain). Front. Genet. 2019, 10, 298. [Google Scholar] [CrossRef] [Green Version]
- Benzie, J.; Kenway, M.; Ballment, E. Growth of Penaeus monodon X Penaeus esculentus tiger prawn hybrids relative to the parental species. Aquaculture 2001, 193, 227–237. [Google Scholar] [CrossRef]
- Coman, F.; Sellars, M.; Norris, B.; Coman, G.; Preston, N. The effects of triploidy on Penaeus (Marsupenaeus) japonicus (Bate) survival, growth and gender when compared to diploid siblings. Aquaculture 2008, 276, 50–59. [Google Scholar] [CrossRef]
- Ma, K.Y.; Yu, S.H.; Du, Y.X.; Feng, S.Q.; Qiu, L.J.; Ke, D.Y.; Luo, M.Z.; Qiu, G.F. Construction of a Genomic Bacterial Artificial Chromosome (BAC) Library for the Prawn Macrobrachium rosenbergii and Initial Analysis of ZW Chromosome-Derived BAC Inserts. Mar. Biotechnol. 2019, 21, 206–216. [Google Scholar] [CrossRef]
- Aflalo, E.; Hoang, T.T.T.; Nguyen, V.H.; Lam, Q.; Nguyen, D.M.; Trinh, Q.S.; Raviv, S.; Sagi, A. A novel two-step procedure for mass production of all-male populations of the giant freshwater prawn Macrobrachium rosenbergii. Aquaculture 2006, 256, 468–478. [Google Scholar] [CrossRef]
- Phoungpetchara, I.; Tinikul, Y.; Poljaroen, J.; Changklungmoa, N.; Siangcham, T.; Sroyraya, M.; Chotwiwatthanakun, C.; Vanichviriyakit, R.; Hanna, P.J.; Sobhon, P. Expression of the male reproduction-related gene (Mar-Mrr) in the spermatic duct of the giant freshwater prawn, Macrobrachium rosenbergii. Cell Tissue Res. 2012, 348, 609–623. [Google Scholar] [CrossRef]
- Yang, G.; Lu, Z.; Qin, Z.; Zhao, L.; Pan, G.; Shen, H.; Zhang, M.; Liang, R.; Lin, L.; Zhang, K. Insight into the Regulatory Relationships between the Insulin-Like Androgenic Gland Hormone Gene and the Insulin-Like Androgenic Gland Hormone-binding Protein Gene in Giant Freshwater Prawns (Macrobrachium rosenbergii). Int. J. Mol. Sci. 2020, 21, 4207. [Google Scholar] [CrossRef]
- Cao, J.X.; Yin, G.L.; Yang, W.J. Identification of a novel male reproduction-related gene and its regulated expression patterns in the prawn, Macrobrachium rosenbergii. Peptides 2006, 27, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Sharabi, O.; Manor, R.; Weil, S.; Aflalo, E.D.; Lezer, Y.; Levy, T.; Aizen, J.; Ventura, T.; Mather, P.B.; Khalaila, I.; et al. Identification and Characterization of an Insulin-Like Receptor Involved in Crustacean Reproduction. Endocrinology 2016, 157, 928–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Ma, W.M.; Dai, J.Q.; Feng, C.Z.; Yang, F.; Ohira, T.; Nagasawa, H.; Yang, W.J. Inhibition of a novel sperm gelatinase in prawn sperm by the male reproduction-related Kazal-type peptidase inhibitor. Mol. Reprod. Dev. 2008, 75, 1327–1337. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Deng, S.P.; Jiang, D.N.; Chen, H.P.; Guang-Li, L.I.; Tian-Li, W.U.; Tian, C.X.; Zhu, C.H.; University, G.O. Screening of Ovarian Genes Associated with Reproduction in Macrobrachium rosenbergii and Their Changes in Expression Pattern in Different Development Stages after ERR Interference. J. Guangdong Ocean Univ. 2018, 3, 8–16. [Google Scholar] [CrossRef]
- Zhao, M.X.; Chen, H.P.; Liu, J.L.; Deng, S.P.; Guang-Li, L.I.; Zhu, C.H.; Hong, Y.C. Prokaryotic Expression and Purification of Estrogen Related Receptor(ERR) Gene from Macrobrachium rosenbergii. J. Guangdong Ocean Univ. 2017, 1, 108–112. [Google Scholar] [CrossRef]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Raviv, S.; Glazer, L.; Sagi, A. Temporal silencing of an androgenic gland-specific insulin-like gene affecting phenotypical gender differences and spermatogenesis. Endocrinology 2009, 150, 1278–1286. [Google Scholar] [CrossRef] [Green Version]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Rosen, O.; Sagi, A. Timing sexual differentiation: Full functional sex reversal achieved through silencing of a single insulin-like gene in the prawn, Macrobrachium rosenbergii. Biol. Reprod. 2012, 86, 90. [Google Scholar] [CrossRef]
- Ventura, T.; Rosen, O.; Sagi, A. From the discovery of the crustacean androgenic gland to the insulin-like hormone in six decades. Gen. Comp. Endocrinol. 2011, 173, 381–388. [Google Scholar] [CrossRef]
- Meeratana, P.; Sobhon, P. Classification of differentiating oocytes during ovarian cycle in the giant freshwater prawn, Macrobrachium rosenbergii De Man. Aquaculture 2007, 270, 249–258. [Google Scholar] [CrossRef]
- Chen, J.; Liu, P.; Li, Z.; Chen, Y.; Qiu, G.-F. The cloning of the cdk2 transcript and the localization of its expression during gametogenesis in the freshwater giant prawn, Macrobrachium rosenbergii. Mol. Biol. Rep. 2013, 40, 4781–4790. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Deng, Y.Y.; Li, J.Q.; Wu, S.F.; Zhu, Y.; Chen, Y.W.; He, F.C. Integrated nr Database in Protein Annotation System and Its Localization. Comput. Eng. 2006, 32, 71–72. [Google Scholar]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The Universal Protein knowledgebase. Nucleic Acids Res. 2004, 32, D115–D119. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Krylov, D.M.; Makarova, K.S.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; Rao, B.S.; et al. A comprehensive evolutionary classification of proteins encoded in complete eukaryotic genomes. Genome Biol. 2004, 5, R7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Kobayashi, K.; Watanabe, H.; Iguchi, T. Environmental sex determination in the branchiopod crustacean Daphnia magna: Deep conservation of a Doublesex gene in the sex-determining pathway. PLoS Genet. 2011, 7, e1001345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toshima, J.; Nakagawara, K.; Mori, M.; Noda, T.; Mizuno, K. Structural organization and chromosomal localization of the mouse tesk1 (testis-specific protein kinase 1) gene. Gene 1998, 206, 237–245. [Google Scholar] [CrossRef]
- Nagaraju, G.P. Reproductive regulators in decapod crustaceans: An overview. J. Exp. Biol. 2011, 214, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Chen, L.L.; Li, G.L.; Deng, S.P.; Zhu, C.H. Accumulation and Depuration of Nonylphenol and Its Effect on the Expressions of Vitellogenin and Vitellogenin Receptor in Freshwater Prawn Macrobrachium rosenbergii. Bull. Environ. Contam. Toxicol. 2019, 103, 729–733. [Google Scholar] [CrossRef]
- Jiang, J.; Yuan, X.; Qiu, Q.; Huang, G.; Jiang, Q.; Fu, P.; Zhang, Y.; Jia, Y.; Yang, X.; Jiang, H. Comparative Transcriptome Analysis of Gonads for the Identification of Sex-Related Genes in Giant Freshwater Prawns (Macrobrachium rosenbergii) Using RNA Sequencing. Genes 2019, 10, 1035. [Google Scholar] [CrossRef] [Green Version]
- Peranandam, R.; Palanisamy, I.; Lourdaraj, A.V.; Natesan, M.; Vimalananthan, A.P.; Thangaiyan, S.; Perumal, A.; Muthukalingan, K. TBT effects on the development of intersex (ovotestis) in female fresh water prawn Macrobrachium rosenbergii. BioMed Res. Int. 2014, 2014, 412619. [Google Scholar] [CrossRef] [Green Version]
- Assis, R.; Zhou, Q.; Bachtrog, D. Sex-biased transcriptome evolution in Drosophila. Genome Biol. Evol. 2012, 4, 1189–1200. [Google Scholar] [CrossRef] [Green Version]
- Ellegren, H.; Parsch, J. The evolution of sex-biased genes and sex-biased gene expression. Nat. Rev. Genet. 2007, 8, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Small, C.M.; Carney, G.E.; Mo, Q.; Vannucci, M.; Jones, A.G. A microarray analysis of sex- and gonad-biased gene expression in the zebrafish: Evidence for masculinization of the transcriptome. BMC Genom. 2009, 10, 579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Sun, L.; Guan, W.; Zhou, C.; Tang, B.; Cheng, Y.; Huang, J.; Xuan, F. De novo transcriptome sequencing and analysis of male and female swimming crab (Portunus trituberculatus) reproductive systems during mating embrace (stage II). BMC Genet. 2018, 19, 3. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.X.; Dai, J.Q.; Dai, Z.M.; Yin, G.L.; Yang, W.J. A male reproduction-related Kazal-type peptidase inhibitor gene in the prawn, Macrobrachium rosenbergii: Molecular characterization and expression patterns. Mar. Biotechnol. 2007, 9, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qian, Y.Q.; Ma, W.M.; Yang, W.J. Inhibition mechanism and the effects of structure on activity of male reproduction-related peptidase inhibitor Kazal-type (MRPINK) of Macrobrachium rosenbergii. Mar. Biotechnol. 2009, 11, 252–259. [Google Scholar] [CrossRef]
- Fafalios, A.; Ma, J.; Tan, X.; Stoops, J.; Luo, J.; Defrances, M.C.; Zarnegar, R. A hepatocyte growth factor receptor (Met)-insulin receptor hybrid governs hepatic glucose metabolism. Nat. Med. 2011, 17, 1577–1584. [Google Scholar] [CrossRef]
- Nef, S.; Verma-Kurvari, S.; Merenmies, J.; Vassalli, J.D.; Efstratiadis, A.; Accili, D.; Parada, L.F. Testis determination requires insulin receptor family function in mice. Nature 2003, 426, 291–295. [Google Scholar] [CrossRef]
- Toyota, K.; Kato, Y.; Sato, M.; Sugiura, N.; Miyagawa, S.; Miyakawa, H.; Watanabe, H.; Oda, S.; Ogino, Y.; Hiruta, C.; et al. Molecular cloning of doublesex genes of four cladocera (water flea) species. BMC Genom. 2013, 14, 239. [Google Scholar] [CrossRef] [Green Version]
- Escriva, H.; Delaunay, F.; Laudet, V. Ligand binding and nuclear receptor evolution. BioEssays News Rev. Mol. Cell. Dev. Biol. 2000, 22, 717–727. [Google Scholar] [CrossRef]
- Song, Y. Cloning and Prokaryotic Expression of Bmand-Sxl Gene In Bombyx mandarina. Jiangsu Seric. 2009, 31, 14–18. [Google Scholar] [CrossRef]
- Fei, J.M.; Shi, L.L.; Li, Y.H. Cloning and expression analysis of the sex-lethal gene of Procterus cruzi. J. Huazhong Agric Univ 2021, 40, 120–128. [Google Scholar] [CrossRef]
- Kopp, A. Dmrt genes in the development and evolution of sexual dimorphism. Trends Genet. TIG 2012, 28, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Wilken, J.; Phillips, N.B.; Narendra, U.; Chan, G.; Stratton, S.M.; Kent, S.B.; Weiss, M.A. Sexual dimorphism in diverse metazoans is regulated by a novel class of intertwined zinc fingers. Genes Dev. 2000, 14, 1750–1764. [Google Scholar] [CrossRef]
- Ma, A.; Wang, Y.; Zou, Z.; Fu, M.; Lin, P.; Zhang, Z. Erk2 in ovarian development of green mud crab Scylla paramamosain. DNA Cell Biol. 2012, 31, 1233–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, B.; Zhang, Z.; Wang, Y.; Wang, G.; Chen, Y.; Lin, P.; Wang, S.; Zou, Z. Differential expression of ubiquitin-conjugating enzyme E2r in the developing ovary and testis of penaeid shrimp Marsupenaeus japonicus. Mol. Biol. Rep. 2009, 36, 1149–1157. [Google Scholar] [CrossRef]
- Archibald, J.M.; Teh, E.M.; Keeling, P.J. Novel ubiquitin fusion proteins: Ribosomal protein P1 and actin. J. Mol. Biol. 2003, 328, 771–778. [Google Scholar] [CrossRef]
- Finley, D.; Bartel, B.; Varshavsky, A. The tails of ubiquitin precursors are ribosomal proteins whose fusion to ubiquitin facilitates ribosome biogenesis. Nature 1989, 338, 394–401. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, L.; Wang, Y.; Li, W.; He, L.; Jiang, H. Expression characteristics of two ubiquitin/ribosomal fusion protein genes in the developing testis, accessory gonad and ovary of Chinese mitten crab, Eriocheir sinensis. Mol. Biol. Rep. 2012, 39, 6683–6692. [Google Scholar] [CrossRef]
- Sun, X.; He, Y.; Hou, L.; Yang, W.X. Myosin Va participates in acrosomal formation and nuclear morphogenesis during spermatogenesis of Chinese mitten crab Eriocheir sinensis. PLoS ONE 2010, 5, e12738. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Qiu, L.; Jiang, S.; Zhou, F.; Huang, J.; Yang, L.; Su, T.; Zhang, D. Molecular cloning and mRNA expression of M-phase phosphoprotein 6 gene in black tiger shrimp (Penaeus monodon). Mol. Biol. Rep. 2013, 40, 1301–1306. [Google Scholar] [CrossRef]
- Ma, W.M.; Qian, Y.Q.; Wang, M.R.; Yang, F.; Yang, W.J. A novel terminal ampullae peptide is involved in the proteolytic activity of sperm in the prawn, Macrobrachium rosenbergii. Reproduction 2010, 140, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q.; Lu, B.; Lin, D.; Huang, H.; Chen, X.; Ye, H. Role of crustacean female sex hormone (CFSH) in sex differentiation in early juvenile mud crabs, Scylla paramamosain. Gen. Comp. Endocrinol. 2020, 289, 113383. [Google Scholar] [CrossRef] [PubMed]
- Roth, Z.; Khalaila, I. Identification and characterization of the vitellogenin receptor in Macrobrachium rosenbergii and its expression during vitellogenesis. Mol. Reprod. Dev. 2012, 79, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Xiong, Y.; Jiang, S.; Zhang, W.; Xu, L.; Jin, S.; Gong, Y.; Wu, Y.; Fu, H. Three neuroparsin genes from oriental river prawn, Macrobrachium nipponense, involved in ovary maturation. 3 Biotech 2020, 10, 537. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Raw Bases (bp) | Clean Reads | Clean Bases (bp) | Valid Ratio (Base) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|
| testis | 89,598,146 | 11,199,768,250 | 84,214,494 | 10,522,214,704 | 93.95% | 92.68% | 43% |
| ovary | 78,074,612 | 9,759,326,500 | 73,971,508 | 9,243,212,064 | 94.71% | 93.58% | 41% |
| Length | All ≥300 bp | ≥500 bp | ≥1000 bp | N50 | Total Length | Max Length | Min Length | Average Length |
|---|---|---|---|---|---|---|---|---|
| Unigene | 75,887 | 42,533 | 22,622 | 2063 | 85,974,378 | 20,859 | 301 | 1132.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Qin, Z.; Lu, Z.; Liang, R.; Zhao, L.; Pan, G.; Lin, L.; Zhang, K. Comparative Transcriptomics of Gonads Reveals the Molecular Mechanisms Underlying Gonadal Development in Giant Freshwater Prawns (Macrobrachium rosenbergii). J. Mar. Sci. Eng. 2022, 10, 737. https://doi.org/10.3390/jmse10060737
Yang G, Qin Z, Lu Z, Liang R, Zhao L, Pan G, Lin L, Zhang K. Comparative Transcriptomics of Gonads Reveals the Molecular Mechanisms Underlying Gonadal Development in Giant Freshwater Prawns (Macrobrachium rosenbergii). Journal of Marine Science and Engineering. 2022; 10(6):737. https://doi.org/10.3390/jmse10060737
Chicago/Turabian StyleYang, Guang, Zhendong Qin, Zhijie Lu, Rishen Liang, Lijuan Zhao, Gan Pan, Li Lin, and Kai Zhang. 2022. "Comparative Transcriptomics of Gonads Reveals the Molecular Mechanisms Underlying Gonadal Development in Giant Freshwater Prawns (Macrobrachium rosenbergii)" Journal of Marine Science and Engineering 10, no. 6: 737. https://doi.org/10.3390/jmse10060737
APA StyleYang, G., Qin, Z., Lu, Z., Liang, R., Zhao, L., Pan, G., Lin, L., & Zhang, K. (2022). Comparative Transcriptomics of Gonads Reveals the Molecular Mechanisms Underlying Gonadal Development in Giant Freshwater Prawns (Macrobrachium rosenbergii). Journal of Marine Science and Engineering, 10(6), 737. https://doi.org/10.3390/jmse10060737