
Effects of Iron Valence on the Growth, Photosynthesis, and Fatty Acid Composition of Phaeodactylum tricornutum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Strain and Medium Composition
2.2. Culture and Growth
2.3. Photosynthesis
2.4. Photosynthetic Pigments
2.5. Fatty Acid Analysis
2.6. Statistical Analysis
3. Results
3.1. Cell Density
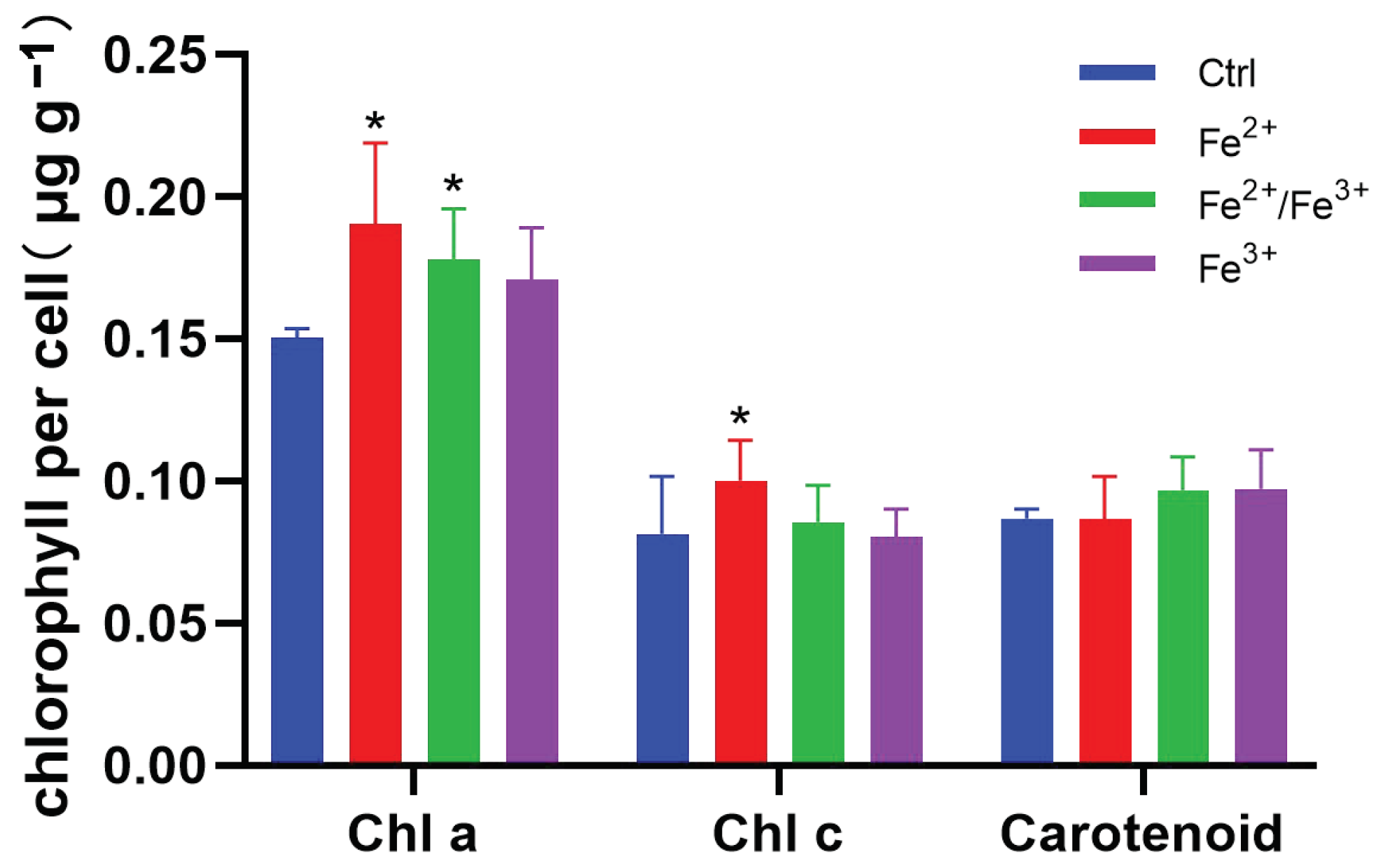
3.2. Photosynthetic Pigments
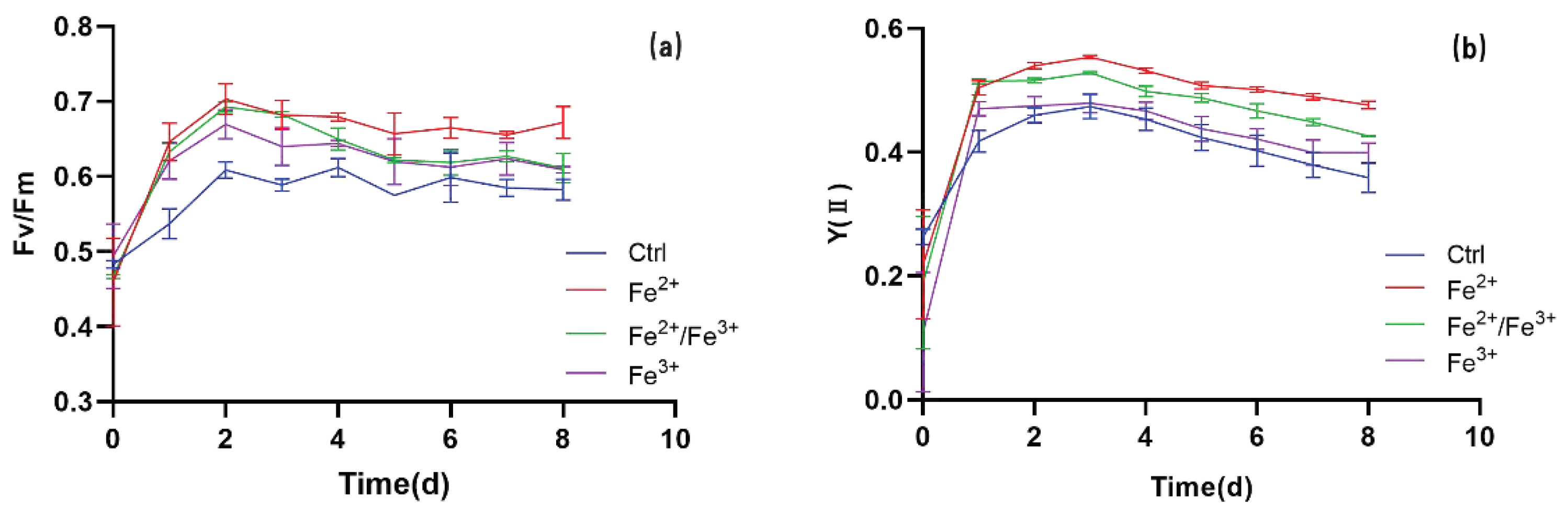
3.3. Chl Fluorescence
3.4. Total Fatty Acid Content (TFAC) and Fatty Acid Composition
4. Discussion
4.1. Effects of Different Iron Valences on the Growth of P. tricornutum
4.2. Effects of Different Iron Valences on Photosynthesis of P. tricornutum
4.3. Effects of Different Iron Valences on Lipid Accumulation and Fatty Acid Composition of P. tricornutum
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duverger, A.; Berg, J.S.; Busigny, V.; Guyot, F.; Bernard, S.; Miot, J. Mechanisms of Pyrite Formation Promoted by Sulfate-Reducing Bacteria in Pure Culture. Front. Earth Sci. 2020, 8, 588310. [Google Scholar] [CrossRef]
- Hudson, R.J.M.; Morel, F.M.M. lron transport in marine phytoplankton: Kinetics of cellular and medium coordination reactions. Limnol. Oceanogr. 1990, 35, 1002–1020. [Google Scholar] [CrossRef]
- Cook, G.; Teufel, A.; Kalra, I.; Li, W.; Wang, X.; Priscu, J.; Morgan-Kiss, R. The Antarctic psychrophiles Chlamydomonas spp. UWO241 and ICE-MDV exhibit differential restructuring of photosystem I in response to iron. Photosynth. Res. 2019, 141, 209–228. [Google Scholar] [CrossRef]
- Terauchi, A.M.; Peers, G.; Kobayashi, M.C.; Niyogi, K.K.; Merchant, S.S. Trophic status of Chlamydomonas reinhardtii influences the impact of iron deficiency on photosynthesis. Photosynth. Res. 2010, 105, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Rajabi Islami, H.; Assareh, R. Enhancement effects of ferric ion concentrations on growth and lipid characteristics of freshwater microalga Chlorococcum oleofaciens KF584224.1 for biodiesel production. Renew. Energy 2020, 149, 264–272. [Google Scholar] [CrossRef]
- de Baar, H.; Gieskes, W.; van Leeuwe, M.; van Oijen, T. Effects of iron limitation on photosynthesis and carbohydrate metabolism in the Antarctic diatom Chaetoceros brevis (Bacillariophyceae). Eur. J. Phycol. 2004, 39, 161–171. [Google Scholar] [CrossRef]
- Das, P.; Thaher, M.; Abdulquadir, M.; Al-jabri, H.; Alghasal, G. Optimization of iron dosage for microalgal biomass production as a feedstock for biofuel. Biofuels 2019, 12, 569–577. [Google Scholar] [CrossRef]
- Polat, E.; Yüksel, E.; Altınbaş, M. Effect of different iron sources on sustainable microalgae-based biodiesel production using Auxenochlorella protothecoides. Renew. Energy 2020, 162, 1970–1978. [Google Scholar] [CrossRef]
- Rizwan, M.; Mujtaba, G.; Lee, K. Effects of iron sources on the growth and lipid/carbohydrate production of marine microalga Dunaliella tertiolecta. Biotechnol. Bioprocess Eng. 2017, 22, 68–75. [Google Scholar] [CrossRef]
- Botebol, H.; Sutak, R.; Scheiber, I.F.; Blaiseau, P.L.; Bouget, F.Y.; Camadro, J.M.; Lesuisse, E. Different iron sources to study the physiology and biochemistry of iron metabolism in marine micro-algae. Biometals 2014, 27, 75–88. [Google Scholar] [CrossRef]
- Qiu, Y.; Wang, Z.; Liu, F.; Liu, J.; Zhou, T. Effect of different kinds of complex iron on the growth of Anabaena flos-aquae. Environ. Technol. 2019, 40, 2889–2896. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos-Hernandez, B.; Beltrán-López, J.; Rosales-Mendoza, S. Production of biopharmaceuticals in microalgae. In Handbook of Marine Microalgae; Kim, S.-K., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 281–298. [Google Scholar]
- Cui, Y.; Thomas-Hall, S.R.; Chua, E.T.; Schenk, P.M. Development of high-level omega-3 eicosapentaenoic acid (EPA) production from Phaeodactylum tricornutum. J. Phycol. 2021, 57, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Kadalag, N.L.; Pawar, P.R.; Prakash, G. Co-cultivation of Phaeodactylum tricornutum and Aurantiochytrium limacinum for polyunsaturated omega-3 fatty acids production. Bioresour. Technol. 2022, 346, 126544. [Google Scholar] [CrossRef]
- Wang, Z.; Mou, J.; Qin, Z.; He, Y.; Sun, Z.; Wang, X.; Lin, C.S.K. An auxin-like supermolecule to simultaneously enhance growth and cumulative eicosapentaenoic acid production in Phaeodactylum tricornutum. Bioresour. Technol. 2022, 345, 126564. [Google Scholar] [CrossRef] [PubMed]
- Hayward, J. Studies on the growth of Phaeodactylum tricornutum III. The Effect of iron on growth. J. Mar. Biol. Assoc. UK 1968, 48, 295–302. [Google Scholar] [CrossRef]
- Castell, C.; Díaz-Santos, E.; Heredia-Martínez, L.G.; López-Maury, L.; Ortega, J.M.; Navarro, J.A.; Roncel, M.; Hervás, M. Iron deficiency promotes the lack of photosynthetic cytochrome c(550) and affects the binding of the luminal extrinsic subunits to photosystem II in the diatom Phaeodactylum tricornutum. Int. J. Mol. Sci. 2022, 23, 2138. [Google Scholar] [CrossRef]
- Charria-Girón, E.; Amazo, V.; De Angulo, D.; Hidalgo, E.; Villegas-Torres, M.F.; Baganz, F.; Caicedo Ortega, N.H. Strategy for managing industrial anaerobic sludge through the heterotrophic cultivation of Chlorella sorokiniana: Effect of iron addition on biomass and lipid production. Bioengineering 2021, 8, 82. [Google Scholar] [CrossRef]
- Chen, Y.; Qiu, Y.; Zhang, W.; Lu, S.; Liu, T. Effect of nutrient elements on growth and lipid accumulation of Phaeodactylum tricornutum. Biomass Chem. Eng. 2011, 45, 1–6. [Google Scholar]
- Zhao, P.; Gu, W.; Huang, A.; Wu, S.; Liu, C.; Huan, L.; Gao, S.; Xie, X.; Wang, G. Effect of iron on the growth of Phaeodactylum tricornutum via photosynthesis. J. Phycol. 2018, 54, 34–43. [Google Scholar] [CrossRef]
- Kosakowska, A.; Lewandowska, J.; Stoń, J.; Burkiewicz, K. Qualitative and quantitative composition of pigments in Phaeodactylum tricornutum (Bacillariophyceae) stressed by iron. Biometals 2004, 17, 45–52. [Google Scholar] [CrossRef]
- Liang, J.; Jiang, X.; Ye, L.; Han, Q. Effects of nitrogen, phosphorus and iron on the growth, total lipid content and fatty acid composition of Phaeodactylum tricornutum mutant strain. Chin. J. Ecol. 2016, 35, 189–198. (In Chinese) [Google Scholar] [CrossRef]
- Guillard, R.R.; Ryther, J.H. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt, and Detonula confervacea (cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Bidwell, J.; Spotte, S.H. Artificial Seawaters: Formulas and Methods; Jones & Bartlett Learning: Burlington, MA, USA, 1985. [Google Scholar]
- Ralph, P.J.; Gademann, R. Rapid light curves: A powerful tool to assess photosynthetic activity. Aquat. Bot. 2005, 82, 222–237. [Google Scholar] [CrossRef]
- Tucker, A. Pigment extraction as a method of quantitative analysis of phytoplankton. Trans. Am. Microsc. Soc. 1949, 68, 21. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1, and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 8, 53–59. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1968. [Google Scholar]
- Qiao, H.; Cong, C.; Sun, C.; Li, B.; Wang, J.; Zhang, L. Effect of culture conditions on growth, fatty acid composition and DHA/EPA ratio of Phaeodactylum tricornutum. Aquaculture 2016, 452, 311–317. [Google Scholar] [CrossRef]
- Duncan, D.B.J.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Williams, L.J.; Abdi, H.H. Fisher’s Least Significant Difference (LSD) Test. 2010. Available online: https://www.graphpad.com/guides/prism/latest/statistics/stat_fishers_lsd.htm (accessed on 16 April 2022).
- Zhou, T.; Wang, Z.; Hu, Q.; Liu, L.; Luo, K. Effects of Fe2+ and Fe3+ on algal proliferation in a natural mixed algal colony in algae-rich raw water in southern China. J. Residuals Sci. Technol. 2016, 13, 15–22. [Google Scholar] [CrossRef]
- Kothamasi, D.; Kothamasi, S. Cobalt interference in iron-uptake could inhibit growth in Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2004, 20, 755–758. [Google Scholar] [CrossRef]
- Rico, M.; López, A.; Santana-Casiano, J.M.; Gonzàlez, A.G.; Gonzàlez-Dàvila, M. Variability of the phenolic profile in the diatom Phaeodactylum tricornutum growing under copper and iron stress. Limnol. Oceanogr. 2013, 58, 144–152. [Google Scholar] [CrossRef]
- Behrenfeld, M.J.; Milligan, A.J. Photophysiological expressions of iron stress in phytoplankton. Annu. Rev. Mar. Sci. 2013, 5, 217–246. [Google Scholar] [CrossRef]
- Peers, G.; Price, N.M. Copper-containing plastocyanin used for electron transport by an oceanic diatom. Nature 2006, 441, 341–344. [Google Scholar] [CrossRef]
- Allen, A.E.; Laroche, J.; Maheswari, U.; Lommer, M.; Schauer, N.; Lopez, P.J.; Finazzi, G.; Fernie, A.R.; Bowler, C. Whole-cell response of the pennate diatom Phaeodactylum tricornutum to iron starvation. Proc. Natl. Acad. Sci. USA 2008, 105, 10438–10443. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.T.; Allen, A.E.; Chong, J.S.; Lin, K.; Leus, D.; Karpenko, N.; Harris, S.L. Copper-dependent iron transport in coastal and oceanic diatoms. Limnol. Oceanogr. 2006, 51, 1729–1743. [Google Scholar] [CrossRef]
- Jung, H.-S.; Niyogi, K. Molecular analysis of photoprotection of photosynthesis. In Advances in Photosynthesis and Respiration; Springer: Berlin/Heidelberg, Germany, 2006; pp. 127–143. [Google Scholar]
- Yoshioka, N.; Imanishi, Y.; Yasuda, K.; Sakai, A. Effects of chloroplast dysfunction in a subpopulation of leaf mesophyll cells on photosynthetic and respiratory activities of a whole leaf: A study using variegated leaves of Hedera helix L. Plant Morphol. 2009, 21, 87–91. [Google Scholar] [CrossRef]
- Spiller, S.C.; Castelfranco, A.M.; Castelfranco, P.A. Effects of iron and oxygen on chlorophyll biosynthesis: I. In vivo observations on iron and oxygen-deficient plants. Plant Physiol. 1982, 69, 107–111. [Google Scholar] [CrossRef]
- Woolhouse, H.W. Toxicity and tolerance in the responses of plants to metals. In Physiological Plant Ecology III: Responses to the Chemical and Biological Environment; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1983; pp. 245–300. [Google Scholar]
- Franck, V.; Bruland, K.; Hutchins, D.; Brzezinski, M.A. Iron and zinc effects on silicic acid and nitrate uptake kinetics in three high-nutrient, low-chlorophyll (HNLC) regions. Mar. Ecol. -Prog. Ser. 2003, 252, 15–33. [Google Scholar] [CrossRef]
- Marsh, H.V., Jr.; Evans, H.J.; Matrone, G. Investigations of the role of iron in chlorophyll metabolism. II. Effect of iron deficiency on chlorophyll synthesis. Plant Physiol. 1963, 38, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Mythili, J.B.; Shivashankara, K.S. Effects of photo-oxidative loss of FeNa2EDTA and of higher iron supply on chlorophyll content, growth and propagation rate in triploid watermelon cultures. Vitr. Cell. Dev. Biol. Plant 2000, 36, 537–542. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Asp. Med. 2004, 24, 345–351. [Google Scholar] [CrossRef]
- Ortega Cabello, L.; Pérez-Méndez, H.I.; Manjarrez-Alvarez, N.; Solís-Oba, A.; López-Luna, A. Efect of iron salts on Rhodococcus sp. and Grodonia sp. on carotenoid production. Rev. Mex. Ing. Quim. 2017, 16, 1–10. [Google Scholar] [CrossRef]
- Jahn, D.; Verkamp, E.; Söll, D. Glutamyl-transfer RNA: A precursor of heme and chlorophyll biosynthesis. Trends Biochem. Sci. 1992, 17, 215–218. [Google Scholar] [CrossRef]
- Duggan, J.; Gassman, M. Induction of porphyrin synthesis in etiolated bean leaves by chelators of iron. Plant Physiol. 1974, 53, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.W.; Denney, A.; Pushnik, J.; Yu, M.H. The formation of delta-Aminolevulinate a precursor of chlorophyll, in barley and the role of iron. J. Plant Nutr. 1982, 5, 289–300. [Google Scholar] [CrossRef]
- Pushnik, J.C.; Miller, G.W.; Manwaring, J.H. The role of iron in higher plant chlorophyll biosynthesis, maintenance and chloroplast biogenesis. Plant Nutr. 1984, 7, 733–758. [Google Scholar] [CrossRef]
- Guo, R.; Lu, D.; Liu, C.; Hu, J.; Wang, P.; Dai, X. Toxic effect of nickel on microalgae Phaeodactylum tricornutum (Bacillariophyceae). Ecotoxicology 2022, 31, 746–760. [Google Scholar] [CrossRef]
- Guikema, J.A. Fluorescence induction characteristics of Anacystis nidulans during recovery from iron deficiency. J. Plant Nutr. 1985, 8, 891–908. [Google Scholar] [CrossRef]
- Vassiliev, I.R.; Kolber, Z.; Wyman, K.D.; Mauzerall, D.; Shukla, V.K.; Falkowski, P.G. Effects of iron limitation on photosystem II composition and light utilization in Dunaliella tertiolecta. Plant Physiol. 1995, 109, 963–972. [Google Scholar] [CrossRef]
- Morales, F.; Abadía, A.; Abadía, J. Chlorophyll fluorescence and photon yield of oxygen evolution in iron-deficient sugar beet (Beta vulgaris L.) leaves. Plant Physiol. 1991, 97, 886–893. [Google Scholar] [CrossRef]
- Ruban, A.V. Quantifying the efficiency of photoprotection. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2017, 372, 20160393. [Google Scholar] [CrossRef]
- Garab, G. Structural changes and non-photochemical quenching of chlorophyll a fluorescence in oxygenic photosynthetic organisms. In Non-Photochemical Quenching and Energy Dissipation in Plants, Algae and Cyanobacteria; Demmig-Adams, B., Garab, G., Adams Iii, W., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 343–371. [Google Scholar]
- Dubey, A.; Kumar, N.; Sanyal, I. Targets of NO in plastids. In Nitric Oxide in Plant Biology; Pratap Singh, V., Singh, S., Tripathi, D.K., Romero-Puertas, M.C., Sandalio, L.M., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 331–344. [Google Scholar]
- Zhu, M.; Mu, X.; Li, R. The role of iron in primary productivity in the sea. J. Oceanograpgy Huanghai Bohai Seas 1997, 15, 50–56. (In Chinese) [Google Scholar]
- Varsano, T.; Kaftan, D.; Pick, U. Effects of iron deficiency on thylakoid membrane structure and composition in the alga Dunaliella salina. Plant Nutr. 2003, 26, 2197–2210. [Google Scholar] [CrossRef]
- Malone, L.A.; Proctor, M.S.; Hitchcock, A.; Hunter, C.N.; Johnson, M.P. Cytochrome b6f—Orchestrator of photosynthetic electron transfer. Biochim. Biophys. Acta Bioenerg. 2021, 1862, 148380. [Google Scholar] [CrossRef] [PubMed]
- Roach, T.; Krieger-Liszkay, A. Regulation of photosynthetic electron transport and photoinhibition. Curr. Protein Pept. Sci. 2014, 15, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Est, A. Photosynthesis: Electron transfer chemistry. In Wiley Encyclopedia of Chemical Biology; Begley, T.P., Ed.; Wiley: Hoboken, NJ, USA, 2008. [Google Scholar] [CrossRef]
- Kroh, G.E.; Pilon, M. Regulation of iron homeostasis and use in chloroplasts. Int. J. Mol. Sci. 2020, 21, 3395. [Google Scholar] [CrossRef]
- Kráľová, K.; Masarovičová, E.; Šeršeň, F.; Ondrejkovičová, I. Effect of different Fe(III) compounds on photosynthetic electron transport in spinach chloroplasts and on iron accumulation in maize plants. Chem. Pap. 2008, 62, 358. [Google Scholar] [CrossRef]
- Singh, B.; Guldhe, A.; Rawat, I.; Bux, F. Towards a sustainable approach for development of biodiesel from plant and microalgae. Renew. Sustain. Energy Rev. 2014, 29, 216–245. [Google Scholar] [CrossRef]
- Liu, Z.-Y.; Wang, G.-C.; Zhou, B.-C. Effect of iron on growth and lipid accumulation in Chlorella vulgaris. Bioresour. Technol. 2008, 99, 4717–4722. [Google Scholar] [CrossRef]
- Savvidou, M.G.; Boli, E.; Logothetis, D.; Lymperopoulou, T.; Ferraro, A.; Louli, V.; Mamma, D.; Kekos, D.; Magoulas, K.; Kolisis, F.N. A Study on the effect of macro- and micro-nutrients on Nannochloropsis oceanica growth, fatty acid composition and magnetic harvesting efficiency. Plants 2020, 9, 660. [Google Scholar] [CrossRef]
- Urzica, E.I.; Vieler, A.; Hong-Hermesdorf, A.; Page, M.D.; Casero, D.; Gallaher, S.D.; Kropat, J.; Pellegrini, M.; Benning, C.; Merchant, S.S. Remodeling of membrane lipids in iron-starved Chlamydomonas. J. Biol. Chem. 2013, 288, 30246–30258. [Google Scholar] [CrossRef]
- Devadasu, E.; Subramanyam, R. Enhanced lipid production in Chlamydomonas reinhardtii caused by severe iron deficiency. Front. Plant Sci. 2021, 12, 615577. [Google Scholar] [CrossRef] [PubMed]
- Carpio, R.; de Leon, R.; Martinez-Goss, M. Growth, lipid content, and lipid profile of the green alga, Chlorella vulgaris Beij., under different concentrations of Fe and CO2. Eng. Sci. Technol. 2015, 10, 19–30. [Google Scholar]
- Aitzetmüller, K.; Tsevegsüren, N. Seed fatty acids, «front-end»-desaturases and chemotaxonomy—A case study in the Ranunculaceae. Plant Physiol. 1994, 143, 538–543. [Google Scholar] [CrossRef]
- Kachroo, P.; Shanklin, J.; Shah, J.; Whittle, E.J.; Klessig, D.F. A fatty acid desaturase modulates the activation of defense signaling pathways in plants. Proc. Natl. Acad. Sci. USA 2001, 98, 9448–9453. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Ctrl | Fe2+ | Fe2+/Fe3+ | Fe3+ |
|---|---|---|---|---|
| α | 0.18 ± 0.01 a | 0.23 ± 0.00 b | 0.21 ± 0.01 c | 0.20 ± 0.01 c |
| Ek | 152.72 ± 1.67 a | 188.95 ± 2.12 b | 148.31 ± 18.40 a | 144.38 ± 11.68 a |
| ETRmax | 27.53 ± 1.31 a | 42.80 ± 1.08 b | 31.48 ± 3.15 c | 29.03 ± 2.14 a c |
| Fatty Acid | Ctrl | Fe2+ | Fe2+/Fe3+ | Fe3+ |
|---|---|---|---|---|
| C14:0 | 6.03 ± 0.02 | 6.24 ± 0.02 | 6.10 ± 0.00 | 6.14 ± 0.01 |
| C16:0 | 13.03 ± 0.04 a | 13.72 ± 0.08 ab | 14.42 ± 1.09 b | 14.21 ± 0.14 ab |
| C16:1n-7 | 19.97 ± 0.03 | 19.80 ± 0.80 | 21.78 ± 3.62 | 20.76 ± 0.09 |
| C16:2 | 1.19 ± 0.00 ab | 1.26 ± 0.00 b | 1.12 ± 0.01 a | 1.19 ± 0.01 ab |
| C16:3 | 0.67 ± 0.00 a | 0.59 ± 0.00 b | 0.67 ± 0.00 a | 0.60 ± 0.00 ab |
| C16:4 | 2.37 ± 0.00 | 2.34 ± 0.01 | 2.18 ± 0.07 | 2.16 ± 0.01 |
| C18:0 | 0.81 ± 0.03 | 0.74 ± 0.00 | 0.84 ± 0.01 | 0.86 ± 0.01 |
| C18:1n-9 | 7.66 ± 0.00 | 7.34 ± 0.01 | 7.61 ± 0.09 | 7.81 ± 0.27 |
| C18:1n-7 | 1.61 ± 0.01 | 1.59 ± 0.00 | 1.65 ± 0.00 | 1.69 ± 0.00 |
| C18:2n-6 | 1.80 ± 0.00 | 1.96 ± 0.01 | 1.74 ± 0.11 | 2.26 ± 0.16 |
| C18:3n-6 | 0.74 ± 0.00 a | 0.77 ± 0.00 a | 0.63 ± 0.00 b | 0.64 ± 0.00 b |
| C18:3n-3 | 0.22 ± 0.00 | 0.21 ± 0.00 | 0.21 ± 0.00 | 0.23 ± 0.00 |
| C18:4n-3 | 0.59 ± 0.00 a | 0.52 ± 0.00 b | 0.42 ± 0.00 c | 0.38 ± 0.00 c |
| C20:3n-3 | - | - | 0.04 ± 0.00 | 0.02 ± 0.00 |
| ARA | 1.02 ± 0.00 a | 1.21 ± 0.00 b | 1.15 ± 0.00 b | 1.15 ± 0.00 b |
| C20:4n-3 | 0.51 ± 0.00 ab | 0.57 ± 0.00 a | 0.50 ± 0.00 bc | 0.43 ± 0.00 c |
| C24:0 | 2.02 ± 0.00 | 2.07 ± 0.00 | 1.94 ± 0.00 | 1.97 ± 0.01 |
| EPA | 25.84 ± 0.74 a | 25.13 ± 0.12 ab | 23.86 ± 1.53 b | 23.96 ± 0.24 b |
| C22:5n-3 | 0.49 ± 0.00 a | 0.49 ± 0.00 a | 0.39 ± 0.00 b | 0.42 ± 0.00 b |
| DHA | 0.94 ± 0.00 | 0.91 ± 0.00 | 0.89 ± 0.01 | 0.91 ± 0.00 |
| others | 12.48 ± 0.33 | 12.54 ± 0.27 | 11.88 ± 0.81 | 12.21 ± 0.49 |
| SFAs | 19.87 ± 0.07 a | 20.69 ± 0.15 ab | 21.36 ± 1.24 b | 21.20 ± 0.13 b |
| MUFAs | 29.25 ± 0.03 ab | 28.73 ± 0.64 b | 31.04 ± 3.09 a | 30.26 ± 0.42 ab |
| PUFAs | 32.15 ± 0.77 a | 31.77 ± 0.33 ab | 29.82 ± 2.75 b | 30.40 ± 0.06 ab |
| DHA/EPA | 0.04 ± 0.00 | 0.04 ± 0.00 | 0.04 ± 0.00 | 0.04 ± 0.00 |
| TFAC | 87.52 ± 0.33 | 87.46 ± 0.27 | 88.12 ± 0.81 | 87.79 ± 0.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Su, Q.; Zhuang, Y.; Wu, C.; Tong, S.; Guan, B.; Zhao, Y.; Qiao, H. Effects of Iron Valence on the Growth, Photosynthesis, and Fatty Acid Composition of Phaeodactylum tricornutum. J. Mar. Sci. Eng. 2023, 11, 316. https://doi.org/10.3390/jmse11020316
Wang H, Su Q, Zhuang Y, Wu C, Tong S, Guan B, Zhao Y, Qiao H. Effects of Iron Valence on the Growth, Photosynthesis, and Fatty Acid Composition of Phaeodactylum tricornutum. Journal of Marine Science and Engineering. 2023; 11(2):316. https://doi.org/10.3390/jmse11020316
Chicago/Turabian StyleWang, Haowen, Qingling Su, Yanmin Zhuang, Chengzong Wu, Shanying Tong, Bo Guan, Yancui Zhao, and Hongjin Qiao. 2023. "Effects of Iron Valence on the Growth, Photosynthesis, and Fatty Acid Composition of Phaeodactylum tricornutum" Journal of Marine Science and Engineering 11, no. 2: 316. https://doi.org/10.3390/jmse11020316
APA StyleWang, H., Su, Q., Zhuang, Y., Wu, C., Tong, S., Guan, B., Zhao, Y., & Qiao, H. (2023). Effects of Iron Valence on the Growth, Photosynthesis, and Fatty Acid Composition of Phaeodactylum tricornutum. Journal of Marine Science and Engineering, 11(2), 316. https://doi.org/10.3390/jmse11020316