


Analysis of the Influence of Seasonal Water Column Dynamics on the Relationship between Marine Viruses and Microbial Food Web Components Using an Artificial Neural Network

 ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
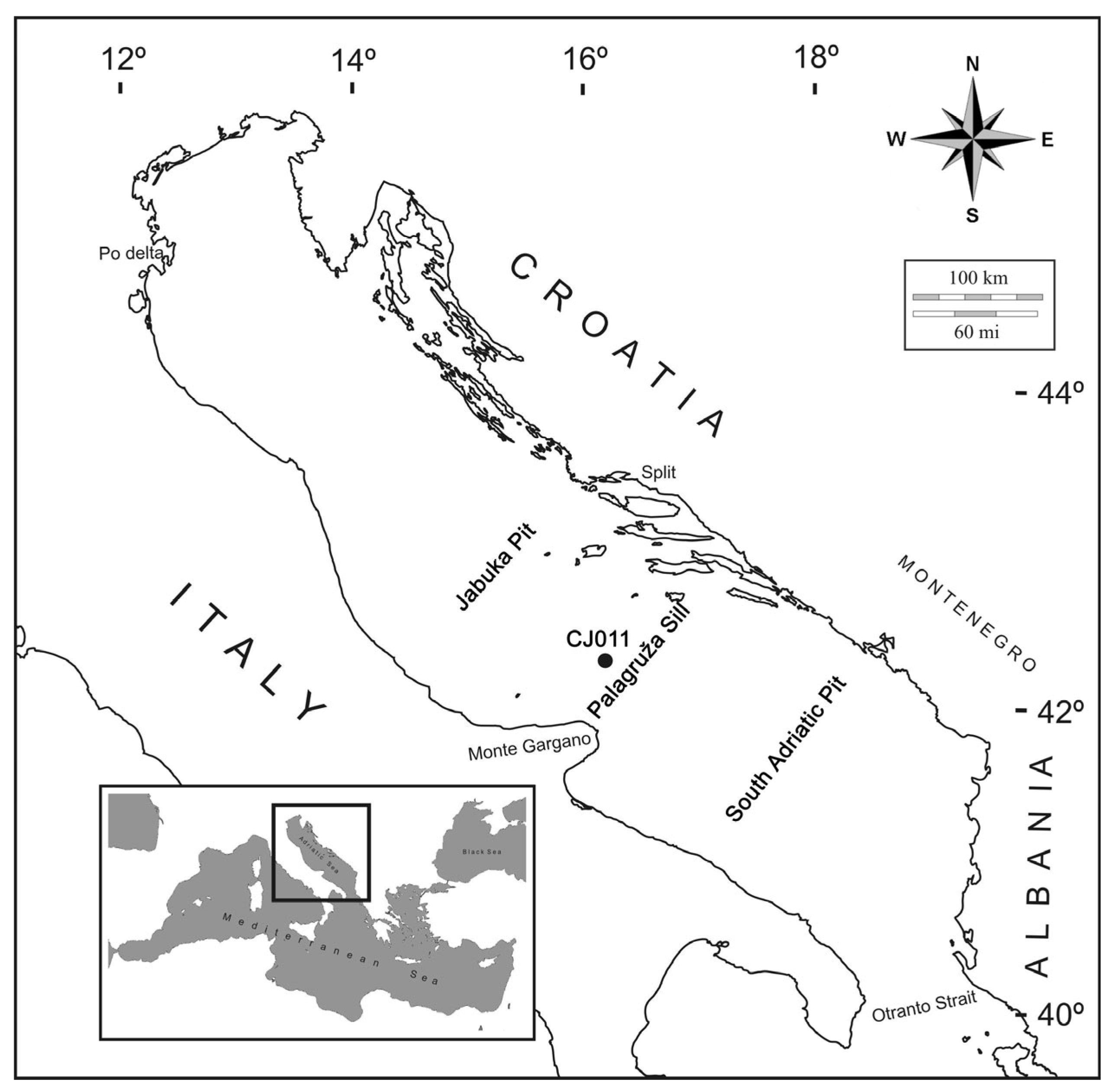
2.1. Study Sites and Sample Collection
2.2. Environmental Parameters
2.3. Marine Virus Counts
2.4. Bacterial, Synechococcus, Prochlorococcus, and Heterotrophic Nanoflagellate Counts
2.5. Bacterial Production
2.6. Characterization of the Studied Area according to Nutrient Status
2.7. Relative Importance of Bacterial Biomass Control Mechanisms
2.8. Statistical Analysis
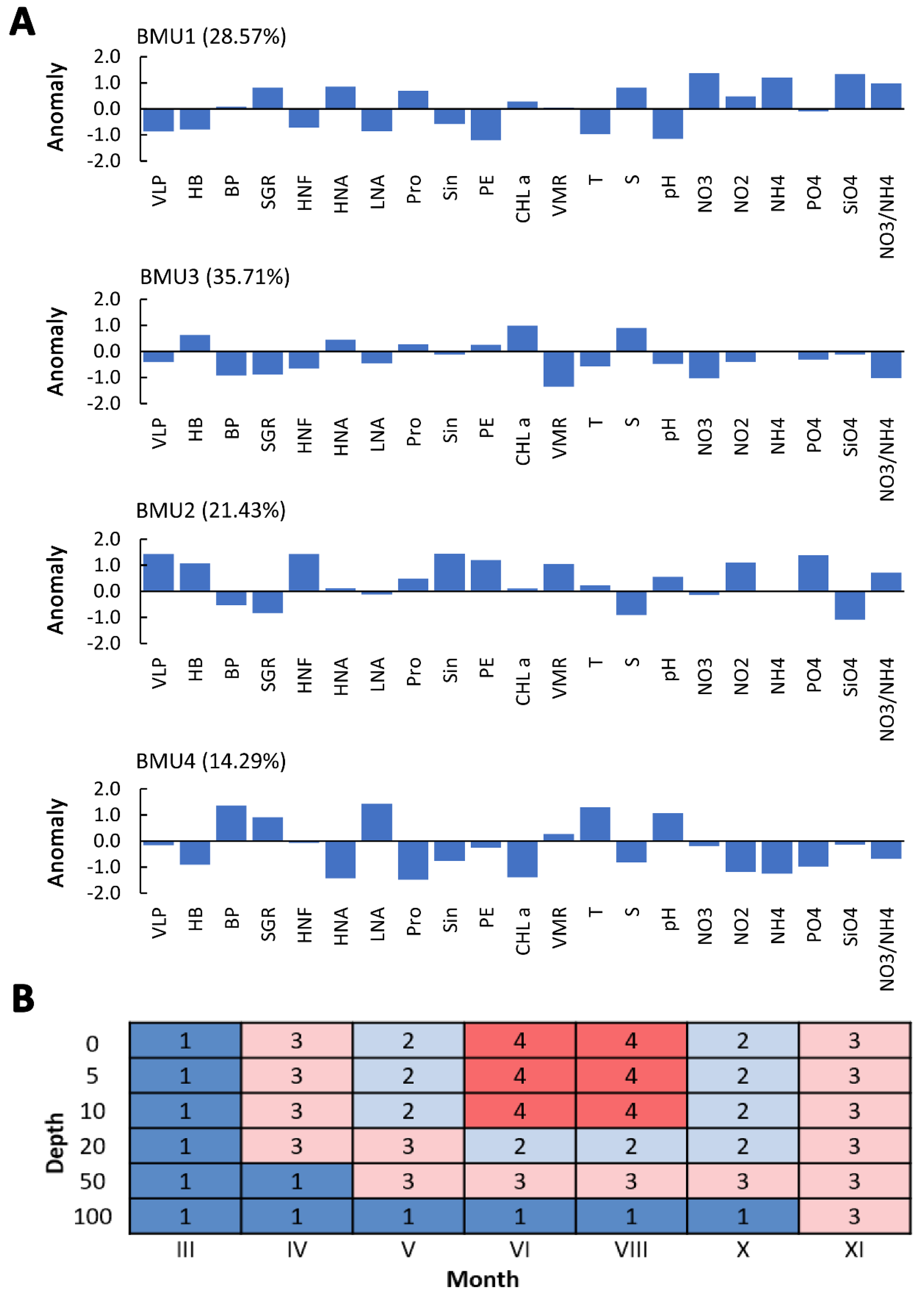
3. Results
3.1. Environmental Parameters
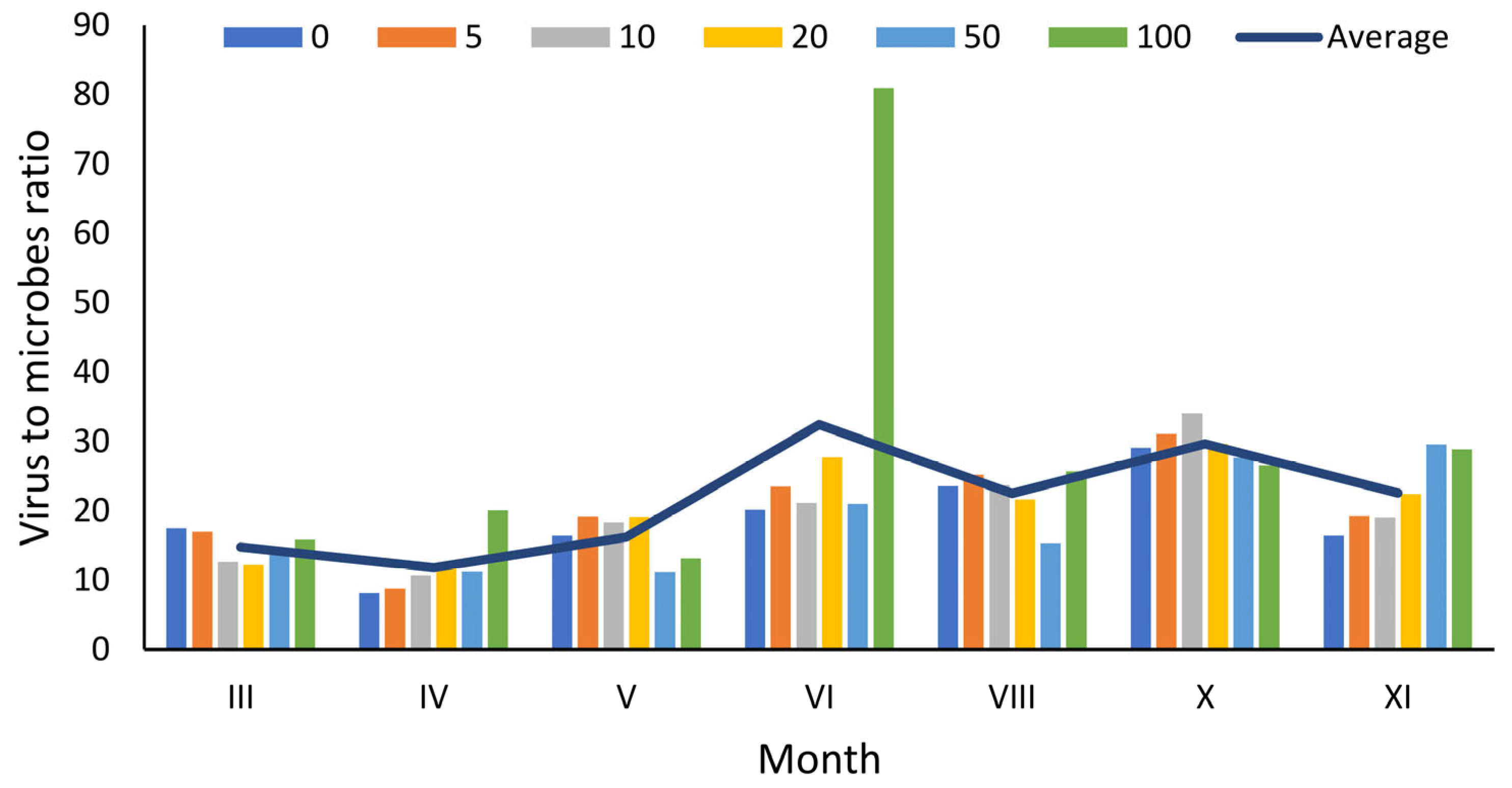
3.2. Distribution of Viruses
3.3. Abundance of Picoplankton and Nano Plankton Communities
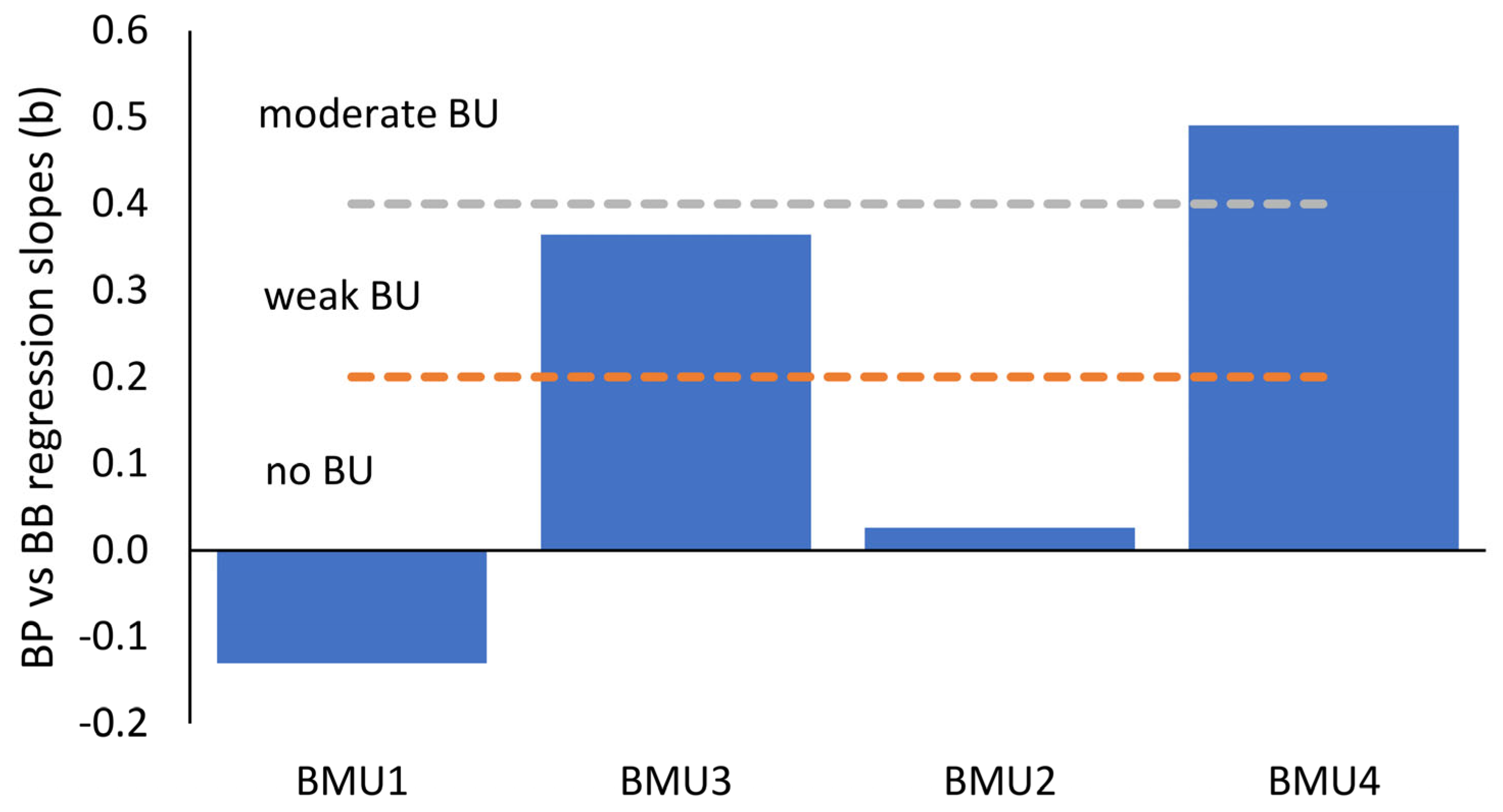
3.4. Top-Down vs. Bottom-Up Control of Heterotrophic Bacteria
3.5. Relationship between Viruses and the MFW Components
4. Discussion
4.1. Relation between Viruses and Microbial Picoplankton
4.2. Virus-to-Microbe Ratio
4.3. Control of Bacterial Populations in the Open Adriatic Sea
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergh, Ø.; Børsheim, K.Y.; Bratbak, G.; Heldal, M. High Abundance of Viruses Found in Aquatic Environments. Nature 1989, 340, 467–468. [Google Scholar] [CrossRef]
- Proctor, L.M.; Fuhrman, J.A. Viral Mortality of Marine Bacteria and Cyanobacteria. Nature 1990, 343, 60–62. [Google Scholar] [CrossRef]
- Suttle, C.A. Viruses in the Sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef]
- Parikka, K.J.; Le Romancer, M.; Wauters, N.; Jacquet, S. Deciphering the Virus-to-Prokaryote Ratio (VPR): Insights into Virus–Host Relationships in a Variety of Ecosystems. Biol. Rev. 2017, 92, 1081–1100. [Google Scholar] [CrossRef] [PubMed]
- Wigington, C.H.; Sonderegger, D.; Brussaard, C.P.D.; Buchan, A.; Finke, J.F.; Fuhrman, J.A.; Lennon, J.T.; Middelboe, M.; Suttle, C.A.; Stock, C.; et al. Re-Examination of the Relationship Between Marine Virus and Microbial Cell Abundances. Nat. Microbiol. 2016, 1, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Baudoux, A.C.; Veldhuis, M.J.W.; Witte, H.J.; Brussaard, C.P.D. Viruses as Mortality Agents of Picophytoplankton in the Deep Chlorophyll Maximum Layer During IRONAGES III. Limnol. Oceanogr. 2007, 52, 2519–2529. [Google Scholar] [CrossRef]
- Mojica, K.D.A.; Huisman, J.; Wilhelm, S.W.; Brussaard, C.P.D. Latitudinal Variation in Virus-Induced Mortality of Phytoplankton Across the North Atlantic Ocean. ISME J. 2016, 10, 500–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, A.Y.; Gong, G.C.; Chung, C.C.; Huang, Y.T. Different Impact of Nanoflagellate Grazing and Viral Lysis on Synechococcus spp. and Picoeukaryotic Mortality in Coastal Waters. Estuar. Coast. Shelf Sci. 2018, 209, 1–6. [Google Scholar] [CrossRef]
- Rohwer, F.; Thurber, R.V. Viruses Manipulate the Marine Environment. Nature 2009, 459, 207–212. [Google Scholar] [CrossRef]
- Weinbauer, M.G. Ecology of Prokaryotic Viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerman, A.E.; Howard-Varona, C.; Needham, D.M.; John, S.G.; Worden, A.Z.; Sullivan, M.B.; Waldbauer, J.R.; Coleman, M.L. Metabolic and Biogeochemical Consequences of Viral Infection in Aquatic Ecosystems. Nat. Rev. Microbiol. 2020, 18, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Poulton, A.J. Shunt or Shuttle. Nat. Geosci. 2021, 14, 180–181. [Google Scholar] [CrossRef]
- Allen, M.J.; Wilson, W.H. Aquatic Virus Diversity Accessed Through Omic Techniques: A Route Map to Function. Curr. Opin. Microbiol. 2008, 11, 226–232. [Google Scholar] [CrossRef]
- Danovaro, R.; Dell’Anno, A.; Corinaldesi, C.; Magagnini, M.; Noble, R.; Tamburini, C.; Weinbauer, M. Major Viral Impact on The Functioning of Benthic Deep-Sea Ecosystems. Nature 2008, 454, 1084–1087. [Google Scholar] [CrossRef]
- Kaneko, H.; Blanc-Mathieu, R.; Endo, H.; Chaffron, S.; Delmont, T.O.; Gaia, M.; Henry, N.; Hernández-Velázquez, R.; Nguyen, C.H.; Mamitsuka, H.; et al. Eukaryotic Virus Composition Can Predict the Efficiency of Carbon Export in the Global Ocean. iScience 2021, 24, 102002. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.B.; Weitz, J.S.; Wilhelm, S. Viral Ecology Comes of Age. Environ. Microbiol. Rep. 2017, 9, 3. [Google Scholar] [CrossRef]
- Riemann, L.; Holmfeldt, K.; Titelman, J. Importance of Viral Lysis and Dissolved DNA For Bacterioplankton Activity in A P-Limited Estuary, Northern Baltic Sea. Microb. Ecol. 2009, 57, 286–294. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Bonilla-Findji, O.; Chan, A.M.; Dolan, J.R.; Short, S.M.; Imek, K.; Wilhelm, S.W.; Suttle, C.A. Synechococcus Growth in the Ocean May Depend on the Lysis of Heterotrophic Bacteria. J. Plankton Res. 2011, 33, 1465–1476. [Google Scholar] [CrossRef]
- Jover, L.F.; Effler, T.C.; Buchan, A.; Wilhelm, S.W.; Weitz, J.S. The Elemental Composition of Virus Particles: Implications for Marine Biogeochemical Cycles. Nat. Rev. Microbiol. 2014, 12, 519–528. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Suttle, C.A. Grazing by Marine Nanoflagellates on Viruses and Virus-Sized Particles: Ingestion and Digestion. Mar. Ecol. Prog. Ser. 1993, 94, 1–10. [Google Scholar] [CrossRef]
- Bongiorni, L.; Magagnini, M.; Armeni, M.; Noble, R.; Danovaro, R. Viral Production, Decay Rates, and Life Strategies Along a Trophic Gradient in the North Adriatic Sea. Appl. Environ. Microbiol. 2005, 71, 6644–6650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, J.E.; Steenhuis, P.; de Moraes, K.R.; van der Meer, J.; Thieltges, D.W.; Brussaard, C.P.D. Marine Virus Predation by Non-Host Organisms. Sci. Rep. 2020, 10, 5221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thingstad, T.F.; Skjoldal, E.F.; Bohne, R.A. Phosphorus Cycling and Algal-Bacterial Competition in Sandsfjord, Western Norway. Mar. Ecol. Prog. Ser. 1993, 99, 239–259. [Google Scholar] [CrossRef]
- Jacquet, S.; Miki, T.; Noble, R.; Peduzzi, P.; Wilhelm, S. Viruses in Aquatic Ecosystems: Important Advancements of the Last 20 Years and Prospects for the Future in the Field of Microbial Oceanography and Limnology. Adv. Oceanogr. Limnol. 2010, 1, 97–141. [Google Scholar] [CrossRef]
- Ordulj, M.; Krstulović, N.; Šantić, D.; Jozić, S.; Šolić, M. Distribution of Marine Viruses in the Central and South Adriatic Sea. Mediterr. Mar. Sci. 2015, 16, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Suttle, C.A. Marine Viruses–Major Players in the Global Ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Winter, C.; Bouvier, T.; Weinbauer, M.G.; Thingstad, T.F. Trade-Offs Between Competition and Defense Specialists Among Unicellular Planktonic Organisms: The “Killing the Winner” Hypothesis Revisited. Microbiol. Mol. Biol. Rev. 2010, 74, 42–57. [Google Scholar] [CrossRef] [Green Version]
- Weinbauer, M.G.; Rassoulzadegan, F. Are Viruses Driving Microbial Diversification and Diversity? Environ. Microbiol. 2004, 6, 1–11. [Google Scholar] [CrossRef]
- Šolić, M.; Krstulović, N.; Kušpilić, G.; Ninčević Gladan, Ž.; Bojanić, N.; Šestanović, S.; Šantić, D.; Ordulj, M.; Ninčević Gladan, T.; Bojanić, N.; et al. Changes in Microbial Food Web Structure in Response to Changed Environmental Trophic Status: A Case Study of the Vranjic Basin (Adriatic Sea). Mar. Environ. Res. 2010, 70, 239–249. [Google Scholar] [CrossRef]
- Šantić, D.; Krstulović, N.; Šolić, M.; Ordulj, M.; Kušpilić, G. Dynamics of Prokaryotic Picoplankton Community in the Central and Southern Adriatic Sea (Croatia). Helgol. Mar. Res. 2013, 67, 471–481. [Google Scholar] [CrossRef] [Green Version]
- Šolić, M.; Grbec, B.; Matić, F.; Šantić, D.; Šestanović, S.; Ninčević Gladan, Ž.; Bojanić, N.; Ordulj, M.; Jozić, S.; Vrdoljak, A. Spatio-Temporal Reproducibility of the Microbial Food Web Structure Associated with the Change in Temperature: Long-Term Observations in the Adriatic Sea. Prog. Oceanogr. 2018, 161, 87–101. [Google Scholar] [CrossRef]
- Šolić, M.; Šantić, D.; Šestanović, S.; Bojanić, N.; Grbec, B.; Jozić, S.; Vrdoljak, A.; Ordulj, M.; Matić, F.; Kušpilić, G.; et al. Impact of Water Column Stability Dynamics on the Succession of Plankton Food Web Types in the Offshore Area of the Adriatic Sea. J. Sea Res. 2020, 158, 101860. [Google Scholar] [CrossRef]
- Šolić, M.; Šantić, D.; Šestanović, S.; Kušpilić, G.; Matić, F.; Vrdoljak Tomaš, A.; Jozić, S.; Bojanić, N.; Ninčević Gladan, Ž. Changing Ecological Conditions in the Marine Environment Generate Different Microbial Food Web Structures in a Repeatable Manner. Front. Mar. Sci. 2022, 8, 2104. [Google Scholar] [CrossRef]
- Malfatti, F.; Turk, V.; Tinta, T.; Mozetič, P.; Manganelli, M.; Samo, T.J.; Ugalde, J.A.; Kovač, N.; Stefanelli, M.; Antonioli, M.; et al. Microbial Mechanisms Coupling Carbon and Phosphorus Cycles in Phosphorus-Limited Northern Adriatic Sea. Sci. Total Environ. 2014, 470–471, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Šolić, M.; Krstulović, N.; Šantić, D.; Šestanović, S.; Ordulj, M.; Bojanić, N.; Kušpilić, G. Structure of Microbial Communities in Phosphorus-Limited Estuaries Along the Eastern Adriatic Coast. J. Mar. Biol. Assoc. UK 2015, 95, 1565–1578. [Google Scholar] [CrossRef]
- Alonso, M.C.; Jimenez-Gomez, F.; Rodriguez, J.; Borrego, J.J. Distribution of Virus-Like Particles in an Oligotrophic Marine Environment (Alboran Sea, Western Mediterranean). Microb. Ecol. 2001, 42, 407–415. [Google Scholar] [CrossRef]
- Magagnini, M.; Corinaldesi, C.; Monticelli, L.S.; De Domenico, E.; Danovaro, R. Viral Abundance and Distribution in Mesopelagic and Bathypelagic Waters of the Mediterranean Sea. Deep. Res. Part I Oceanogr. Res. Pap. 2007, 54, 1209–1220. [Google Scholar] [CrossRef]
- Boras, J.A.; Sala, M.M.; Vázquez-Domínguez, E.; Weinbauer, M.G.; Vaqué, D. Annual Changes of Bacterial Mortality due to Viruses and Protists in an Oligotrophic Coastal Environment (NW Mediterranean). Environ. Microbiol. 2009, 11, 1181–1193. [Google Scholar] [CrossRef]
- Winter, C.; Kerros, M.E.; Weinbauer, M.G. Seasonal and Depth-Related Dynamics of Prokaryotes and Viruses in Surface and Deep Waters of the Northwestern Mediterranean Sea. Deep. Res. Part I Oceanogr. Res. Pap. 2009, 56, 1972–1982. [Google Scholar] [CrossRef]
- Magiopoulos, I.; Pitta, P. Viruses in a Deep Oligotrophic Sea: Seasonal Distribution of Marine Viruses in the Epi-, Meso- and Bathypelagic Waters of the Eastern Mediterranean Sea. Deep. Res. Part I Oceanogr. Res. Pap. 2012, 66, 1–10. [Google Scholar] [CrossRef]
- Johnstone, C.; Salles, S.; Mercado, J.M.; Cortés, D.; Yebra, L.; Gómez-Jakobsen, F.; Sánchez, A.; Alonso, A.; Valcárcel-Pérez, N. Abundance of Virus-Like Particles (Vlps) and Microbial Plankton Community Composition in a Mediterranean Sea Coastal Area. Aquat. Microb. Ecol. 2018, 81, 137–148. [Google Scholar] [CrossRef]
- Corinaldesi, C.; Crevatin, E.; Del Negro, P.; Marini, M.; Russo, A.; Fonda-Umani, S.; Danovaro, R. Large-Scale Spatial Distribution of Virioplankton in the Adriatic Sea: Testing the Trophic State Control Hypothesis. Appl. Environ. Microbiol. 2003, 69, 2664–2673. [Google Scholar] [CrossRef] [Green Version]
- Šantić, D.; Kovačević, V.; Bensi, M.; Giani, M.; Tomaš, A.V.A.V.; Ordulj, M.; Santinelli, C.; Šestanović, S.; Šolić, M.; Grbec, B. Picoplankton Distribution and Activity in the Deep Waters of the Southern Adriatic Sea. Water 2019, 11, 1655. [Google Scholar] [CrossRef] [Green Version]
- Grasshoff, K.; Ehrhardt, M.; Kremling, K. Methods of Sea Water Analysis. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1983, 70, 302–303. [Google Scholar] [CrossRef]
- Parsons, R.J.; Breitbart, M.; Lomas, M.W.; Carlson, C.A. Ocean Time-Series Reveals Recurring Seasonal Patterns of Virioplankton Dynamics in the Northwestern Sargasso Sea. ISME J. 2012, 6, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1972; Volume 167. [Google Scholar]
- Noble, R.T.; Fuhrman, J.A. Use of SYBR Green I for Rapid Epifluorescence Counts of Marine Viruses and Bacteria. Aquat. Microb. Ecol. 1998, 14, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Zubkov, M.V.; Burkill, P.H.; Topping, J.N. Flow Cytometric Enumeration of DNA-Stained Oceanic Planktonic Protists. J. Plankton Res. 2006, 29, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Christaki, U.; Van Wambeke, F.; Lefevre, D.; Lagaria, A.; Prieur, L.; Pujo-Pay, M.; Grattepanche, J.D.; Colombet, J.; Psarra, S.; Dolan, J.R.; et al. Microbial Food Webs and Metabolic State Across Oligotrophic Waters of the Mediterranean Sea During Summer. Biogeosciences 2011, 8, 1839–1852. [Google Scholar] [CrossRef] [Green Version]
- Fuhrman, J.A.; Azam, F. Thymidine Incorporation as a Measure of Heterotrophic Bacterioplankton Production in Marine Surface Waters: Evaluation and Field Results. Mar. Biol. 1982, 66, 109–120. [Google Scholar] [CrossRef]
- Riemann, B.; Bjørnsen, P.K.; Newell, S.; Fallon, R. Calculation of Cell Production of Coastal Marine Bacteria Based on Measured Incorporation of [3H]thymidine 1, 2. Limnol. Oceanogr. 1987, 32, 471–476. [Google Scholar] [CrossRef]
- Rhee, G.Y. Phosphate uptake under nitrate limitation by Scenedesmus sp. and its ecological implications. J. Phycol. 1974, 9, 495–506. [Google Scholar] [CrossRef]
- Perry, M.J.; Eppley, R.W. Phosphate Uptake by Phytoplankton in the Central North Pacific Ocean. Deep Sea Res. Part A Oceanogr. Res. Pap. 1981, 28, 39–49. [Google Scholar] [CrossRef]
- Brzezinski, M.A. The Si:C:N Ratio of Marine Diatoms: Interspecific Variability and the Effect of Some Environmental Variables. J. Phycol. 1985, 21, 347–357. [Google Scholar] [CrossRef]
- Goldman, J.C.; Glibert, P.M. Kinetics of Inorganic Nitrogen Uptake by Phytoplankton. In Nitrogen in the Marine Environment; Elsevier: Amsterdam, The Netherlands, 1983; pp. 233–274. [Google Scholar]
- Dortch, Q.; Whitledge, T.E. Does Nitrogen or Silicon Limit Phytoplankton Production in the Mississippi River Plume and Nearby Regions? Cont. Shelf Res. 1992, 12, 1293–1309. [Google Scholar] [CrossRef]
- Justić, D.; Rabalais, N.N.; Turner, R.E.; Dortch, Q. Changes in Nutrient Structure of River-Dominated Coastal Waters: Stoichiometric Nutrient Balance and Its Consequences. Estuar. Coast. Shelf Sci. 1995, 40, 339–356. [Google Scholar] [CrossRef]
- Billen, G.; Servais, P.; Becquevort, S. Dynamics of Bacterioplankton in Oligotrophic and Eutrophic Aquatic Environments: Bottom-Up or Top-Down Control? Hydrobiologia 1990, 207, 37–42. [Google Scholar] [CrossRef]
- Ducklow, H.W. Factors Regulating Bottom-Up Control of Bacteria Biomass in Open Ocean Plankton Communities. Arch. Hydrobiol. Beih. Ergebn. Limnol. 1992, 37, 207–217. [Google Scholar]
- Šolić, M.; Šantić, D.; Šestanović, S.; Bojanić, N.; Jozić, S.; Ordulj, M.; Tomaš, A.V.; Kušpilić, G. Changes in the Trophic Pathways within the Microbial Food Web in the Global Warming Scenario: An Experimental Study in the Adriatic Sea. Microorganisms 2020, 8, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Fuhrman, J.A. Relationships between Biovolume and Biomass of Naturally Derived Marine Bacterioplankton. Appl. Environ. Microbiol. 1987, 53, 1298–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchman, D.L.; Keil, R.G.; Simon, M. Biomass and Production of Heterotrophic Bacterioplankton in the Oceanic Subarctic Pacific Samples for Bacterial Abundance and Incorporation Rates Were Taken from CTD Niskin Bottles. These Bottles All Had Silicone Tubing During the Cruises Reported Here. Deep. Res. Part 1. Oceanogr. Res. Pap. 1993, 40, 967–988. [Google Scholar] [CrossRef]
- Kohonen, T. Self-Organized Formation of Topologically Correct Feature Maps. Biol. Cybern. 1982, 43, 59–69. [Google Scholar] [CrossRef]
- Martinetz, T.; Schulten, K. A “Neural Gas” Network Learns Topologies. In Artificial Neural Networks; Elsevier: Amsterdam, The Netherlands, 1991; pp. 397–402. [Google Scholar]
- Martinetz, T.; Berkovich, S.G.; Schulten, K.J. “Neural-gas” Network for Vector Quantization and its Application to Time-Series Prediction. IEEE Trans. Neural Netw. 1993, 4, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Fonda, U.S.; Malisana, E.; Focaracci, F.; Magagnini, M.; Corinaldesi, C.; Danovaro, R. Disentangling the Effect of Viruses and Nanoflagellates on Prokaryotes in Bathypelagic Waters of the Mediterranean Sea. Mar. Ecol. Prog. Ser. 2010, 418, 73–85. [Google Scholar] [CrossRef]
- Xie, L.; Wei, W.; Cai, L.; Chen, X.; Huang, Y.; Jiao, N.; Zhang, R.; Luo, Y.-W. A Global Viral Oceanography Database (gVOD). Earth Syst. Sci. Data Discuss. 2020, 13, 1251–1271. [Google Scholar] [CrossRef]
- Cochlan, W.P.; Wikner, J.; Steward, G.F.; Smith, D.C. Environments. Mar. Ecol. -Prog. Ser. 1993, 92, 77–87. [Google Scholar] [CrossRef]
- Liang, Y.; Li, L.; Luo, T.; Zhang, Y.; Zhang, R.; Jiao, N. Horizontal and Vertical Distribution of Marine Virioplankton: A basin Scale Investigation Based on a Global Cruise. PLoS ONE 2014, 9, e111634. [Google Scholar] [CrossRef]
- Ordulj, M.; Krstulović, N.; Šantić, D.; Jozić, S.; Šolić, M. Viral Dynamics in two Trophically Different Areas in the Central Adriatic Sea. Helgol. Mar. Res. 2017, 71, 22. [Google Scholar] [CrossRef] [Green Version]
- Brum, J.R.; Hurwitz, B.L.; Schofield, O.; Ducklow, H.W.; Sullivan, M.B. Seasonal Time Bombs: Dominant Temperate Viruses Affect Southern Ocean Microbial Dynamics. ISME J. 2016, 10, 437–449. [Google Scholar] [CrossRef]
- Sotomayor-Garcia, A.; Sala, M.M.; Ferrera, I.; Estrada, M.; Vázquez-Domínguez, E.; Emelianov, M.; Cortés, P.; Marrasé, C.; Ortega-Retuerta, E.; Nunes, S.; et al. Assessing Viral Abundance and Community Composition in Four Contrasting Regions of the Southern Ocean. Life 2020, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- De Corte, D.; Sintes, E.; Yokokawa, T.; Reinthaler, T.; Herndl, G.J. Links Between Viruses and Prokaryotes Throughout the Water Column Along a North Atlantic Latitudinal Transect. ISME J. 2012, 6, 1566–1577. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhang, Y.; Zhang, Y.; Luo, T.; Rivkin, R.B.; Jiao, N. Distributions and Relationships of Virio- and Picoplankton in The Epi-, Meso- and Bathypelagic Zones of the Western Pacific Ocean. FEMS Microbiol. Ecol. 2017, 93, fiw238. [Google Scholar] [CrossRef]
- Sabbagh, E.I.; Huete-Stauffer, T.M.; Calleja, M.L.L.; Silva, L.; Viegas, M.; Morán, X.A.G. Weekly Variations of Viruses and Heterotrophic Nanoflagellates and Their Potential Impact on Bacterioplankton in Shallow Waters of the Central Red Sea. FEMS Microbiol. Ecol. 2021, 96, fiaa033. [Google Scholar] [CrossRef] [PubMed]
- Šantić, D.; Piwosz, K.; Matić, F.; Vrdoljak Tomaš, A.; Arapov, J.; Dean, J.L.; Šolić, M.; Koblížek, M.; Kušpilić, G.; Šestanović, S. Artificial Neural Network Analysis of Microbial Diversity in the Central and Southern Adriatic Sea. Sci. Rep. 2021, 11, 11186. [Google Scholar] [CrossRef] [PubMed]
- Winter, C.; Smit, A.; Szoeke-Dénes, T.; Herndl, G.J.; Weinbauer, M.G. Modelling Viral Impact on Bacterioplankton in the North Sea Using Artificial Neural Networks. Environ. Microbiol. 2005, 7, 881–893. [Google Scholar] [CrossRef]
- Pereira, G.C.; Oliveira, M.M.F.; Ebecken, N.F.F. Genetic Optimization of Artificial Neural Networks to Forecast Virioplankton Abundance from Cytometric Data. J. Intell. Learn. Syst. Appl. 2013, 05, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Hurwitz, B.L.; Sullivan, M.B. The Pacific Ocean Virome (POV): A Marine Viral Metagenomic Dataset and Associated Protein Clusters for Quantitative Viral Ecology. PLoS ONE 2013, 8, e57355. [Google Scholar] [CrossRef]
- Breitbart, M.; Thompson, L.R.; Suttle, C.A.; Sullivan, M.B. Exploring the Vast Diversity of Marine Viruses. Oceanography 2007, 20, 135–139. [Google Scholar] [CrossRef]
- Wommack, K.E.; Colwell, R.R. Virioplankton: Viruses in Aquatic Ecosystems. Microbiol. Mol. Biol. Rev. 2000, 64, 69–114. [Google Scholar] [CrossRef] [Green Version]
- Šantić, D.; Šestanović, S.; Šolić, M.; Krstulović, N.; Kušpilić, G.; Ordulj, M.; Ninčević Gladan, Ž. Dynamics of the Picoplankton Community from Coastal Waters to the Open Sea in the Central Adriatic. Mediterr. Mar. Sci. 2014, 15, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Šantić, D.; Krstulović, N.; Šolić, M.; Kušpilić, G. HNA and LNA Bacteria in Relation to the Activity of Heterotrophic Bacteria. Acta Adriat. 2012, 53, 25–39. [Google Scholar]
- Trombetta, T.; Vidussi, F.; Roques, C.; Scotti, M.; Mostajir, B. Marine Microbial Food Web Networks During Phytoplankton Bloom and Non-bloom Periods: Warming Favors Smaller Organism Interactions and Intensifies Trophic Cascade. Front. Microbiol. 2020, 11, 502336. [Google Scholar] [CrossRef] [PubMed]
- Thingstad, T.F. Elements of a Theory for the Mechanisms Controlling Abundance, Diversity, and Biogeochemical Role of Lytic Bacterial Viruses in Aquatic Systems. Limnol. Oceanogr. 2000, 45, 1320–1328. [Google Scholar] [CrossRef]
- Touchon, M.; Bernheim, A.; Rocha, E.P.C. Genetic and Life-History Traits Associated with the Distribution of Prophages in Bacteria. ISME J. 2016, 10, 2744–2754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Ma, R.; Yang, Y.; Jiao, N.; Zhang, R. Viral Regulation on Bacterial Community Impacted by Lysis-Lysogeny Switch: A Microcosm Experiment in Eutrophic Coastal Waters. Front. Microbiol. 2019, 10, 1763. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yokokawa, T.; Motegi, C.; Nagata, T. Large-Scale Distribution of Viruses in Deep Waters of the Pacific and Southern Oceans. Aquat. Microb. Ecol. 2014, 71, 193–202. [Google Scholar] [CrossRef] [Green Version]
- De Corte, D.; Sintes, E.; Yokokawa, T.; Lekunberri, I.; Herndl, G.J. Large-Scale Distribution of Microbial and Viral Populations in the South Atlantic Ocean. Environ. Microbiol. Rep. 2016, 8, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Johannessen, T.V.; Larsen, A.; Bratbak, G.; Pagarete, A.; Edvardsen, B.; Egge, E.D.; Sandaa, R.A. Seasonal Dynamics of Haptophytes and Dsdna Algal Viruses Suggest Complex Virus-Host Relationship. Viruses 2017, 9, 84. [Google Scholar] [CrossRef]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera D’Alcala, M.; Vaque, D.; Zingone, A. Plankton in the Open Mediterranean Sea: A Review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef] [Green Version]
- Parada, V.; Sintes, E.; Van Aken, H.M.; Weinbauer, M.G.; Herndl, G.J. Viral Abundance, Decay, and Diversity in the Meso- and Bathypelagic Waters of the North Atlantic. Appl. Environ. Microbiol. 2007, 73, 4429–4438. [Google Scholar] [CrossRef] [Green Version]
- Hurwitz, B.L.; Brum, J.R.; Sullivan, M.B. Depth-stratified functional and taxonomic niche specialization in the “core” and “flexible” Pacific Ocean Virome. ISME J. 2015, 9, 472–484. [Google Scholar] [CrossRef] [Green Version]
- Winter, C.; Köstner, N.; Kruspe, C.P.; Urban, D.; Muck, S.; Reinthaler, T.; Herndl, G.J. Mixing Alters the Lytic Activity of Viruses in the Dark Ocean. Ecology 2018, 99, 700–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Corte, D.; Sintes, E.; Winter, C.; Yokokawa, T.; Reinthaler, T.; Herndl, G.J. Links Between Viral and Prokaryotic Communities Throughout the Water Column in the (Sub)Tropical Atlantic Ocean. ISME J. 2010, 4, 1431–1442. [Google Scholar] [CrossRef] [PubMed]
- Šolić, M.; Krstulović, N.; Vilibić, I.; Bojanić, N.; Kušpilić, G.; Šestanović, S.; Šantić, D.; Ordulj, M. Variability in the Bottom-Up and Top-Down Controls of Bacteria on Trophic and Temporal Scales in the Middle Adriatic Sea. Aquat. Microb. Ecol. 2010, 58, 15–29. [Google Scholar] [CrossRef] [Green Version]
- Weinbauer, M.G.; Christaki, U.; Nedoma, J.; Šimek, K. Comparing the Effects of Resource Enrichment and Grazing on Viral Production in a Meso-Eutrophic Reservoir. Aquat. Microb. Ecol. 2003, 31, 137–144. [Google Scholar] [CrossRef]
- Middelboe, M.; Riemann, L.; Steward, G.F.; Hansen, V.; Nybroe, O. Virus-induced transfer of organic carbon between marine bacteria in a model community. Aquat. Microb. Ecol. 2003, 33, a033P001. [Google Scholar] [CrossRef] [Green Version]
- Ory, P.; Hartmann, H.J.; Jude, F.; Dupuy, C.; Del Amo, Y.; Catala, P.; Mornet, F.; Huet, V.; Jan, B.; Vincent, D.; et al. Pelagic Food Web Patterns: Do They Modulate Virus and Nanoflagellate Effects on Picoplankton During the Phytoplankton Spring Bloom? Environ. Microbiol. 2010, 12, 2755–2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuhrman, J.A. Marine Viruses and Their Biogeochemical and Ecological Effects. Nature 1999, 399, 541–548. [Google Scholar] [CrossRef]
- Brussaard, C.P.D.; Wilhelm, S.W.; Thingstad, F.; Weinbauer, M.G.; Bratbak, G.; Heldal, M.; Kimmance, S.A.; Middelboe, M.; Nagasaki, K.; Paul, J.H.; et al. Global-Scale Processes with a Nanoscale Drive: The Role of Marine Viruses. ISME J. 2008, 2, 575–578. [Google Scholar] [CrossRef] [Green Version]
- Gasol, J.M. A Framework for the Assessment of Top-Down vs. Bottom-Up Control of Heterotrophic Nanoflagellate Abundance. Mar. Ecol. Prog. Ser. 1994, 113, 291–300. [Google Scholar] [CrossRef]
- Šimek, K.; Pernthaler, J.; Weinbauer, M.G.; Hornák, K.; Dolan, J.R.; Nedoma, J.; Mašín, M.; Amann, R. Changes in Bacterial Community Composition and Dynamics and Viral Mortality Rates Associated with Enhanced Flagellate Grazing in a Mesoeutrophic Reservoir. Appl. Environ. Microbiol. 2001, 67, 2723–2733. [Google Scholar] [CrossRef] [Green Version]
- Ho, P.; Gong, G.; Mukhanov, V.; Zhu, Z.; Tsai, A. Annual Cycle of the Synechococcus spp. and Picoeukaryotic Growth and Loss Rates in a Subtropical Coastal Ecosystem. Diversity 2022, 14, 49. [Google Scholar] [CrossRef]
- Shiah, F.-K.; Lai, C.-C.; Chen, T.-Y.; Ko, C.-Y.; Tai, J.-H.; Chang, C.-W. Viral Shunt in Tropical Oligotrophic Ocean. Sci. Adv. 2022, 8, eabo2829. [Google Scholar] [CrossRef]
- Vaqué, D.; Gasol, J.M.; Marrasé, C. Grazing rates on bacteria: The Significance of Methodology and Ecological Factors. Mar. Ecol. Prog. Ser. 1994, 109, 263–274. [Google Scholar] [CrossRef]
- Chen, X.; Weinbauer, M.G.; Jiao, N.; Zhang, R. Revisiting Marine Lytic and Lysogenic Virus-Host Interactions: Kill-the-Winner and Piggyback-the-Winner. Sci. Bull. 2021, 66, 871–874. [Google Scholar] [CrossRef] [PubMed]
- Weinbauer, M.G.; Kerros, M.E.; Motegi, C.; Wilhartitz, I.C.; Rassoulzadegan, F.; Torreton, J.P.; Mari, X. Bacterial Community Composition and Potential Controlling Mechanisms Along a Trophic Gradient in a Barrier Reef System. Aquat. Microb. Ecol. 2010, 60, 15–28. [Google Scholar] [CrossRef]
- Christaki, U.; Giannakourou, A.; Van Wambeke, F.; Grégori, G. Nanoflagellate Predation on Auto- And Heterotrophic Picoplankton in the Oligotrophic Mediterranean Sea. J. Plankton Res. 2001, 23, 1297–1310. [Google Scholar] [CrossRef] [Green Version]
- Frias-Lopez, J.; Thompson, A.; Waldbauer, J.; Chisholm, S.W. Use of Stable Isotope-Labelled Cells to Identify Active Grazers of Picocyanobacteria in Ocean Surface Waters. Environ. Microbiol. 2009, 11, 512–525. [Google Scholar] [CrossRef] [Green Version]
- Bouvy, M.; Bettarel, Y.; Bouvier, C.; Domaizon, I.; Jacquet, S.; Le Floc’h, E.; Montanié, H.; Mostajir, B.; Sime-Ngando, T.; Torréton, J.P.; et al. Trophic Interactions between Viruses, Bacteria and Nanoflagellates Under Various Nutrient Conditions and Simulated Climate Change. Environ. Microbiol. 2011, 13, 1842–1857. [Google Scholar] [CrossRef] [PubMed]
- Parada, V.; Herndl, G.J.; Weinbauer, M.G. Viral Burst Size of Heterotrophic Prokaryotes in Aquatic Systems. J. Mar. Biol. Assoc. UK 2006, 86, 613–621. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Fuks, D.; Peduzzi, P. Distribution of Viruses and Dissolved DNA Along a Coastal Trophic Gradient in the Northern Adriatic Sea. Appl. Environ. Microbiol. 1993, 59, 4074–4082. [Google Scholar] [CrossRef] [Green Version]
- Šolić, M.; Šantić, D.; Šestanović, S.; Bojanić, N.; Ordulj, M.; Jozić, S.; Vrdoljak, A. The effect of Temperature Increase on Microbial Carbon Fluxes in the Adriatic Sea: An Experimental Approach. FEMS Microbiol. Ecol. 2018, 94, fiy169. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CJ011 | BMU1 | BMU2 | BMU3 | BMU4 | |
|---|---|---|---|---|---|
| VLP (×106 VLP mL−1) | 6.7 ± 2.28 | 5.14 ± 0.68 | 9.89 ± 1.99 | 6.09 ± 1.76 | 6.59 ± 1.13 |
| HB (×106 cells mL−1) | 0.34 ± 0.09 | 0.29 ± 0.1 | 0.39 ± 0.02 | 0.37 ± 0.1 | 0.29 ± 0.06 |
| BP (×104 cells mL−1) | 0.27 ± 0.18 | 0.28 ± 0.3 | 0.26 ± 0.07 | 0.25 ± 0.12 | 0.32 ± 0.09 |
| HNF (×103 cells mL−1) | 2.04 ± 1.27 | 1.55 ± 0.96 | 3.33 ± 1.76 | 1.59 ± 0.75 | 2.08 ± 0.78 |
| HNA (%) | 49.9 ± 6.21 | 51.53 ± 4.49 | 49.62 ± 9.65 | 50.47 ± 5.12 | 45.6 ± 24.24 |
| LNA (%) | 50.1 ± 6.21 | 48.48 ± 4.49 | 50.38 ± 9.65 | 49.53 ± 5.12 | 54.4 ± 4.44 |
| PRO (×103 cells mL−1) | 1.95 ± 1.71 | 2.36 ± 1.04 | 2.19 ± 2.45 | 2.02 ± 1.81 | 0.61 ± 0.32 |
| SYN (×103 cells mL−1) | 2.66 ± 3.39 | 1.65 ± 1.39 | 5.21 ± 6.19 | 2.46 ± 2.07 | 1.34 ± 0.43 |
| PE (×103 cells mL−1) | 1.23 ± 0.79 | 0.74 ± 0.51 | 1.74 ± 1.16 | 1.34 ± 0.67 | 1.14 ± 0.27 |
| CHL a (mg m−3) | 0.27 ± 0.26 | 0.27 ± 0.19 | 0.25 ± 0.36 | 0.37 ± 0.25 | 0.05 ± 0.06 |
| SGR (day−1) | 0.56 ± 0.32 | 0.62 ± 0.59 | 0.59 ± 0.40 | 0.50 ± 0.22 | 0.54 ± 0.39 |
| VMR | 21.42 ± 11.50 | 23 ± 21.11 | 25.7 ± 6.11 | 18.29 ± 7.34 | 23.1 ± 1.86 |
| T (°C) | 16.93 ± 3.36 | 13.85 ± 0.57 | 18.97 ± 0.88 | 15.52 ± 0.56 | 23.55 ± 1 |
| S | 38.62 ± 0.34 | 38.81 ± 0.07 | 38.25 ± 0.34 | 38.84 ± 0.07 | 38.28 ± 0.38 |
| O2 (mL/L) | 5.65 ± 0.31 | 5.7 ± 0.2 | 5.66 ± 0.29 | 5.61 ± 0.33 | 5.61 ± 0.47 |
| pH | 8.20 ± 0.03 | 8.18 ± 0.02 | 8.22 ± 0.02 | 8.2 ± 0.03 | 8.24 ± 0.01 |
| NO3− (μmol L−1) | 1.01 ± 0.99 | 1.6 ± 1.36 | 0.96 ± 0.68 | 0.59 ± 0.71 | 0.94 ± 0.77 |
| NO2− (μmol L−1) | 0.13 ± 0.14 | 0.15 ± 0.14 | 0.19 ± 0.23 | 0.11 ± 0.06 | 0.06 ± 0.04 |
| NH4+ (μmol L−1) | 0.74 ± 0.54 | 0.93 ± 0.72 | 0.71 ± 0.52 | 0.71 ± 0.43 | 0.47 ± 0.38 |
| PO4− (μmol L−1) | 0.02 ± 0.02 | 0.02 ± 0.02 | 0.04 ± 0.03 | 0.02 ± 0.03 | 0.01 ± 0.01 |
| SiO4− (μmol L−1) | 0.99 ± 0.64 | 1.29 ± 0.36 | 0.7 ± 0.21 | 0.94 ± 0.96 | 0.93 ± 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ordulj, M.; Šantić, D.; Matić, F.; Jozić, S.; Šestanović, S.; Šolić, M.; Veža, J.; Ninčević Gladan, Ž. Analysis of the Influence of Seasonal Water Column Dynamics on the Relationship between Marine Viruses and Microbial Food Web Components Using an Artificial Neural Network. J. Mar. Sci. Eng. 2023, 11, 639. https://doi.org/10.3390/jmse11030639
Ordulj M, Šantić D, Matić F, Jozić S, Šestanović S, Šolić M, Veža J, Ninčević Gladan Ž. Analysis of the Influence of Seasonal Water Column Dynamics on the Relationship between Marine Viruses and Microbial Food Web Components Using an Artificial Neural Network. Journal of Marine Science and Engineering. 2023; 11(3):639. https://doi.org/10.3390/jmse11030639
Chicago/Turabian StyleOrdulj, Marin, Danijela Šantić, Frano Matić, Slaven Jozić, Stefanija Šestanović, Mladen Šolić, Jere Veža, and Živana Ninčević Gladan. 2023. "Analysis of the Influence of Seasonal Water Column Dynamics on the Relationship between Marine Viruses and Microbial Food Web Components Using an Artificial Neural Network" Journal of Marine Science and Engineering 11, no. 3: 639. https://doi.org/10.3390/jmse11030639
APA StyleOrdulj, M., Šantić, D., Matić, F., Jozić, S., Šestanović, S., Šolić, M., Veža, J., & Ninčević Gladan, Ž. (2023). Analysis of the Influence of Seasonal Water Column Dynamics on the Relationship between Marine Viruses and Microbial Food Web Components Using an Artificial Neural Network. Journal of Marine Science and Engineering, 11(3), 639. https://doi.org/10.3390/jmse11030639