
Physicochemical Control of Caribbean Coral Calcification Linked to Host and Symbiont Responses to Varying pCO2 and Temperature

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Experimental Culturing
2.3. Net Calcification Rate, Coral Symbiont and Host Physiological Measurements
2.4. Coral Sample Preparation for Geochemical Analyses
2.5. Boron Isotopic Analysis
2.6. B/Ca Analysis
2.7. Modelled pHCF
2.8. Modelled [CO32−]CF, DICCF, and ΩCF
2.9. Carbon Stable Isotope Analyses
2.10. Statistical Methods
2.11. Data Compilation
3. Results
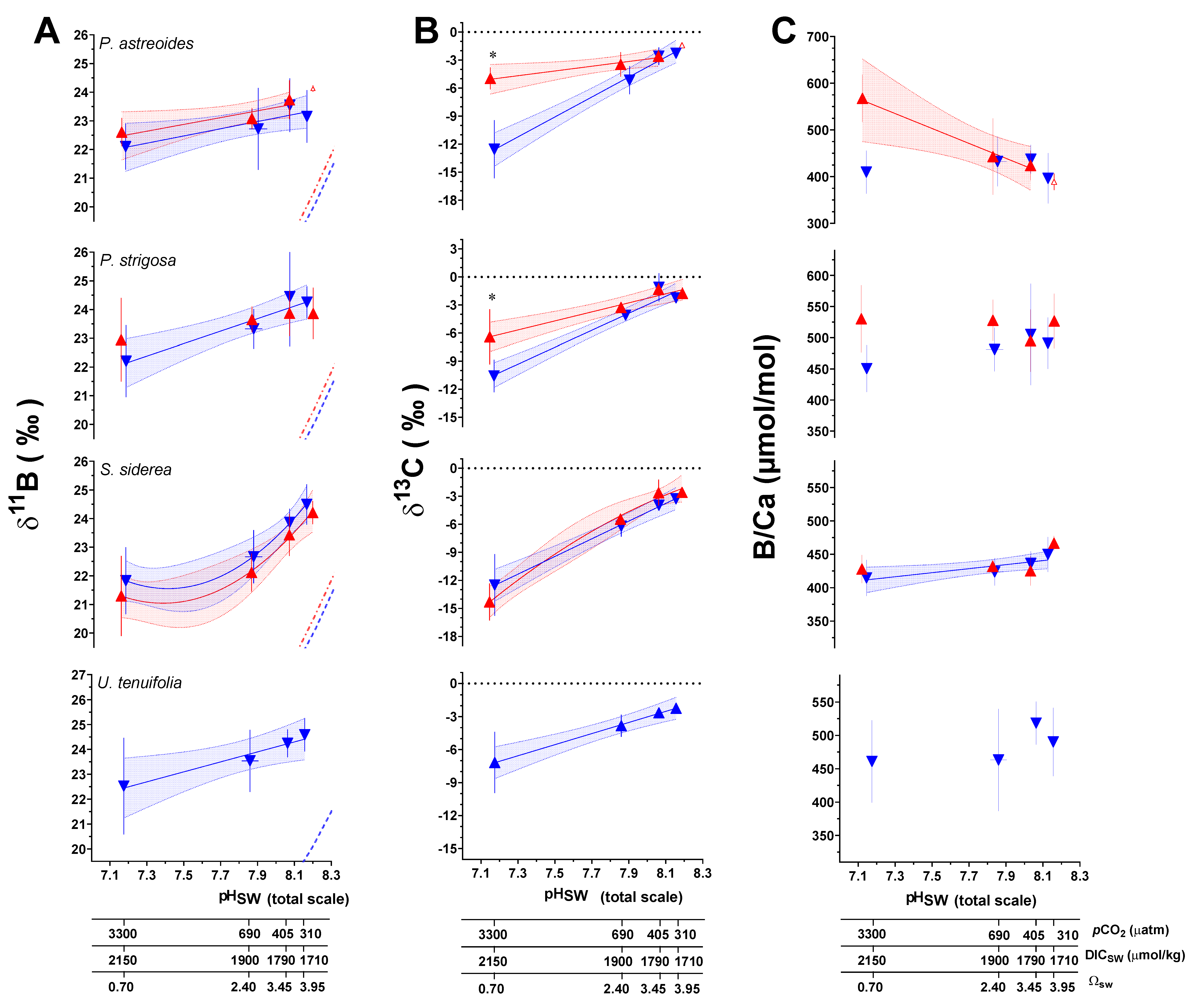
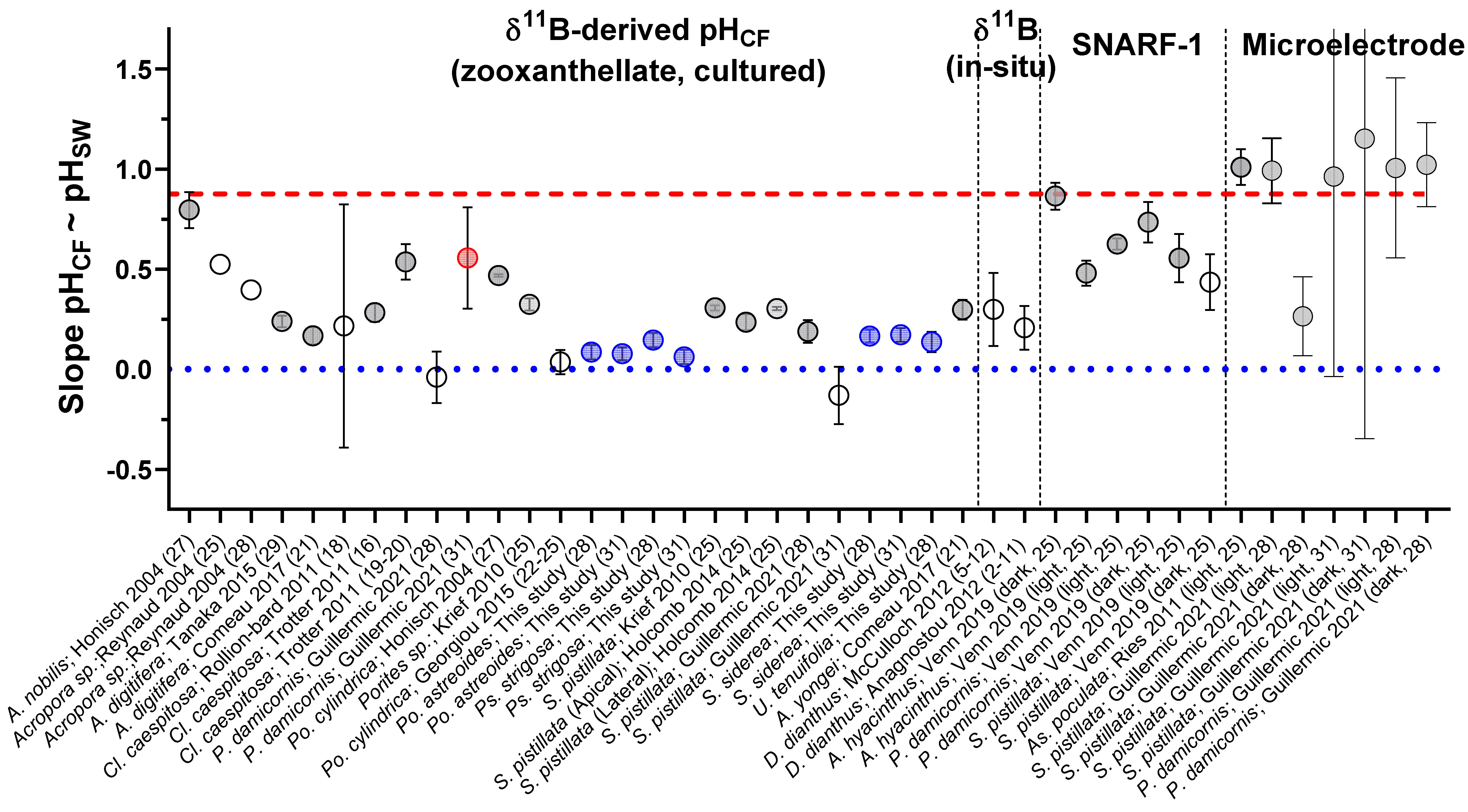
3.1. Analyses of (δ11B), B/Ca, and δ13C
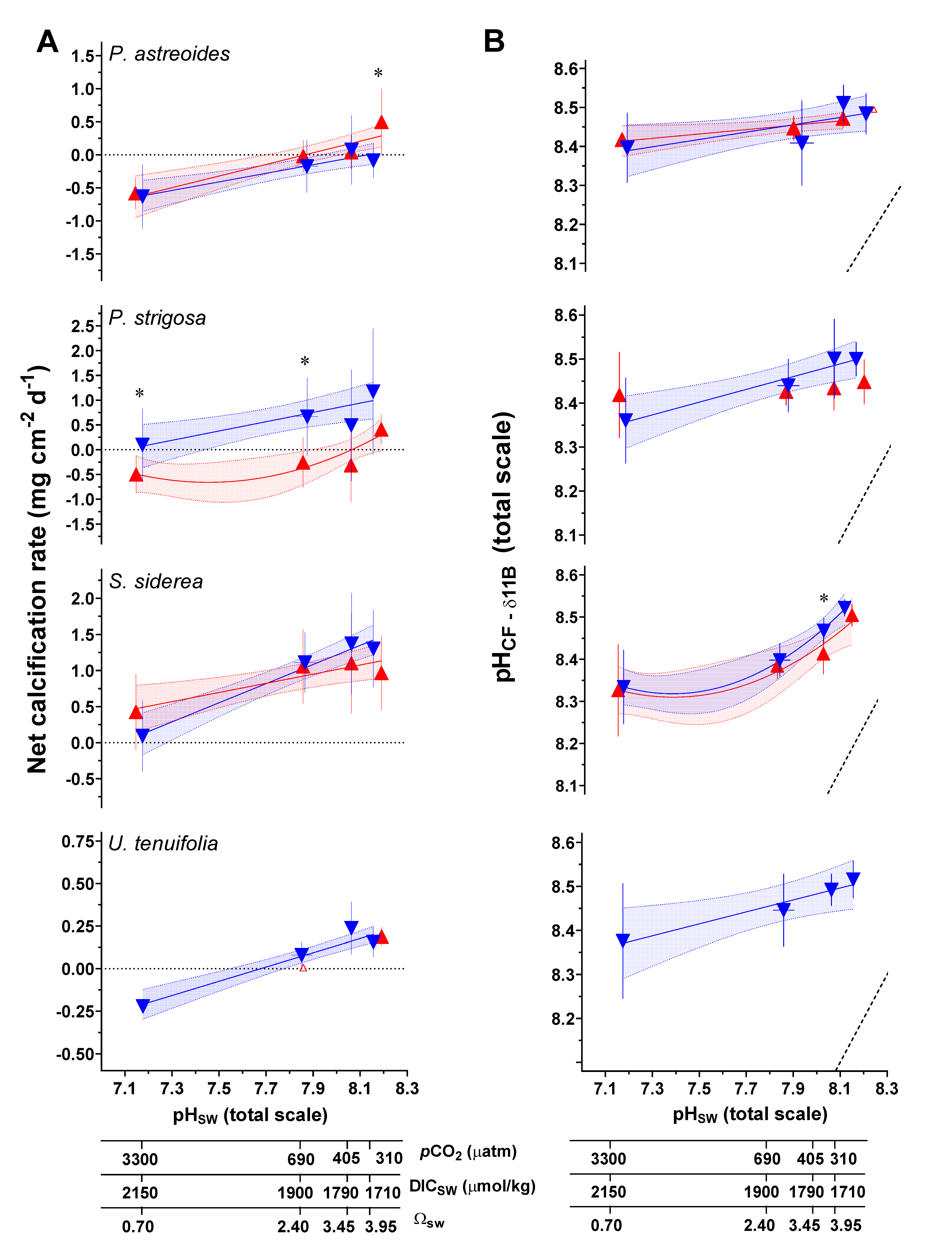
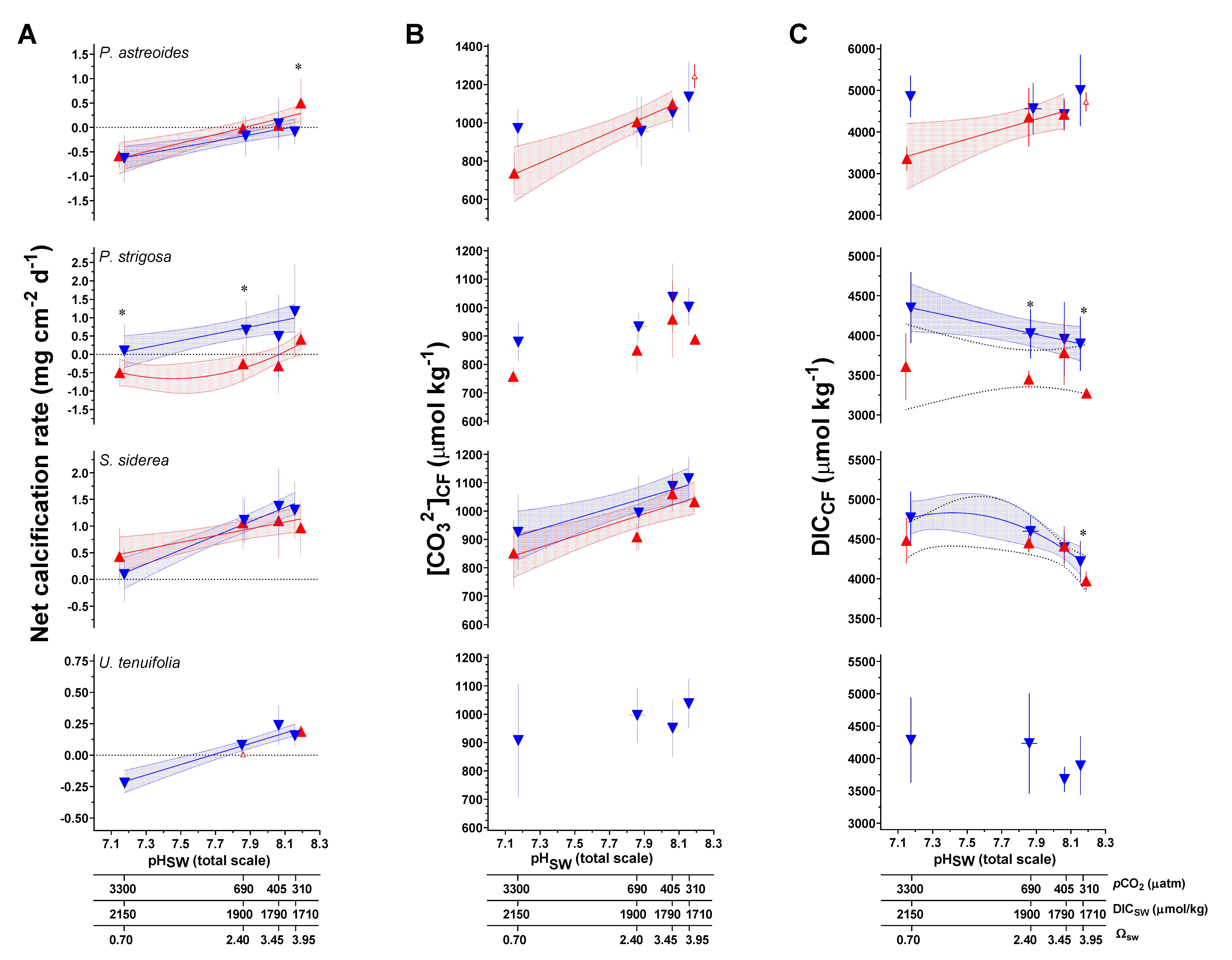
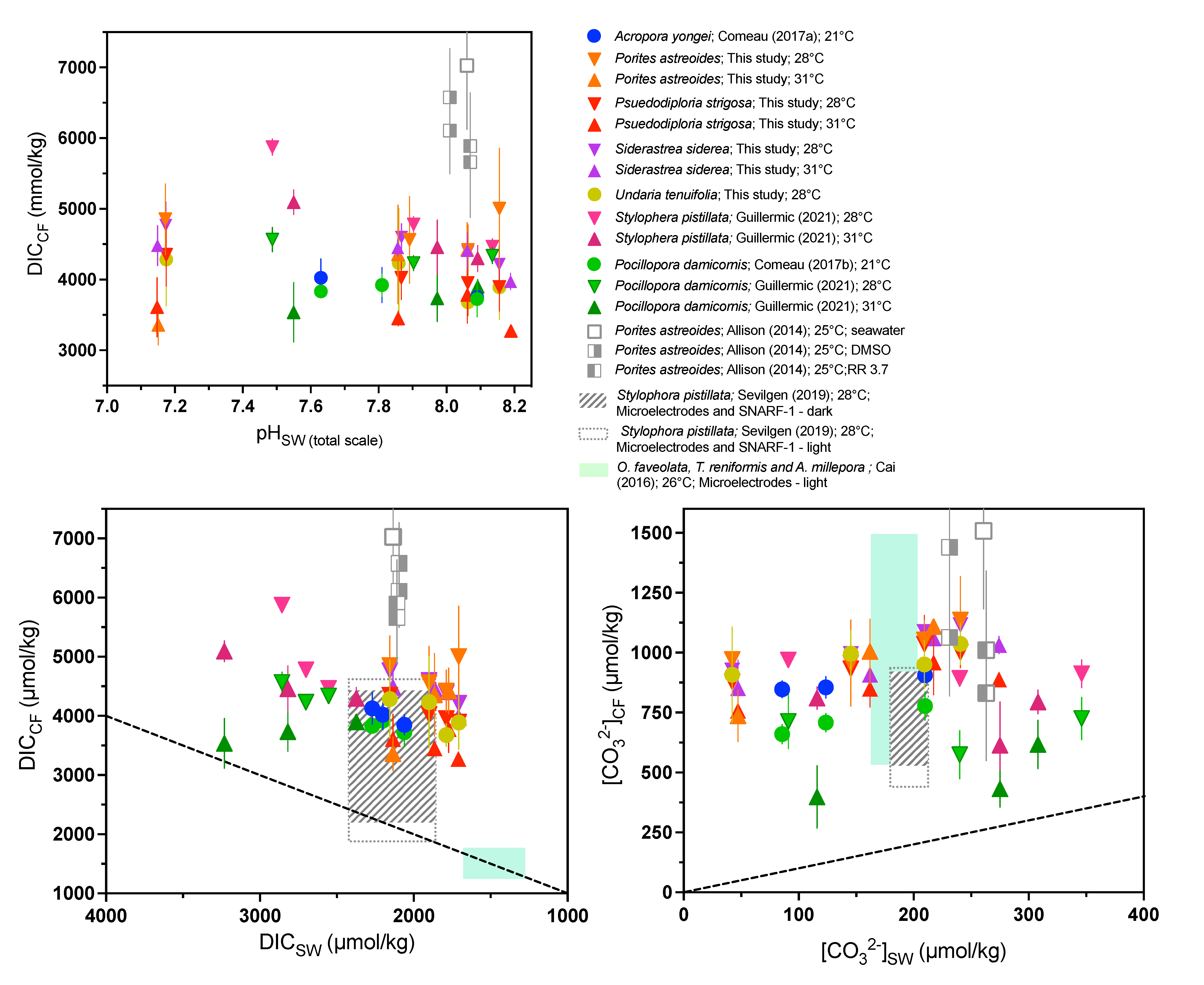
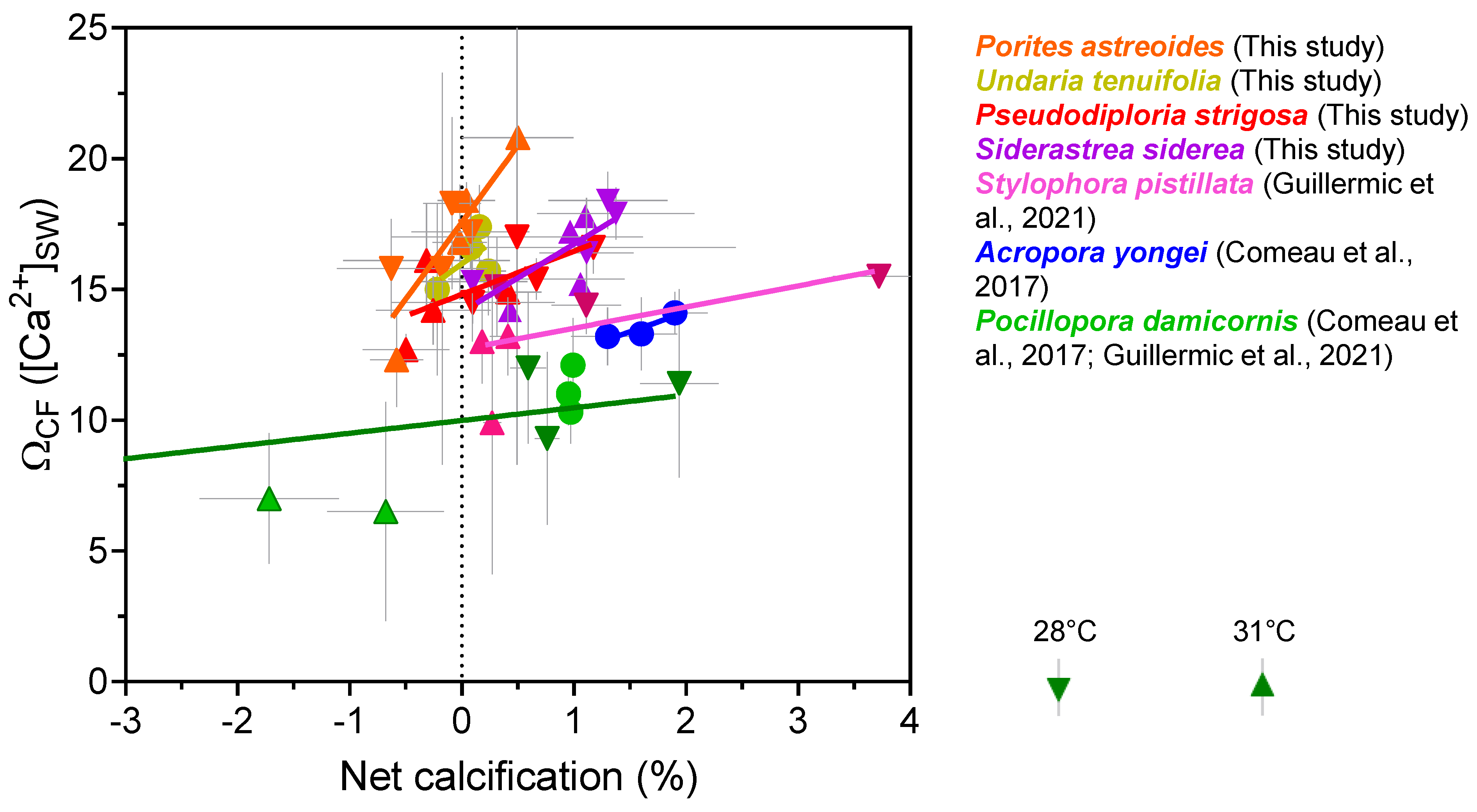
3.2. Net Calcification, pHCF, [CO32−]CF, and DICCF
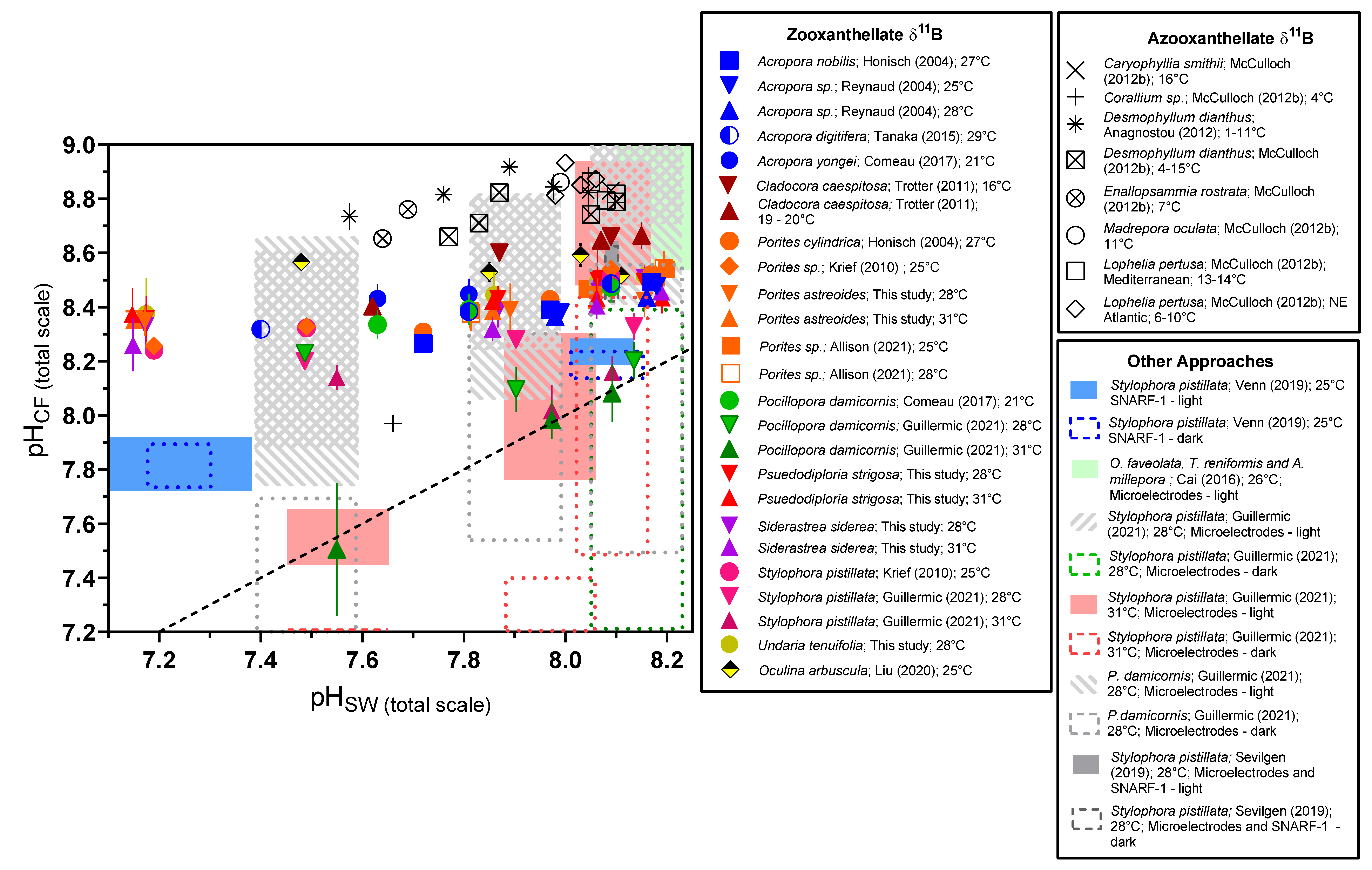
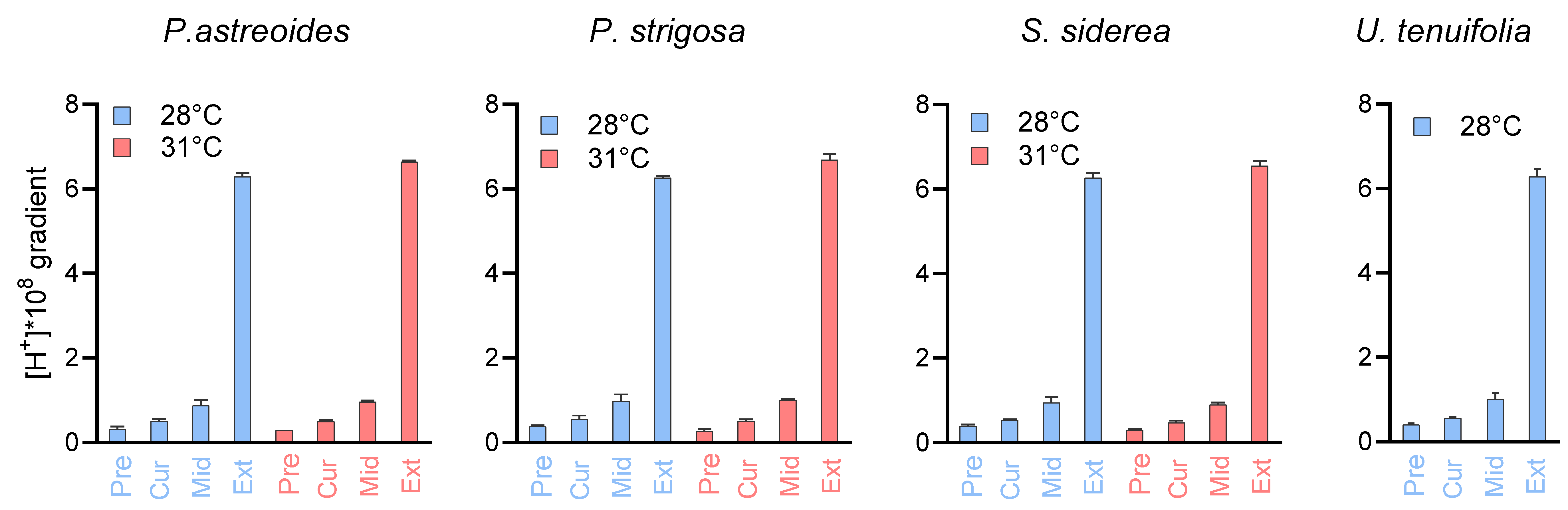
3.3. Biological Compensation for External pH Changes, and the Importance of Coral and Symbiont Physiology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glynn, P.W. Coral reef bleaching in the 1980s and possible connections with global warming. Trends Ecol. Evol. 1991, 6, 175–179. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Doney, S.C.; Fabry, V.J.; Feely, R.A.; Kleypas, J.A. Ocean Acidification: The Other CO2 Problem. Annu. Rev. Mar. Sci. 2009, 1, 169–192. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Rodriguez, M.D.; Halloran, P.R.; Rickaby, R.E.M.; Hall, I.R.; Colmenero-Hidalgo, E.; Gittins, J.R.; Green, D.R.H.; Tyrrell, T.; Gibbs, S.J.; von Dassow, P.; et al. Phytoplankton Calcification in a High-CO2 World. Science 2008, 320, 336–340. [Google Scholar] [CrossRef]
- Ries, J.B.; Cohen, A.L.; McCorkle, D.C. Marine calcifiers exhibit mixed responses to CO2-induced ocean acidification. Geology 2009, 37, 1131–1134. [Google Scholar] [CrossRef]
- Kroeker, K.; Micheli, F.; Gambi, M.C. Ocean acidification causes ecosystem shifts via altered competitive interactions. Nat. Clim. Chang. 2012, 3, 156–159. [Google Scholar] [CrossRef]
- Guillermic, M.; Cameron, L.P.; De Corte, I.; Misra, S.; Bijma, J.; de Beer, D.; Reymond, C.E.; Westphal, H.; Ries, J.B.; Eagle, R.A. Thermal stress reduces pocilloporid coral resilience to ocean acidification by impairing control over calcifying fluid chemistry. Sci. Adv. 2021, 7, eaba9958. [Google Scholar] [CrossRef]
- Marubini, F.; Atkinson, M. Effects of lowered pH and elevated nitrate on coral calcification. Mar. Ecol. Prog. Ser. 1999, 188, 117–121. [Google Scholar] [CrossRef]
- Langdon, C.; Takahashi, T.; Sweeney, C.; Chipman, D.; Goddard, J.; Marubini, F.; Aceves, H.; Barnett, H.; Atkinson, M.J. Effect of calcium carbonate saturation state on the calcification rate of an experimental coral reef. Glob. Biogeochem. Cycles 2000, 14, 639–654. [Google Scholar] [CrossRef]
- Langdon, C. Effect of elevated pCO2 on photosynthesis and calcification of corals and interactions with seasonal change in temperature/irradiance and nutrient enrichment. J. Geophys. Res. Earth Surf. 2005, 110, C9. [Google Scholar] [CrossRef]
- Marubini, F.; Barnett, H.; Langdon, C.; Atkinson, M.J. Dependence of calcification on light and carbonate ion concentration for the hermatypic coral Porites compressa. Mar. Ecol. Prog. Ser. 2001, 220, 153–162. [Google Scholar] [CrossRef]
- Renegar, D.; Riegl, B. Effect of nutrient enrichment and elevated CO2 partial pressure on growth rate of Atlantic scleractinian coral Acropora cervicornis. Mar. Ecol. Prog. Ser. 2005, 293, 69–76. [Google Scholar] [CrossRef]
- Schneider, K.; Erez, J. The effect of carbonate chemistry on calcification and photosynthesis in the hermatypic coral Acropora eurystoma. Limnol. Oceanogr. 2006, 51, 1284–1293. [Google Scholar] [CrossRef]
- Anthony, K.R.N.; Kline, D.I.; Diaz-Pulido, G.; Dove, S.; Hoegh-Guldberg, O. Ocean acidification causes bleaching and productivity loss in coral reef builders. Proc. Natl. Acad. Sci. USA 2008, 105, 17442–17446. [Google Scholar] [CrossRef] [PubMed]
- Jokiel, P.L.; Rodgers, K.S.; Kuffner, I.B.; Andersson, A.J.; Cox, E.F.; Mackenzie, F.T. Ocean acidification and calcifying reef organisms: A mesocosm investigation. Coral Reefs 2008, 27, 473–483. [Google Scholar] [CrossRef]
- Marubini, F.; Ferrier-Pagès, C.; Furla, P.; Allemand, D. Coral calcification responds to seawater acidification: A working hypothesis towards a physiological mechanism. Coral Reefs 2008, 27, 491–499. [Google Scholar] [CrossRef]
- Krief, S.; Hendy, E.J.; Fine, M.; Yam, R.; Meibom, A.; Foster, G.L.; Shemesh, A. Physiological and isotopic responses of scleractinian corals to ocean acidification. Geochim. Cosmochim. Acta 2010, 74, 4988–5001. [Google Scholar] [CrossRef]
- Holcomb, M.; Cohen, A.L.; McCorkle, D.C. An investigation of the calcification response of the scleractinian coral Astrangia poculata to elevated pCO2 and the effects of nutrients, zooxanthellae and gender. Biogeosciences 2012, 9, 29–39. [Google Scholar] [CrossRef]
- Edmunds, P.J.; Brown, D.; Moriarty, V. Interactive effects of ocean acidification and temperature on two scleractinian corals from Moorea, French Polynesia. Glob. Chang. Biol. 2012, 18, 2173–2183. [Google Scholar] [CrossRef]
- Comeau, S.; Carpenter, R.C.; Edmunds, P.J. Effects of irradiance on the response of the coral Acropora pulchra and the calcifying alga Hydrolithon reinboldii to temperature elevation and ocean acidification. J. Exp. Mar. Biol. Ecol. 2014, 453, 28–35. [Google Scholar] [CrossRef]
- Reynaud, S.; Leclercq, N.; Romaine-Lioud, S.; Ferrier-Pagés, C.; Jaubert, J.; Gattuso, J.-P. Interacting effects of CO2 partial pressure and temperature on photosynthesis and calcification in a scleractinian coral. Glob. Chang. Biol. 2003, 9, 1660–1668. [Google Scholar] [CrossRef]
- Rodolfo-Metalpa, R.; Martin, S.; Ferrier-Pagès, C.; Gattuso, J.-P. Response of the temperate coral Cladocora caespitosa to mid- and long-term exposure to pCO2 and temperature levels projected for the year 2100 AD. Biogeosciences 2010, 7, 289–300. [Google Scholar] [CrossRef]
- Edmunds, P.J. Zooplanktivory ameliorates the effects of ocean acidification on the reef coral Porites spp. Limnol. Oceanogr. 2011, 56, 2402–2410. [Google Scholar] [CrossRef]
- Schoepf, V.; Grottoli, A.G.; Warner, M.E.; Cai, W.-J.; Melman, T.F.; Hoadley, K.D.; Pettay, D.T.; Hu, X.; Li, Q.; Xu, H.; et al. Coral Energy Reserves and Calcification in a High-CO2 World at Two Temperatures. PLoS ONE 2013, 8, e75049. [Google Scholar] [CrossRef] [PubMed]
- Cameron, L.P.; Reymond, C.E.; Bijma, J.; Büscher, J.; De Beer, D.; Guillermic, M.; Eagle, R.A.; Gunnel, J.; Müller-Lundin, F.; Schmidt-Greib, G.; et al. Impacts of warming and acidification on coral calcification linked to symbiont loss and deregulation of calcifying fluid pH. J. Mar. Sci. Eng. 2022, unpublished. [Google Scholar]
- Innis, T.; Allen-Waller, L.; Brown, K.T.; Sparagon, W.; Carlson, C.; Kruse, E.; Huffmyer, A.S.; Nelson, C.E.; Putnam, H.M.; Barott, K.L. Marine heatwaves depress metabolic activity and impair cellular acid–base homeostasis in reef-building corals regardless of bleaching susceptibility. Glob. Chang. Biol. 2021, 27, 2728–2743. [Google Scholar] [CrossRef]
- D’Olivo, J.P.; McCulloch, M. Response of coral calcification and calcifying fluid composition to thermally induced bleaching stress. Sci. Rep. 2017, 7, 2207. [Google Scholar] [CrossRef]
- D’Olivo, J.P.; Ellwood, G.; DeCarlo, T.M.; McCulloch, M. Deconvolving the long-term impacts of ocean acidification and warming on coral biomineralisation. Earth Planet. Sci. Lett. 2019, 526, 115785. [Google Scholar] [CrossRef]
- Schoepf, V.; D’Olivo, J.P.; Rigal, C.; Jung, E.M.U.; McCulloch, M.T. Heat stress differentially impacts key calcification mechanisms in reef-building corals. Coral Reefs 2021, 40, 459–471. [Google Scholar] [CrossRef]
- Venn, A.; Tambutté, E.; Holcomb, M.; Allemand, D.; Tambutté, S. Live Tissue Imaging Shows Reef Corals Elevate pH under Their Calcifying Tissue Relative to Seawater. PLoS ONE 2011, 6, e20013. [Google Scholar] [CrossRef]
- Venn, A.A.; Tambutté, E.; Holcomb, M.; Laurent, J.; Allemand, D.; Tambutté, S. Impact of seawater acidification on pH at the tissue–skeleton interface and calcification in reef corals. Proc. Natl. Acad. Sci. USA 2012, 110, 1634–1639. [Google Scholar] [CrossRef] [PubMed]
- Allison, N.; Cohen, I.; Finch, A.A.; Erez, J.; Tudhope, A.W. Corals concentrate dissolved inorganic carbon to facilitate calcification. Nat. Commun. 2014, 5, 5741. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, M.T.; D’Olivo, J.P.; Falter, J.; Georgiou, L.; Holcomb, M.; Montagna, P.; Trotter, J.A. Boron Isotopic Systematics in Scleractinian Corals and the Role of pH Up-regulation. In Boron Isotopes. Advances in Isotope Geochemistry; Springer: Cham, Switzerland, 2017; pp. 145–162. [Google Scholar] [CrossRef]
- Venn, A.A.; Tambutté, E.; Caminiti-Segonds, N.; Techer, N.; Allemand, D. Effects of light and darkness on pH regulation in three coral species exposed to seawater acidification. Sci. Rep. 2019, 9, 2201. [Google Scholar] [CrossRef] [PubMed]
- Sevilgen, D.S.; Venn, A.A.; Hu, M.Y.; Tambutté, E.; de Beer, D.; Planas-Bielsa, V.; Tambutté, S. Full in vivo characterization of carbonate chemistry at the site of calcification in corals. Sci. Adv. 2019, 5, eaau7447. [Google Scholar] [CrossRef]
- Guo, W. Seawater temperature and buffering capacity modulate coral calcifying pH. Sci. Rep. 2019, 9, 1189. [Google Scholar] [CrossRef] [PubMed]
- Chalk, T.B.; Standish, C.D.; D’Angelo, C.; Castillo, K.D.; Milton, J.A.; Foster, G.L. Mapping coral calcification strategies from in situ boron isotope and trace element measurements of the tropical coral Siderastrea siderea. Sci. Rep. 2021, 11, 472. [Google Scholar] [CrossRef]
- Bove, C.B.; Ries, J.B.; Davies, S.W.; Westfield, I.T.; Umbanhowar, J.; Castillo, K.D. Common Caribbean corals exhibit highly variable responses to future acidification and warming. Proc. R. Soc. B Boil. Sci. 2019, 286, 20182840. [Google Scholar] [CrossRef]
- Bove, C.B.; Davies, S.W.; Ries, J.B.; Umbanhowar, J.; Thomasson, B.C.; Farquhar, E.B.; McCoppin, J.A.; Castillo, K.D. Global change differentially modulates coral physiology and suggests future shifts in Caribbean reef assemblages. bioRxiv 2021. [Google Scholar] [CrossRef]
- Castillo, K.D.; Lima, F.P. Comparison of in situ and satellite-derived (MODIS-Aqua/Terra) methods for assessing temperatures on coral reefs. Limnol. Oceanogr. Methods 2010, 8, 107–117. [Google Scholar] [CrossRef]
- Barker, S.; Greaves, M.; Elderfield, H. A study of cleaning procedures used for foraminiferal Mg/Ca paleothermometry. Geochem. Geophys. Geosyst. 2003, 4. [Google Scholar] [CrossRef]
- Gaillardet, J.; Lemarchand, D.; Göpel, C.; Manhès, G. Evaporation and Sublimation of Boric Acid: Application for Boron Purification from Organic Rich Solutions. Geostand. Geoanal. Res. 2001, 25, 67–75. [Google Scholar] [CrossRef]
- Misra, S.; Owen, R.; Kerr, J.; Greaves, M.; Elderfield, H. Determination of δ11B by HR-ICP-MS from mass limited samples: Application to natural carbonates and water samples. Geochim. Cosmochim. Acta 2014, 140, 531–552. [Google Scholar] [CrossRef]
- Guillermic, M.; Misra, S.; Eagle, R.; Villa, A.; Chang, F.; Tripati, A. Seawater pH reconstruction using boron isotopes in multiple planktonic foraminifera species with different depth habitats and their potential to constrain pH and pCO2 gradients. Biogeosciences 2020, 17, 3487–3510. [Google Scholar] [CrossRef]
- Lloyd, N.S.; Sadekov, A.Y.; Misra, S. Application of 1013 ohm Faraday cup current amplifiers for boron isotopic analyses by solution mode and laser ablation multicollector inductively coupled plasma mass spectrometry. Rapid Commun. Mass Spectrom. 2017, 32, 9–18. [Google Scholar] [CrossRef]
- McCulloch, M.T.; Holcomb, M.; Rankenburg, K.; Trotter, J.A. Rapid, high-precision measurements of boron isotopic compositions in marine carbonates. Rapid Commun. Mass Spectrom. 2014, 28, 2704–2712. [Google Scholar] [CrossRef]
- Holcomb, M.; Venn, A.A.; Tambutté, E.; Allemand, D.; Trotter, J.; McCulloch, M.; Tambutte, S. Coral calcifying fluid pH dictates response to ocean acidification. Sci. Rep. 2014, 4, 5207. [Google Scholar] [CrossRef] [PubMed]
- Sutton, J.N.; Liu, Y.-W.; Ries, J.B.; Guillermic, M.; Ponzevera, E.; Eagle, R.A. δ11B as monitor of calcification site pH in divergent marine calcifying organisms. Biogeosciences 2018, 15, 1447–1467. [Google Scholar] [CrossRef]
- Gutjahr, M.; Bordier, L.; Douville, E.; Farmer, J.; Foster, G.L.; Hathorne, E.C.; Hönisch, B.; Lemarchand, D.; Louvat, P.; McCulloch, M.; et al. Sub-Permil Interlaboratory Consistency for Solution-Based Boron Isotope Analyses on Marine Carbonates. Geostand. Geoanal. Res. 2021, 45, 59–75. [Google Scholar] [CrossRef]
- Hemming, N.; Hanson, G. Boron isotopic composition and concentration in modern marine carbonates. Geochim. Cosmochim. Acta 1992, 56, 537–543. [Google Scholar] [CrossRef]
- Decarlo, T.M.; Comeau, S.; Cornwall, C.E.; McCulloch, M.T. Coral resistance to ocean acidification linked to increased calcium at the site of calcification. Proc. R. Soc. B Boil. Sci. 2018, 285, 20180564. [Google Scholar] [CrossRef]
- Foster, G.L.; von Strandmann, P.P.; Rae, J. Boron and magnesium isotopic composition of seawater. Geochem. Geophys. Geosyst. 2010, 11. [Google Scholar] [CrossRef]
- Dickson, A.G. Thermodynamics of the dissociation of boric acid in synthetic seawater from 273.15 to 318.15 K. Deep Sea Res. Part A Oceanogr. Res. Pap. 1990, 37, 755–766. [Google Scholar] [CrossRef]
- Lee, K.; Kim, T.-W.; Byrne, R.H.; Millero, F.J.; Feely, R.A.; Liu, Y.-M. The universal ratio of boron to chlorinity for the North Pacific and North Atlantic oceans. Geochim. Cosmochim. Acta 2010, 74, 1801–1811. [Google Scholar] [CrossRef]
- Upadhyay, D.; Lucarelli, J.; Arnold, A.; Flores, R.; Bricker, H.; Ulrich, R.N.; Jesmok, G.; Santi, L.; Defliese, W.; Eagle, R.A.; et al. Carbonate clumped isotope analysis (Δ47) of 21 carbonate standards determined via gas-source isotope-ratio mass spectrometry on four instrumental configurations using carbonate-based standardization and multiyear data sets. Rapid Commun. Mass Spectrom. 2021, 35, e9143. [Google Scholar] [CrossRef] [PubMed]
- Tukey, J. Exploratory Data Analysis; Addison-Wesley: Reading, MA, USA, 1977. [Google Scholar]
- Hönisch, B.; Hemming, N.; Grottoli, A.; Amat, A.; Hanson, G.; Bijma, J. Assessing scleractinian corals as recorders for paleo-pH: Empirical calibration and vital effects. Geochim. Cosmochim. Acta 2004, 68, 3675–3685. [Google Scholar] [CrossRef]
- Reynaud, S.; Hemming, N.G.; Juillet-Leclerc, A.; Gattuso, J.-P. Effect of pCO2 and temperature on the boron isotopic composition of the zooxanthellate coral Acropora sp. Coral Reefs 2004, 23, 539–546. [Google Scholar] [CrossRef]
- Trotter, J.; Montagna, P.; McCulloch, M.; Silenzi, S.; Reynaud, S.; Mortimer, G.; Martin, S.; Ferrier-Pagès, C.; Gattuso, J.-P.; Rodolfo-Metalpa, R. Quantifying the pH ‘vital effect’ in the temperate zooxanthellate coral Cladocora caespitosa: Validation of the boron seawater pH proxy. Earth Planet. Sci. Lett. 2011, 303, 163–173. [Google Scholar] [CrossRef]
- Rollion-Bard, C.; Blamart, D.; Trebosc, J.; Tricot, G.; Mussi, A.; Cuif, J.-P. Boron isotopes as pH proxy: A new look at boron speciation in deep-sea corals using 11B MAS NMR and EELS. Geochim. Cosmochim. Acta 2011, 75, 1003–1012. [Google Scholar] [CrossRef]
- Ries, J.B. A physicochemical framework for interpreting the biological calcification response to CO2-induced ocean acidification. Geochim. Cosmochim. Acta 2011, 75, 4053–4064. [Google Scholar] [CrossRef]
- Anagnostou, E.; Huang, K.-F.; You, C.-F.; Sikes, E.; Sherrell, R. Evaluation of boron isotope ratio as a pH proxy in the deep sea coral Desmophyllum dianthus: Evidence of physiological pH adjustment. Earth Planet. Sci. Lett. 2012, 349–350, 251–260. [Google Scholar] [CrossRef]
- McCulloch, M.; Falter, J.; Trotter, J.; Montagna, P. Coral resilience to ocean acidification and global warming through pH up-regulation. Nat. Clim. Chang. 2012, 2, 623–627. [Google Scholar] [CrossRef]
- Tanaka, K.; Holcomb, M.; Takahashi, A.; Kurihara, H.; Asami, R.; Shinjo, R.; Sowa, K.; Rankenburg, K.; Watanabe, T.; McCulloch, M. Response of Acropora digitifera to ocean acidification: Constraints from δ11B, Sr, Mg, and Ba compositions of aragonitic skeletons cultured under variable seawater pH. Coral Reefs 2015, 34, 1139–1149. [Google Scholar] [CrossRef]
- Georgiou, L.; Falter, J.; Trotter, J.; Kline, D.I.; Holcomb, M.; Dove, S.G.; Hoegh-Guldberg, O.; McCulloch, M. pH homeostasis during coral calcification in a free ocean CO 2 enrichment (FOCE) experiment, Heron Island reef flat, Great Barrier Reef. Proc. Natl. Acad. Sci. USA 2015, 112, 13219–13224. [Google Scholar] [CrossRef]
- Cai, W.J.; Ma, Y.; Hopkinson, B.M.; Grottoli, A.G.; Warner, M.E.; Ding, Q.; Hu, X.; Yuan, X.; Schoepf, V.; Xu, H.; et al. Microelectrode characterization of coral daytime interior pH and carbonate chemistry. Nat. Commun. 2016, 7, 11144. [Google Scholar] [CrossRef]
- Comeau, S.; Cornwall, C.E.; McCulloch, M. Decoupling between the response of coral calcifying fluid pH and calcification to ocean acidification. Sci. Rep. 2017, 7, 7573. [Google Scholar] [CrossRef]
- Comeau, S.; Tambutté, E.; Carpenter, R.C.; Edmunds, P.J.; Evensen, N.R.; Allemand, D.; Ferrier-Pagès, C.; Venn, A.A. Coral calcifying fluid pH is modulated by seawater carbonate chemistry not solely seawater pH. Proc. R. Soc. B Boil. Sci. 2017, 284, 20161669. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-W.; Sutton, J.N.; Ries, J.B.; Eagle, R.A. Regulation of calcification site pH is a polyphyletic but not always governing response to ocean acidification. Sci. Adv. 2020, 6, eaax1314. [Google Scholar] [CrossRef]
- Allison, N.; Cole, C.; Hintz, C.; Hintz, K.; Rae, J.; Finch, A. Resolving the interactions of ocean acidification and temperature on coral calcification media pH. Coral Reefs 2021, 40, 1807–1818. [Google Scholar] [CrossRef]
- Allison, N.; Cohen, I.; Finch, A.A.; Erez, J. Controls on Sr/Ca and Mg/Ca in scleractinian corals: The effects of Ca-ATPase and transcellular Ca channels on skeletal chemistry. Geochim. Cosmochim. Acta 2011, 75, 6350–6360. [Google Scholar] [CrossRef]
- Pierrot, D.; Lewis, E.; Wallace, D.W.R. MS Excel Program Developed for CO2 System Calculations. In Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory; U.S. Department of Energy: Oak Ridge, TN, USA, 2006. [Google Scholar] [CrossRef]
- Al-Horani, F.A.; Almoghrabi, S.M.; De Beer, D. The mechanism of calcification and its relation to photosynthesis and respiration in the scleractinian coral Galaxea fascicularis. Mar. Biol. 2003, 142, 419–426. [Google Scholar] [CrossRef]
- McCulloch, M.T.; D’Olivo, J.P.; Falter, J.; Holcomb, M.; Trotter, J. Coral calcification in a changing World and the interactive dynamics of pH and DIC upregulation. Nat. Commun. 2017, 8, 15686. [Google Scholar] [CrossRef] [PubMed]
- Allison, N. Reconstructing coral calcification fluid dissolved inorganic carbon chemistry from skeletal boron: An exploration of potential controls on coral aragonite B/Ca. Heliyon 2017, 3, e00387. [Google Scholar] [CrossRef] [PubMed]
- Mavromatis, V.; Montouillout, V.; Noireaux, J.; Gaillardet, J.; Schott, J. Characterization of boron incorporation and speciation in calcite and aragonite from co-precipitation experiments under controlled pH, temperature and precipitation rate. Geochim. Cosmochim. Acta 2015, 150, 299–313. [Google Scholar] [CrossRef]
- Holcomb, M.; DeCarlo, T.; Gaetani, G.; McCulloch, M. Factors affecting B/Ca ratios in synthetic aragonite. Chem. Geol. 2016, 437, 67–76. [Google Scholar] [CrossRef]
- Mass, T.; Giuffre, A.J.; Sun, C.-Y.; Stifler, C.A.; Frazier, M.J.; Neder, M.; Tamura, N.; Stan, C.V.; Marcus, M.A.; Gilbert, P.U.P.A. Amorphous calcium carbonate particles form coral skeletons. Proc. Natl. Acad. Sci. USA 2017, 114, E7670–E7678. [Google Scholar] [CrossRef]
- Evans, D.; Gray, W.R.; Rae, J.; Greenop, R.; Webb, P.B.; Penkman, K.; Kröger, R.; Allison, N. Trace and major element incorporation into amorphous calcium carbonate (ACC) precipitated from seawater. Geochim. Cosmochim. Acta 2020, 290, 293–311. [Google Scholar] [CrossRef]
- Ulrich, R.N.; Guillermic, M.; Campbell, J.; Hakim, A.; Han, R.; Singh, S.; Stewart, J.D.; Román-Palacios, C.; Carroll, H.M.; De Corte, I.; et al. Patterns of Element Incorporation in Calcium Carbonate Biominerals Recapitulate Phylogeny for a Diverse Range of Marine Calcifiers. Front. Earth Sci. 2021, 9, 641760. [Google Scholar] [CrossRef]
- Giuffre, A.J.; Gagnon, A.C.; De Yoreo, J.J.; Dove, P.M. Isotopic tracer evidence for the amorphous calcium carbonate to calcite transformation by dissolution–reprecipitation. Geochim. Cosmochim. Acta 2015, 165, 407–417. [Google Scholar] [CrossRef]
- Von Euw, S.; Zhang, Q.; Manichev, V.; Murali, N.; Gross, J.; Feldman, L.C.; Gustafsson, T.; Flach, C.; Mendelsohn, R.; Falkowski, P.G. Biological control of aragonite formation in stony corals. Science 2017, 356, 933–938. [Google Scholar] [CrossRef]
- Swart, P.; Leder, J.; Szmant, A.; Dodge, R. The origin of variations in the isotopic record of scleractinian corals: II. Carbon. Geochim. Cosmochim. Acta 1996, 60, 2871–28856. [Google Scholar] [CrossRef]
- Grottoli, A.G. Effect of light and brine shrimp on skeletal δ13C in the Hawaiian coral Porites compressa: A tank experiment. Geochim. Cosmochim. Acta 2002, 66, 1955–1967. [Google Scholar] [CrossRef]
- Baker, A.C. Reef corals bleach to survive change. Nature 2001, 411, 765–766. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.M.; Freeman, C.J.; Wong, J.C.; Fogel, M.L.; Knowlton, N. Climate change promotes parasitism in a coral symbiosis. ISME J. 2018, 12, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Castillo, K.D.; Ries, J.B.; Bruno, J.F.; Westfield, I.T. The reef-building coral Siderastrea siderea exhibits parabolic responses to ocean acidification and warming. Proc. R. Soc. B Boil. Sci. 2014, 281, 20141856. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, R.R.; Towle, E.K.; Hooidonk, R.; Mor, C.; Winter, R.N.; Piggot, A.M.; Cunning, R.; Baker, A.; Klaus, J.S.; Swart, P.K.; et al. Species-specific responses to climate change and community composition determine future calcification rates of Florida Keys reefs. Glob. Chang. Biol. 2016, 23, 1023–1035. [Google Scholar] [CrossRef]
- Bove, C.B.; Umbanhowar, J.; Castillo, K.D. Meta-Analysis Reveals Reduced Coral Calcification Under Projected Ocean Warming but Not Under Acidification Across the Caribbean Sea. Front. Mar. Sci. 2020, 7, 127. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
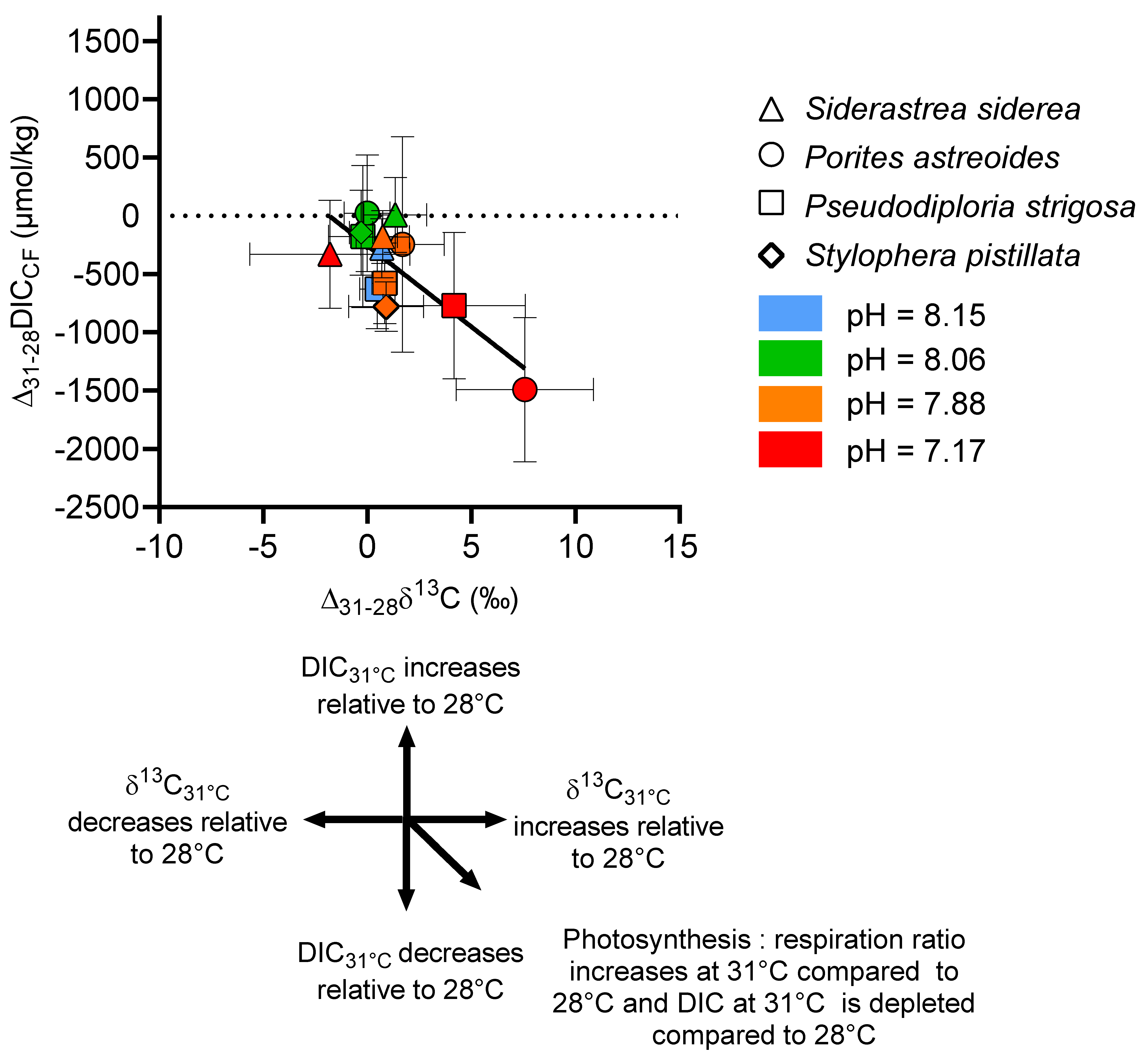{kind=link}
| Species | T (°C) | CO2 (μatm) | δ11B (‰) 1 | Error (SD) | N * | B/Ca (μmol/mol) | Error (SD) | N | δ13C (‰) 2 | Error (SD) | N |
|---|---|---|---|---|---|---|---|---|---|---|---|
| P. astreoides | |||||||||||
| 28 | 311 | 23.2 | 0.9 | 6 | 396 | 54 | 6 | −2.3 | 0.5 | 4 | |
| 28 | 405 | 23.6 | 0.9 | 6 | 437 | 31 | 6 | −2.6 | 0.9 | 4 | |
| 28 | 702 | 22.7 | 1.4 | 5 | 432 | 53 | 4 | −5.2 | 1.5 | 4 | |
| 28 | 3309 | 22.1 | 0.8 | 6 | 410 | 46 | 6 | 12.5 | 3.1 | 3 | |
| 31 | 288 | 24.1 | 0.1 | 1 | 389 | 17 | 1 | −1.4 | 0.0 | 1 | |
| 31 | 442 | 23.4 | 1.3 | 5 | 424 | 32 | 5 | −2.6 | 0.7 | 4 | |
| 31 | 674 | 23.1 | 0.4 | 5 | 443 | 82 | 5 | −3.5 | 1.3 | 4 | |
| 31 | 3285 | 22.6 | 0.5 | 2 | 568 | 51 | 2 | −5.0 | 1.2 | 2 | |
| P. strigosa | |||||||||||
| 28 | 312 | 24.2 | 0.6 | 6 | 496 | 43 | 6 | −2.2 | 0.6 | 5 | |
| 28 | 406 | 24.5 | 1.5 | 5 | 495 | 70 | 4 | −1.1 | 1.5 | 3 | |
| 28 | 702 | 23.3 | 0.7 | 6 | 481 | 35 | 6 | −4.0 | 0.6 | 4 | |
| 28 | 3320 | 22.2 | 1.3 | 6 | 455 | 41 | 6 | 10.9 | 1.6 | 4 | |
| 31 | 288 | 23.9 | 0.9 | 4 | 527 | 44 | 4 | −1.8 | 0.6 | 4 | |
| 31 | 443 | 23.9 | 0.8 | 5 | 495 | 50 | 5 | −1.3 | 0.7 | 4 | |
| 31 | 674 | 23.7 | 0.5 | 4 | 528 | 33 | 4 | −3.4 | 0.4 | 4 | |
| 31 | 3284 | 23.0 | 1.5 | 4 | 530 | 54 | 2 | −6.4 | 2.9 | 4 | |
| S. siderea | |||||||||||
| 28 | 312 | 24.5 | 0.7 | 6 | 450 | 26 | 6 | −3.3 | 0.2 | 4 | |
| 28 | 405 | 23.9 | 0.5 | 6 | 436 | 18 | 6 | −3.9 | 0.6 | 4 | |
| 28 | 703 | 22.7 | 0.9 | 6 | 424 | 21 | 6 | −6.2 | 1.1 | 4 | |
| 28 | 3317 | 21.8 | 1.2 | 6 | 415 | 27 | 6 | 12.5 | 1.7 | 3 | |
| 31 | 288 | 24.2 | 0.4 | 5 | 467 | 11 | 5 | −2.6 | 0.2 | 3 | |
| 31 | 449 | 23.4 | 0.7 | 6 | 425 | 22 | 6 | −2.6 | 1.4 | 4 | |
| 31 | 673 | 22.1 | 0.7 | 5 | 432 | 10 | 4 | −5.5 | 1.4 | 3 | |
| 31 | 3285 | 21.3 | 1.4 | 6 | 428 | 21 | 6 | 14.3 | 2.0 | 3 | |
| U. tenuifolia | |||||||||||
| 28 | 312 | 24.59 | 0.68 | 4 | 490 | 51 | 4 | −2.2 | 0.4 | 4 | |
| 28 | 404 | 24.25 | 0.56 | 4 | 518 | 32 | 4 | −2.6 | 0.3 | 4 | |
| 28 | 698 | 23.54 | 1.25 | 4 | 464 | 76 | 4 | −3.8 | 1.0 | 4 | |
| 28 | 3303 | 22.53 | 1.94 | 4 | 461 | 62 | 4 | −7.2 | 2.8 | 4 | |
| Geochemical Parameter | Species | Experimental Variable | ANOVA p-Value |
|---|---|---|---|
| δ11B | P. astreoides | Temperature | 0.1171 |
| CO2 | 0.0152 | ||
| Temperature + CO2 | 0.6094 | ||
| P. strigosa | Temperature | 0.5521 | |
| CO2 | 0.00532 | ||
| Temperature + CO2 | 0.59757 | ||
| S. siderea | Temperature | 0.36 | |
| CO2 | 7.58 × 10−10 | ||
| Temperature + CO2 | 0.441 | ||
| U. tenuifolia | CO2 | 0.322 | |
| δ13C | P. astreoides | Temperature | 0.152 |
| CO2 | 4.12 × 10−5 | ||
| Temperature + CO2 | 0.193 | ||
| P. strigosa | Temperature | 0.5521 | |
| CO2 | 0.00532 | ||
| Temperature + CO2 | 0.59757 | ||
| S. siderea | Temperature | 0.36 | |
| CO2 | 7.58 × 10−10 | ||
| Temperature + CO2 | 0.441 | ||
| U. tenuifolia | CO2 | 0.322 | |
| B/Ca | P. astreoides | Temperature | 0.22201 |
| CO2 | 0.78951 | ||
| Temperature + CO2 | 0.00939 | ||
| P. strigosa | Temperature | 0.00911 | |
| CO2 | 0.74462 | ||
| Temperature + CO2 | 0.16759 | ||
| S. siderea | Temperature | 0.17474 | |
| CO2 | 0.00103 | ||
| Temperature + CO2 | 0.84438 | ||
| U. tenuifolia | CO2 | 0.484 |
| Species | T (°C) | CO2 (μatm) | Net Calcification (mg cm−2 d−1) | Error (SD) | N 1 | pHCF | Error (SD) | DICCF (mmol/mol) | Error (SD) | [CO32−]CF (mmol/mol) | Error (SD) | ΩCF 2 | Error (SD) | N |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. astreoides | ||||||||||||||
| 28 | 311 | −0.08 | 0.26 | 11 | 8.42 | 0.06 | 4983 | 863 | 1131 | 181 | 20.9 | 3.4 | 6 | |
| 28 | 405 | 0.07 | 0.52 | 12 | 8.45 | 0.06 | 4421 | 352 | 1053 | 81 | 19.5 | 1.5 | 6 | |
| 28 | 702 | −0.17 | 0.4 | 10 | 8.39 | 0.10 | 4561 | 618 | 957 | 180 | 17.8 | 3.3 | 4 | |
| 28 | 3309 | −0.65 | 0.5 | 12 | 8.35 | 0.05 | 4841 | 512 | 969 | 102 | 17.9 | 1.9 | 6 | |
| 31 | 288 | 0.5 | 0.5 | 6 | 8.45 | 0 | 4777 | 0 | 1242 | 0 | 23 | 0 | 1 | |
| 31 | 442 | 0.04 | 0.29 | 8 | 8.40 | 0.09 | 4434 | 394 | 1055 | 107 | 19.5 | 2 | 5 | |
| 31 | 674 | −0.02 | 0.24 | 9 | 8.38 | 0.02 | 4365 | 703 | 1004 | 133 | 18.6 | 2.5 | 5 | |
| 31 | 3285 | −0.54 | 0.27 | 3 | 8.35 | 0.03 | 3356 | 286 | 736 | 106 | 13.6 | 2 | 2 | |
| P. strigosa | ||||||||||||||
| 28 | 312 | 0.92 | 1.28 | 13 | 8.49 | 0.04 | 3862 | 362 | 989 | 59 | 18.3 | 1.1 | 6 | |
| 28 | 406 | 1 | 1.12 | 6 | 8.50 | 0.10 | 3881 | 590 | 1013 | 85 | 18.7 | 1.6 | 4 | |
| 28 | 702 | 0.66 | 0.79 | 14 | 8.43 | 0.04 | 4025 | 311 | 933 | 47 | 17.3 | 0.9 | 6 | |
| 28 | 3320 | 0.03 | 0.71 | 16 | 8.36 | 0.08 | 4339 | 452 | 876 | 68 | 16.2 | 1.3 | 6 | |
| 31 | 288 | 0.41 | 0.3 | 9 | 8.43 | 0.06 | 3269 | 9 | 889 | 12 | 16.4 | 0.2 | 2 | |
| 31 | 443 | −0.32 | 0.74 | 6 | 8.43 | 0.05 | 3782 | 399 | 959 | 134 | 17.8 | 2.5 | 5 | |
| 31 | 674 | −0.23 | 0.55 | 7 | 8.42 | 0.03 | 3450 | 109 | 850 | 78 | 15.7 | 1.4 | 3 | |
| 31 | 3284 | −0.5 | 0.39 | 8 | 8.37 | 0.10 | 3609 | 423 | 759 | 35 | 14 | 0.6 | 2 | |
| S. siderea | ||||||||||||||
| 28 | 312 | 1.30 | 0.57 | 10 | 8.51 | 0.05 | 4216 | 261 | 1115 | 76 | 20.6 | 1.4 | 6 | |
| 28 | 405 | 1.37 | 0.70 | 12 | 8.47 | 0.03 | 4391 | 192 | 1087 | 63 | 20.1 | 1.2 | 6 | |
| 28 | 703 | 1.08 | 0.45 | 11 | 8.39 | 0.06 | 4596 | 194 | 993 | 129 | 18.4 | 2.4 | 6 | |
| 28 | 3317 | 0.13 | 0.52 | 11 | 8.33 | 0.08 | 4765 | 333 | 926 | 133 | 17.1 | 2.5 | 6 | |
| 31 | 288 | 1.00 | 0.54 | 8 | 8.46 | 0.03 | 3969 | 117 | 1033 | 34 | 19.1 | 0.6 | 4 | |
| 31 | 449 | 1.16 | 0.70 | 11 | 8.41 | 0.05 | 4407 | 252 | 1060 | 62 | 19.6 | 1.1 | 6 | |
| 31 | 673 | 1.06 | 0.51 | 11 | 8.32 | 0.05 | 4449 | 120 | 909 | 47 | 16.8 | 0.9 | 3 | |
| 31 | 3285 | 0.43 | 0.53 | 12 | 8.26 | 0.10 | 4480 | 284 | 851 | 119 | 15.7 | 2.2 | 5 | |
| U. tenuifolia | ||||||||||||||
| 28 | 312 | 0.16 | 0.10 | 11 | 8.52 | 0.04 | 3886 | 449 | 1037 | 85 | 19.2 | 1.6 | 4 | |
| 28 | 404 | 0.24 | 0.17 | 7 | 8.49 | 0.04 | 3676 | 188 | 950 | 102 | 17.6 | 1.9 | 4 | |
| 28 | 698 | 0.05 | 0.08 | 4 | 8.44 | 0.08 | 4230 | 767 | 996 | 99 | 18.5 | 1.8 | 4 | |
| 28 | 3303 | −0.23 | 0.05 | 5 | 8.38 | 0.13 | 4286 | 657 | 908 | 198 | 16.8 | 3.7 | 4 | |
| Parameter/Species/Experimental Variable | |||
|---|---|---|---|
| p Value | p Value | ||
| Symbiont cell density | pHCF | ||
| P. astreoides | P. astreoides | ||
| Temperature | 0.417 | Temperature | 0.676 |
| CO2 | 0.11 | CO2 | 6.54 × 10−5 |
| Temperature + CO2 | 0.466 | Temperature + CO2 | 0.286 |
| P. strigosa | P. strigosa | ||
| Temperature | 2.59 × 10−11 | Temperature | 0.42275 |
| CO2 | 0.648 | CO2 | 0.00536 |
| Temperature: CO2 | 0.15 | Temperature + CO2 | 0.62005 |
| S. siderea | S. siderea | ||
| Temperature | 9.09 × 10−5 | Temperature | Not Normal * |
| CO2 | 0.000251 | CO2 | Not Normal * |
| Temperature + CO2 | 0.858348 | Temperature + CO2 | Not Normal * |
| U. tenuifolia | U. tenuifolia | ||
| CO2 | 0.727 | CO2 | 0.317 |
| Chl a | [CO32−] | ||
| P. astreoides | P. astreoides | ||
| Temperature | 9.89 × 10−7 | Temperature | 0.41309 |
| CO2 | 6.10 × 10−11 | CO2 | 0.00333 |
| Temperature + CO2 | 2.80 × 10−9 | Temperature + CO2 | 0.22932 |
| P. strigosa | P. strigosa | ||
| Temperature | 6.15 × 10−8 | Temperature | 0.0988 |
| CO2 | 0.000217 | CO2 | 0.0224 |
| Temperature: CO2 | 0.070441 | Temperature: CO2 | 0.855 |
| S. siderea | S. siderea | ||
| Temperature | 0.00394 | Temperature | 0.679 |
| CO2 | 1.37 × 10−7 | CO2 | 5.98 × 10−5 |
| Temperature + CO2 | 0.25756 | Temperature + CO2 | 0.996 |
| U. tenuifolia | U. tenuifolia | ||
| CO2 | 0.033 | CO2 | 0.181 |
| Total host energy | [DIC]CF | ||
| P. astreoides | P. astreoides | ||
| Temperature | 0.02755 | Temperature | 0.00163 |
| CO2 | 0.00659 | CO2 | 0.69095 |
| Temperature: CO2 | 0.06197 | Temperature + CO2 | 0.04313 |
| P. strigosa | P. strigosa | ||
| Temperature | 8.35 × 10−5 | Temperature | 0.00395 |
| CO2 | 0.103 | CO2 | 0.26074 |
| Temperature + CO2 | 0.857 | Temperature + CO2 | 0.3616 |
| S. siderea | S. siderea | ||
| Temperature | 0.000522 | Temperature | 0.00119 |
| CO2 | 0.118746 | CO2 | 2.62 × 10−5 |
| Temperature + CO2 | 0.710086 | Temperature + CO2 | 0.3765 |
| U. tenuifolia | U. tenuifolia | ||
| CO2 | 0.168 | CO2 | 0.463 |
| Calcification rate | |||
| P. astreoides | |||
| Temperature | 0.0178 | ||
| CO2 | 5.11 × 10−6 | ||
| Temperature + CO2 | 0.1779 | ||
| P. strigosa | |||
| Temperature | 0.000142 | ||
| CO2 | 0.000364 | ||
| Temperature + CO2 | 0.678234 | ||
| S. siderea | |||
| Temperature | 0.52 | ||
| CO2 | 3.27 × 10−8 | ||
| Temperature + CO2 | 0.209 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eagle, R.A.; Guillermic, M.; De Corte, I.; Alvarez Caraveo, B.; Bove, C.B.; Misra, S.; Cameron, L.P.; Castillo, K.D.; Ries, J.B. Physicochemical Control of Caribbean Coral Calcification Linked to Host and Symbiont Responses to Varying pCO2 and Temperature. J. Mar. Sci. Eng. 2022, 10, 1075. https://doi.org/10.3390/jmse10081075
Eagle RA, Guillermic M, De Corte I, Alvarez Caraveo B, Bove CB, Misra S, Cameron LP, Castillo KD, Ries JB. Physicochemical Control of Caribbean Coral Calcification Linked to Host and Symbiont Responses to Varying pCO2 and Temperature. Journal of Marine Science and Engineering. 2022; 10(8):1075. https://doi.org/10.3390/jmse10081075
Chicago/Turabian StyleEagle, Robert A., Maxence Guillermic, Illian De Corte, Blanca Alvarez Caraveo, Colleen B. Bove, Sambuddha Misra, Louise P. Cameron, Karl D. Castillo, and Justin B. Ries. 2022. "Physicochemical Control of Caribbean Coral Calcification Linked to Host and Symbiont Responses to Varying pCO2 and Temperature" Journal of Marine Science and Engineering 10, no. 8: 1075. https://doi.org/10.3390/jmse10081075
APA StyleEagle, R. A., Guillermic, M., De Corte, I., Alvarez Caraveo, B., Bove, C. B., Misra, S., Cameron, L. P., Castillo, K. D., & Ries, J. B. (2022). Physicochemical Control of Caribbean Coral Calcification Linked to Host and Symbiont Responses to Varying pCO2 and Temperature. Journal of Marine Science and Engineering, 10(8), 1075. https://doi.org/10.3390/jmse10081075