
An End-to-End DNA Taxonomy Methodology for Benthic Biodiversity Survey in the Clarion-Clipperton Zone, Central Pacific Abyss

Abstract
:1. Introduction

1.1. The Importance of Combined DNA and Morphological Study

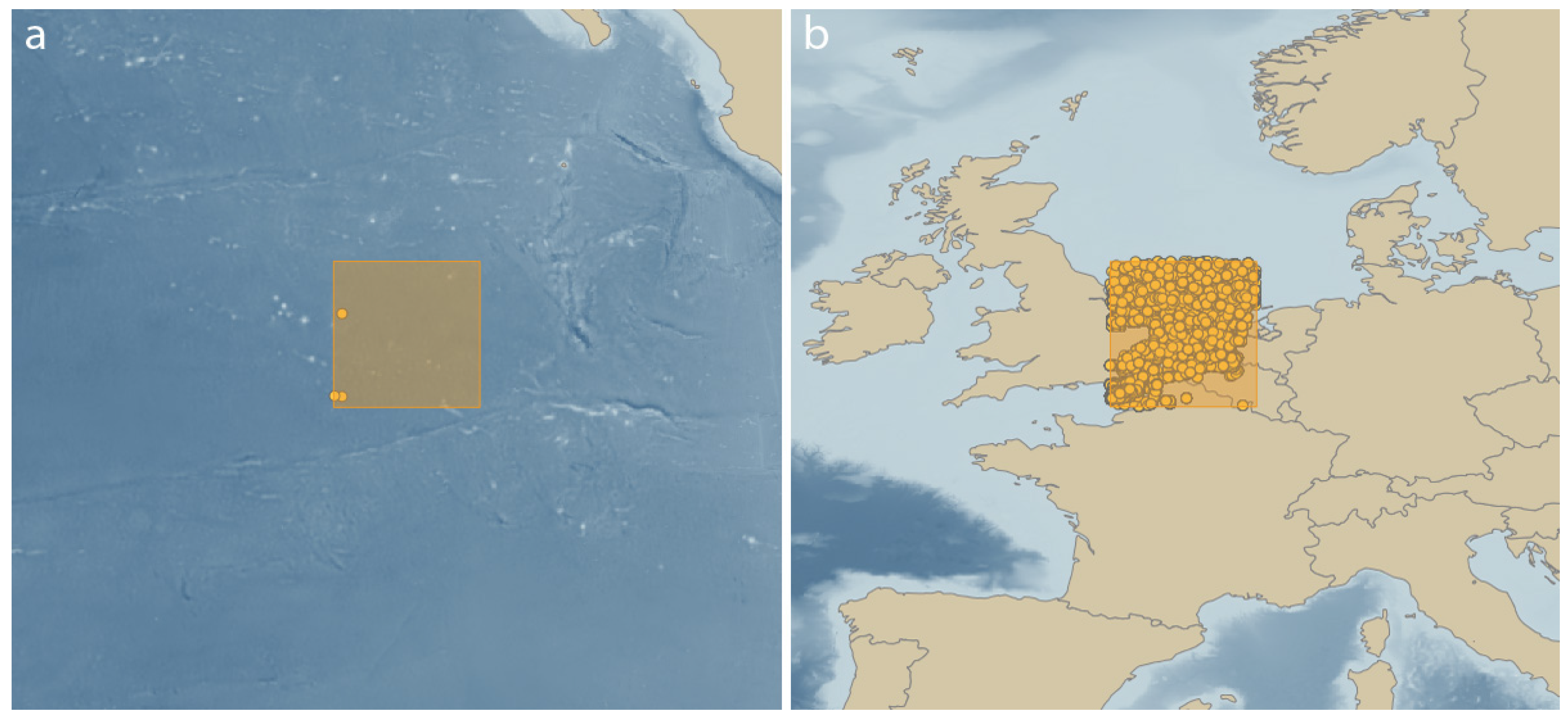
1.2. The Challenges of Zoology in the Clarion-Clipperton Zone


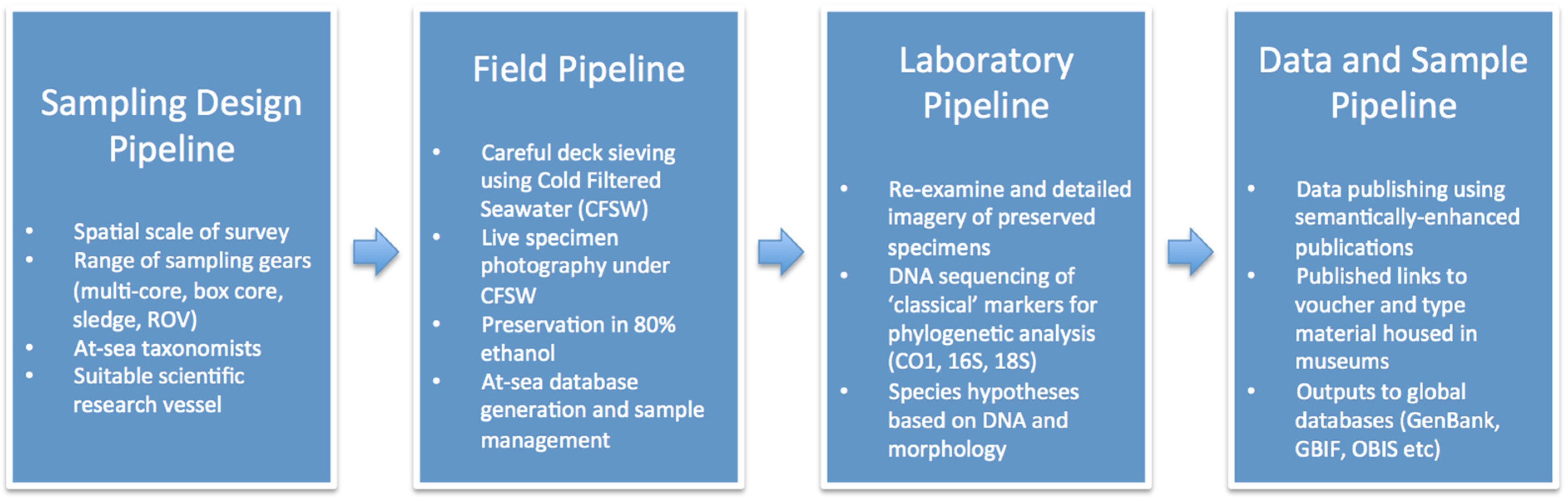
1.3. Methodological Pipelines for DNA Taxonomy in the CCZ
- A sampling design pipeline with consideration to the spatial scale of the required data, the differing biases in sampling gear and the requirement for at-sea taxonomic study
- A field pipeline with consideration to the successful collection of high-quality specimens from depths of 4000–5000 m in the central tropical Pacific
- A laboratory pipeline with consideration to the needs to collect both DNA sequences and morphological data in a timely and cost-effective manner suited to the immediate needs of the science community
- A data and sample management pipeline that includes the publication of results with consideration to the accessibility of data and materials

2. Methodology
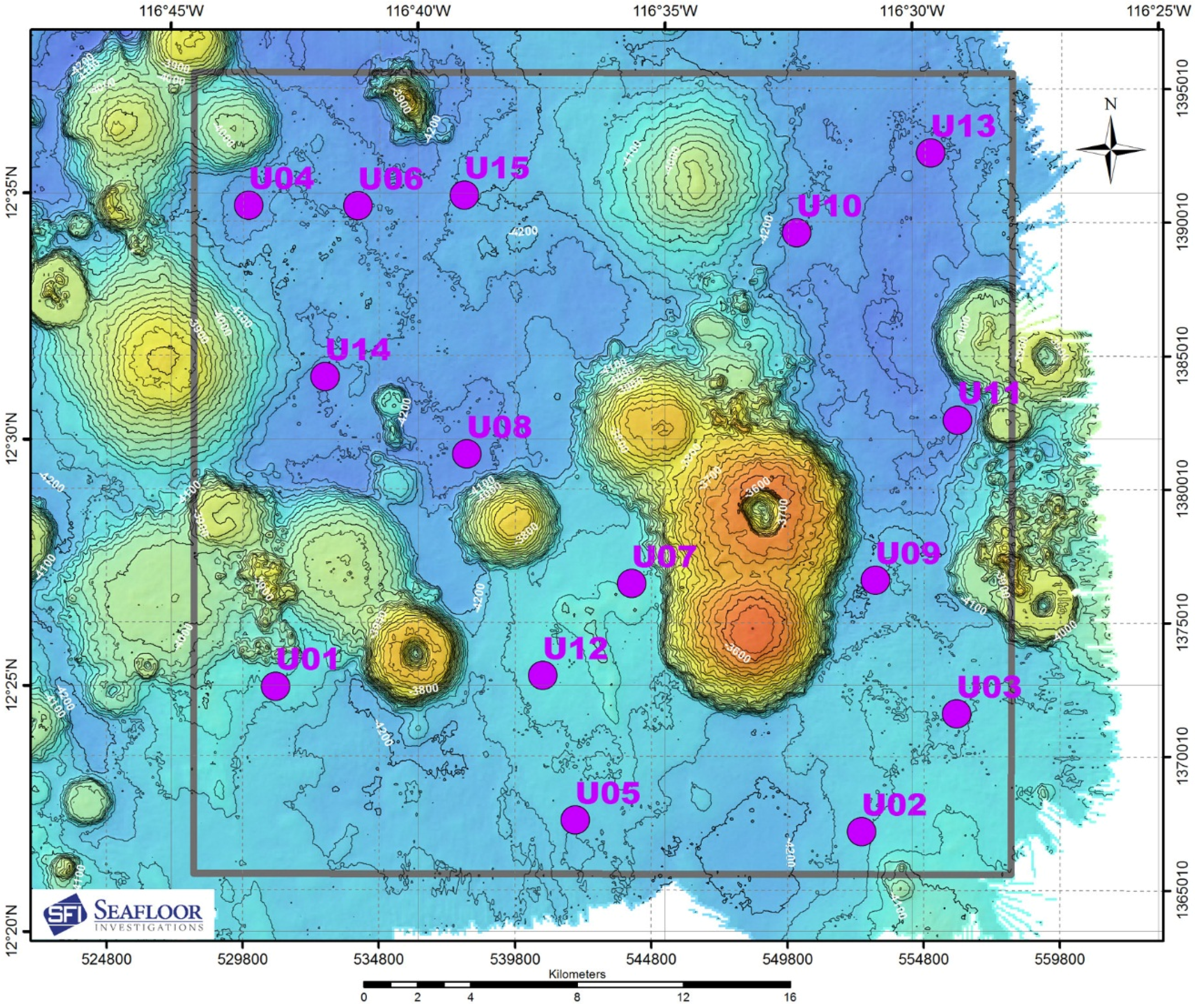
2.1. Sampling Design
- Research vessel to hold position using Dynamic Positioning (DP) at location required
- MC prepared with 12 core tubes and armed and readied as per manufacturer instructions
- MC lifting ring is attached with a swivel to the ship’s trawl wire that is passed through the ship’s starboard A-frame (use of the stern A-frame is not recommended due to pitching)
- MC is lifted off the deck with the combination of A-frame and trawl winch, movement is stopped and the securing pin should be set to “armed”
- Using the A-frame and the trawl winch, the MC is deployed over the side with swing controlled by soft manila hemp tag lines, which are released when the MC is fully submerged in the water
- Wire-out meter is reset with MC at surface, and the MC is then lowered to 50 metres wire out (mwo) and a 12 kHz pinger (1 s time base) is attached to the trawl wire pointing downwards towards the MC
- MC is lowered at 40 m/min until 50 metres above bottom (mab), distance from the MC to the seafloor is monitored using combination of the pinger trace on the ships echosounder (e.g., Kongsberg EA 600) and the wire out meter
- From 50 mab the MC is lowered at 15 m/min until it reaches the seafloor (visible on the pinger trace) and an additional 10 m wire is then paid out
- The winch is then hauled in at 15 m/min until the MC is 50 mab again
- The MC is then recovered at 45 m/min
- MC is recovered with the assistance of tag-lines attached to poles, and secured on deck ready for processing.


- 1.
- Pre-deployment Preparation
- The BX-S50 MK-III BC must be examined and prepared carefully prior to deployment
- The spade lifting cable within the BC is easily damaged from normal wear and tear and from marginal sea states; failure of this cable is the most common loss mode of the box corer. The cable should be carefully inspected for damage (e.g., broken or displaced strands) after each deployment, and typically needs to be changed after every 10–15 deployments (Figure 8)
- 2.
- Deployment
- The BC can be moved around the deck on a pallet with pallet jacks, or it can be positioned for deployment using a crane and tag lines. In the open ocean, the BC is ideally deployed from a side A-frame (Figure 8) or crane to reduce the amplitude of ship motion which can cause pre-tripping of the corer; on large vessels (i.e., >75 m LOA), deployment from a side A-frame can dramatically increase box-coring success
- The Spade Corer should be attached to the trawl wire with a 3-ton swivel; larger swivels can interfere with firing of the corer
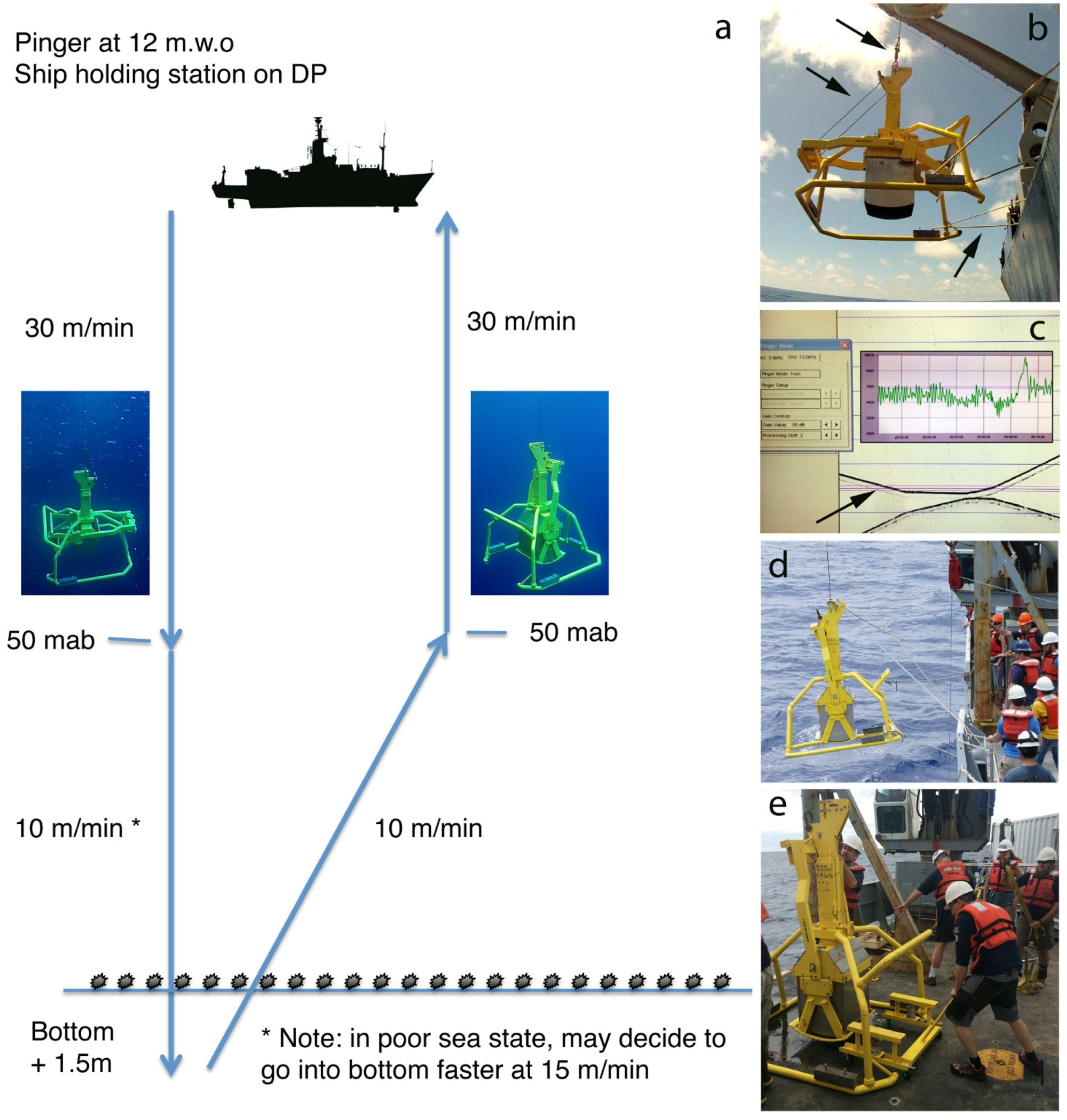
- During deployment, the box corer should be controlled by at least three tag lines to restrain swinging of the corer (Figure 8); tag lines should be pulled after the corer has entered the water. Once in the water, the winch payout should be zeroed, and the box corer lowered to 12 m depth. The pinger is then attached to the wire and the box corer lowered at 30 m per minute to within 50 m of the seafloor, while the ship maintains station (preferably with dynamic positioning) (Figure 8)
- 3.
- Control Room and Seafloor Sampling Protocols
- Early in the deployment, it is important to check that good traces are received from the pinger, the corer and the bottom. The echo from the corer/pinger and the bottom will cross every 750 m above the bottom with a 1 s interval. On the trace, carefully measure the distance between the corer and the pinger, which will increase by 2 m after coring (Figure 8), or if the corer pre-trips. Check the distance regularly to see if a pre-trip has occurred
- When the box corer reaches 50 m above the seafloor, slow the winch speed to 10 m/min and watch the tension plot to observe the normal range in variation before the box core reaches the seafloor. This makes it easier to recognize the drop in tension of about 1000 lbs when the box core reaches the seafloor (note that bottom contact is detectable on the tensiometer before it is visible on the echosounder (Figure 8)). In poor sea states the box core can be put into the seafloor at higher velocities (15–20 m/min) but this typically results in larger bow waves and poorer quality core samples. On the graphical tension display, note when the tension drops ~800–1000 lbs and there is a simultaneous dampening of tension oscillations—this indicates the touchdown of the box corer on the seafloor
- Run out an additional 1.5–2.0 meter of wire after touchdown and then stop the winch and immediately haul up at 10 m/min. As the box core is lifted out of the seafloor (after about 3.5–4.0 m of wire have been hauled in) the winch tension will peak and rapidly drop off to background levels. This tension peak is called the “pullout” tension, and reflects closing of the spade and mud suction as the box corer is pulled out the bottom (Figure 8). For successful abyssal box cores, typical pullout tensions are 2500–3500 lbs above background tensions
- Immediately following pullout (i.e., the box corer leaving the seafloor) check the distance on the echosounder trace between box corer and the pinger; if the box corer has tripped this distance will have increased by 2 m. (Figure 8) If the box corer has not tripped, it may be set it down again, although trip failures can be caused by set down on a hard bottom. If the distance between the box corer and the pinger has decreased, it is likely that the cable has fouled around the box corer and precaution should be taken during box corer recovery. Once the box core is well off the seafloor, it can be hauled back to the surface at 30 m/min without washing the sample
- 4.
- Recovery
- Once the box core has neared ocean surface, tag lines should be attached to the wire below the swivel to limit swinging, and two more attached to the box-corer frame just as it reaches the air-water interface (Figure 8)
- Upon recovery, the BC frame must be gently touched to the deck with tension maintained on the cable to prevent to spade from dropping several centimetres and losing the sample topwater; while the box corer is in this position, the dolly-cart should be quickly wheeled under the spade and the carted jacked up to support the spade and box (Figure 8). In addition, the top of the spade arm must be quickly secured to the box-core frame to hold the spade in the vertical position. Once the steps are taken, the sample is secure and can be methodically removed from the box corer (see BC processing protocol, below). The wire tension can also be released and the box corer frame secured to the deck



2.2. Field Pipeline
2.2.1. Cold Filtered Seawater (CFSW) System Protocol
- Pumped seawater supply system from the ship (this may claim to be filtered, but almost always requires additional filtration as detailed below). Note that these systems are standard on oceanographic research vessels
- A large constant temperature (CT) van or room that can be set to 5 °C or lower (ideally 2 °C) with easy access to the back deck. The chilling system or van needs to be robust and of high quality to enable operations in high humidity, high temperature for months at sea. This system is not standard on oceanographic vessels and must be hired or purchased
- Water reservoir of as large a volume as can be achieved, ideally something in the region of 750–1000 litres (200–300 US gallons). A low-cost option is 5 × 170 litre (44 US gallon) plastic trash cans, these need to be secured carefully in the CT room in case of ship movement. An alternative is high-quality water storage reservoirs built into the CT lab that can be plumbed in and connected together using a flow-through system.
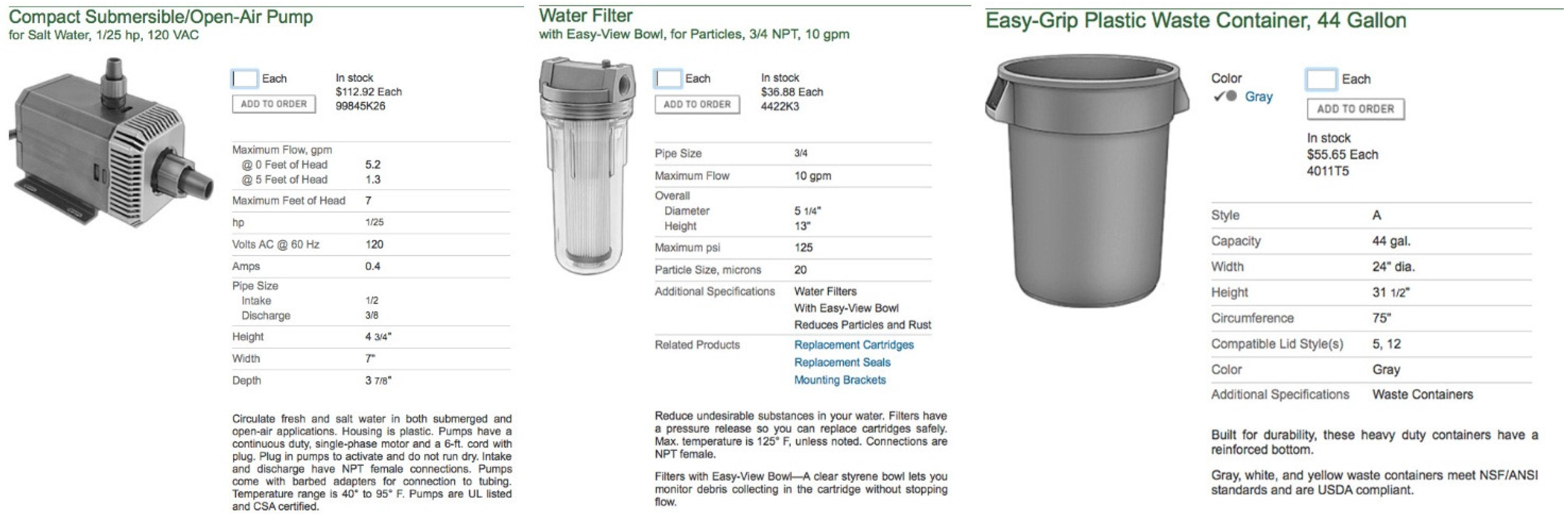
- At least 3 saltwater submersible pumps with approx 1/25 hp rating (Figure 11)
- Garden hose connectable water filter system (20 µm rating) with replaceable cartridges
- ~50 m of garden hose
- ~15 m of clear soft 12 mm (½ inch) “tygon tube” type plastic hose
- Misc hose connectors to connect filters and pumps to hoses, including adaptor from the ship’s outlet to the garden hose
- Some flow control valves and “Y” splitter valves to put in various places (e.g., at end of hose near sieve, next to filters, etc.).
- 2–3 large plastic trays 10–20 cm deep approx 80 × 50 cm that will fit the sieves and if you are sorting in a lab, fit in the sink (see processing protocol)
- Most items can be purchased in any good large hardware store (Figure 11)


- Install 5 × 170 litre (44 gallon) drums, or similar water storage system (e.g., flow-through connected drums) in the CT room or CT van, secured
- Connect ship seawater system to water filter on deck, and run from filter with garden hose into the CT room, with flow valves inside to control flow into the drums
- Install a salt-water submersible pump in the CT room tanks, with control switch inside CT room
- Run 12 mm (½ inch) tygon (clear plastic hose) from submersible pumps back out of the CT room onto the deck, with Y-splitter and control valves as required
- Typical usage to process 1 box core is 190–380 litres (50–100 gallons) of CFSW
- Typical time to chill 190 litres (50 gallons) surface water from the CCZ at 28 °C in a 2 °C CT room to ~4 °C is 4 h
2.2.2. Box Core Processing Protocol (for DNA Taxonomy)

- Supply of CFSW from cold van, 5–6 wash bottles pre-filled with CFSW
- Plastic 20 litre buckets labelled 0–2 cm, 2–5 cm, 5–10 cm, each pre-filled 1/3 with CFSW
- 3–4 plastic trays for nodules
- 300 µm sieve (2) with deep trays to hold sieves inside
- Short (1.5 m) lengths of vinyl hose for siphoning water from top of box core
- Large syringe or turkey basters for siphoning water from top of box core
- Forceps and small dishes/trays for megafauna to be collected immediately
- Trowels for cutting mud
- Camera, flashlight, thermometer
- Examine box core surface with flashlight
- Measure top water temperature (make note)
- With two tubes, drain some of the surface water through 300 µm sieve but without disturbing top layer
- Roll box core out on trolley (slowly) and secure on deck in shade.
- Drain remaining top water through 300 µm sieve and use turkey basters or syringes, etc., for remainder of the water
- Photograph exposed surface with label
- Pick obvious unattached megafauna to place in small CFSW buckets/dishes on ice
- Remove nodules carefully, rinse each nodule in the 0–2 cm bucket and with CFSW wash bottles
- Place nodules onto a nodule-storing foam tray or egg tray from galley, keeping them upright → tray to lab (keep cool)
- Remove sub-section (depending on amount of sample to live sort) of 0–2 cm for DNA taxonomy/live sorting, place into 0–2 cm CFSW bucket and store in cold van
- Repeat with 2–5 cm and 5–10 cm sections
- Sieve samples with 2 people, 1 to hold sieve and 1 person to pour water and collect sample
- Gently agitate buckets of mud and pour slowly onto sieve, keeping sieve underwater in tray (Figure 13)
- Tap sieve gently from underneath to help material go through sieve if necessary
- Regularly wash material (with wash bottle) retained on the sieve to a small beaker, as soon as material is in this dish store in the lab (fridge) for live sorting/DNA taxonomy
2.2.3. Epibenthic Sledge (EBS) or Trawl Processing Protocol
- Supply of CFSW from cold van, 5–6 wash bottles pre-filled with CFSW
- ~1 m wide 50 litre tubs labelled “epi” and “supra” net for the two cod-end samples from the EBS
- 3–4 plastic trays for nodules
- 300 µm sieve (2) with deep trays to hold sieves inside
- Forceps and small dishes/trays for megafauna to be collected immediately
- Remove cod-ends into CFSW “epi” and “supra” buckets (rinse cod-ends with CFSW from hose)
- Wash material on nets into buckets (with CFSW)
- Remove remaining nodules to large buckets
- Photograph sample
- Pick megafaunal animals into CFSW trays or small dishes -> transfer to lab for DNA taxonomy
- Sieve EBS mud/cod-end sample with submerged 300 µm sieve in CFSW as for box core processing (Figure 13), transfer sample to small beakers to take to lab for DNA taxonomy/live sorting
- Pick through remaining large “gravel” fraction using white trays outside (Figure 13)
2.2.4. Large Megafauna Processing Protocol (e.g., ROV, HOV or Trawl Collected)
- Supply of CFSW from cold van, 2–3 wash bottles pre-filled with CFSW
- Selection of small and large buckets pre-filled with CFSW to receive ROV samples
- 300 µm sieve (2) with deep trays to hold sieves inside
- Forceps and small dishes/trays for megafauna to be collected immediately
- Collect specimens by hand from ROV or HOV bioboxes/storage trays
- Maintain specimens in buckets or trays of CFSW and remove to lab for live sorting
- Wash remaining biobox water through 300 µm sieves (as with EBS protocol) to collect smaller fauna and associated fauna
2.2.5. Live Sorting for DNA Taxonomy Protocol
- ~6 m of laboratory bench space inside a comfortable research vessel general purpose lab with chairs suited to microscope work (Figure 14)
- Easy access fridge for temporary live-sample storage, and −20 °C freezer for storage of preserved specimens
- Machine for crushed ice (not ice cubes)
- 2× high-quality stereomicroscopes (e.g., Leica MZ9.5 or equivalent) equipped with photography tube
- 2× high-quality DSLR cameras attached to microscopes (e.g., Canon 700D or equivalent) together with 4× strobe (flashguns) equipped with eTTL and wireless (slave) mode (e.g., Canon 430EXII or equivalent)
- 2× full HD 1080p screens to output HDMI live view from the DSLR cameras
- 2× high-power fibre optic light sources (e.g., Leica CLS150 or equivalent)
- 1× macro-photography suite consisting of table tripod, DSLR camera with 45 mm (90 mm in 35 mm equivalent) macro lens, light source (flexible fibre optic work lamps that clamp to bench, low cost and effective is IKEA “Tived” LED clamp light article 701.696.94)
- Acrylic trays for photography and ice baths (Figure 14)
- General laboratory sorting consumables including 2 ml, 20 mL and 50 mL plastic microtubes (e.g., Sarstedt), plastic pipettes, dissection kits, wash bottles of CFSW, tube racks (including low-temperature gel pack tube racks), labels for jars (Figure 14)
- Compact desktop computer (e.g., Mac Mini) for storing sample database and images from cameras
- Supply of 80% ethanol in wash bottles, kept chilled. This needs to be non-denatured ethanol that is diluted to 80% with laboratory DI water (not tapwater or seawater). It is important that the ethanol used is 80% absolute ethanol (non-denatured) in 20% DI water, not pure 100% ethanol as this greatly reduces the dehydration and physical damage to the specimens, allowing them to be used for good quality morphological study as well as DNA. As the ethanol is only used in the final storage of the specimens in the microtube vials, the amount of ethanol required equates to the number of final vials stored. As a guideline, on the recent March 2015 ABYSSLINE cruise to the CCZ (a 42 day cruise using a range of sampling gear) we used ~1500 2 mL microtubes and ~100 20 mL vials a volume of ethanol equal to 5 liters (although only macrofauna and DNA tissue samples from large animals were taken). We would recommend taking 2× the amount of ethanol calculated and consumables on top of that anticipated, and in particular if large megafauna are to be fixed intact then obviously larger quantities are needed, or large megafauna (20 cm long holothurians, etc.) can be fixed in low-cost 10% buffered formalin using standard methods with just a small tissue sample (e.g., body wall, muscle) taken for DNA.

- Place the specimen (or nodule) into a clean CFSW water bath (using acrylic trays) fully submerged with a wet black cloth under the tray to create a black background. This can be achieved by placing 1 bath inside another, with ice surrounding the internal bath (Figure 14).
- Photograph the specimen or nodule with macro-lens DSLR held in tripod with good lighting (powerful LED lights are sufficient, strobe not required unless the specimen is moving) on black background with scale bar.
- For nodule fauna, remove individual specimens after photography and if necessary image under the photomicroscope camera for higher-resolution.
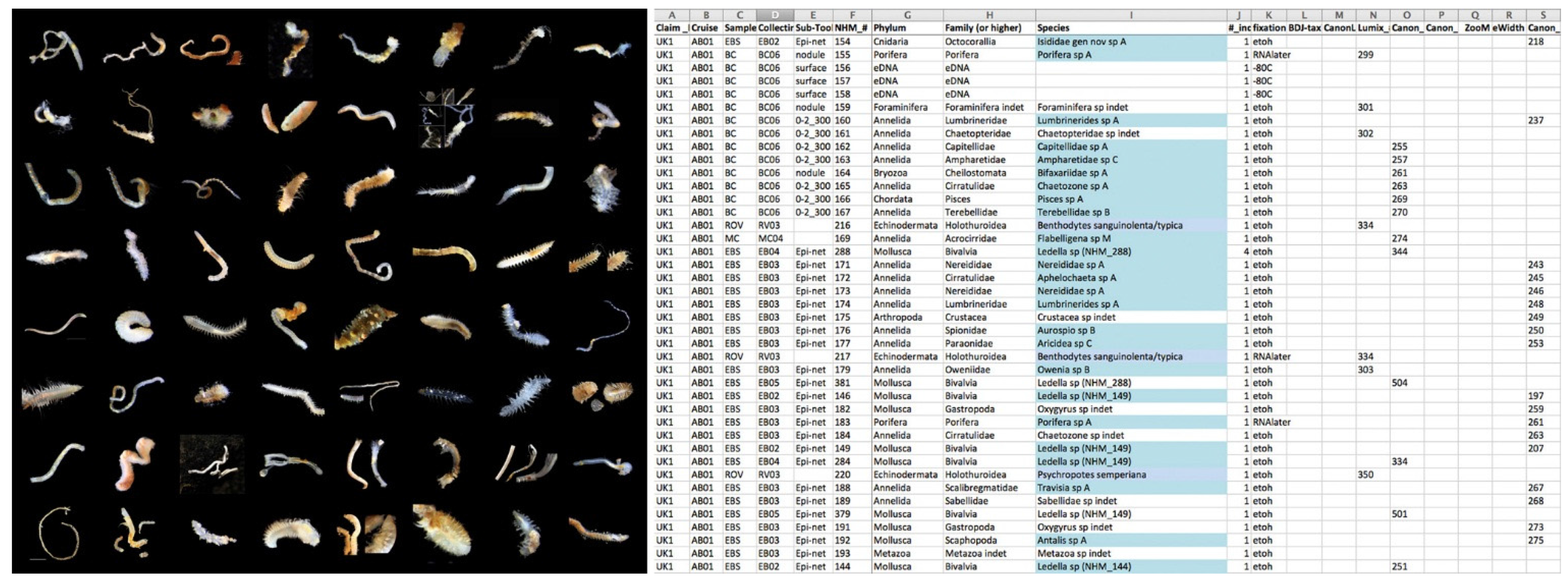
- Using the spreadsheet database, assign a specimen number and add a record of the camera photograph number (normally the best one). Make sure the spreadsheet has enough columns to cover various different cameras. Add any other relevant data (gear, station, preliminary identification, notes, etc.).
- For very large specimens (e.g., big sponges) remove a small piece of tissue and place in vial labelled with the specimen number. Add chilled 80% ethanol and store in chilled tube racks. Remainder of specimen can be fixed in formalin or ethanol in a large jar with the same specimen number.
- For small specimens the whole animal can go in a microtube vial (normally 2 mL or 20 mL in size for CCZ organisms). Label and top up with chilled 80% ethanol.
- All data recorded to an on-the-fly lab live sorting spreadsheet (Figure 15).
- Depending on fixed use, some samples may be fixed in RNAlater (e.g., for metagenomic eDNA studies) or −80 °C freezer.

- Small aliquots of the sieved residues are transferred to petri dishes with CFSW. The petri dishes are then sorted under the microscope using standard picking methods. The petri dishes can be kept cool by placing them on pre-chilled microscope bases (kept in freezer) or pre-chilled, wet black cloths
- Picked animals are sorted initially into major taxon under the microscope (bivalves, polychaetes, etc.) and kept in small petri dishes on ice (Figure 14)
- Once the sort is complete, the residues for the box core sample (i.e., sorted waste) is returned to the quantitative box core sample. Unwanted EBS sample can be discarded or bulk-fixed
- Specimens can be cleaned by transferring to new petri dishes and fresh CFSW, for photography.
- Specimens are photographed using the photomicroscope and strobes (Figure 14)
- Specimens are transferred to 2 mL microtubes with 80% ethanol and kept chilled, data is added to the spreadsheet as for the large/nodule fauna protocol above
- Photographic tips. A clean specimen in clean CFSW on a new petri dish, the dish supported on a wet black cloth (under the dish) helps to achieve a high-resolution image with black background (Figure 15). Periodically wetting the black cloth with chilled seawater also helps to keep the specimen cool
- Photographic tips. Specimens should be imaged using two strobes in wireless (slave mode) with the image taken using a remote switch (to avoid vibration). Typically, white balance should be carefully adjusted to conform to the strobe lighting, and the exposure compensation function used to prevent over-exposure of any part of the specimen. Patience is required to achieve good photographs
2.3. Laboratory Pipeline
- All specimens are re-examined using high-power microscopy and imaged for morphological features (e.g., polychaete chaetae) (Figure 16)
- A small tissue sample is taken for DNA sequencing, this can be an individual polychaete parapodia, antenna or slice of body wall. A simple rule is that if the tissue sample is visible to the naked eye it is big enough for DNA sequencing. A typical minimum we use is a piece of tissue of 0.5 mm × 0.5 mm, roughly the size of the end of a fine pencil
- Remainder of the specimen is kept for morphological study or further DNA sampling if required
- Species hypotheses are created later based on DNA phylogenetics (not before)

- Extraction of DNA is undertaken using DNeasy Blood and Tissue Kit (Qiagen) using a Hamilton Microlab STAR Robotic Workstation (Figure 16)
- Typically, about 1800 bp of 18S, 800 bp of 28S (D1-D2 region), 450 bp of 16S, and 650 bp of cytochrome c oxidase subunit I (COI) are amplified using either published or newly-designed primers (depending on the taxa of choice). Primer pair combination for 16S that is working on most CCZ phyla is Ann16SF together with 16SbrH. Furthermore, the COI primers originally designed for polychaetes, polyLCO-polyHCO (Table 1), works well on a range of other phyla, e.g., Bryozoa, Cnidaria, Echinodermata, Mollusca and Chordata
- Typically, PCR mixtures contain 1 µL of each primer (10 µM), 2 µL template DNA and 21 µL of Red Taq DNA Polymerase 1.1× MasterMix (VWR) in a mixture of total 25 µL. The PCR amplification profile consists of initial denaturation at 95 °C for 5 min, 35 cycles of denaturation at 94 °C for 45 s, annealing at 55 °C for 45 s, extension at 72 °C for 2 min, and a final extension at 72 °C for 10 min
- Typically, PCR products were purified using Millipore Multiscreen 96-well PCR Purification System, and sequencing was performed on an ABI 3730XL DNA Analyser (Applied Biosystems) at Sequencing Facility, using the same primers as in the PCR reactions plus two internal primers for 18S
- Bayesian phylogenetic analyses (BA) are conducted with MrBayes 3.1.2 [32] following standard methods for phylogenetic inference

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence 5′-3′ | Reference |
|---|---|---|
| 18S | ||
| 18SA | AYCTGGTTGATCCTGCCAGT | [33] |
| 18SB | ACCTTGTTACGACTTTTACTTCCTC | [34] |
| 620F | TAAAGYTGYTGCAGTTAAA | [34] |
| 1324R | CGGCCATGCACCACC | [35] |
| 28S (D1-D2) | ||
| C1′ | ACCCGCTGAATTTAAGCAT | [36] |
| D2 | AACTCTCTCMTTCARAGTTC | [36] |
| COI | ||
| LCO1490 | GGTCAACAAATCATAAAGATATTGG | [37] |
| HCO2198 | TAAACTTCAGGGTGACCAAAAAATCA | [37] |
| COI-E | TATACTTCTGGGTGTCCGAAGAATCA | [38] |
| polyLCO | GAYTATWTTCAACAAATCATAAAGATATTGG | [39] |
| polyHCO | TAMACTTCWGGGTGACCAAARAATCA | [39] |
| 16S | ||
| 16SarL | CGCCTGTTTATCAAAAACAT | [40] |
| 16SbrH | CCGGTCTGAACTCAGATCACGT | [40] |
| Ann16SF | GCGGTATCCTGACCGTRCWAAGGTA | [41] |
| Ann16SR | TCCTAAGCCAACATCGAGGTGCCAA | [41] |
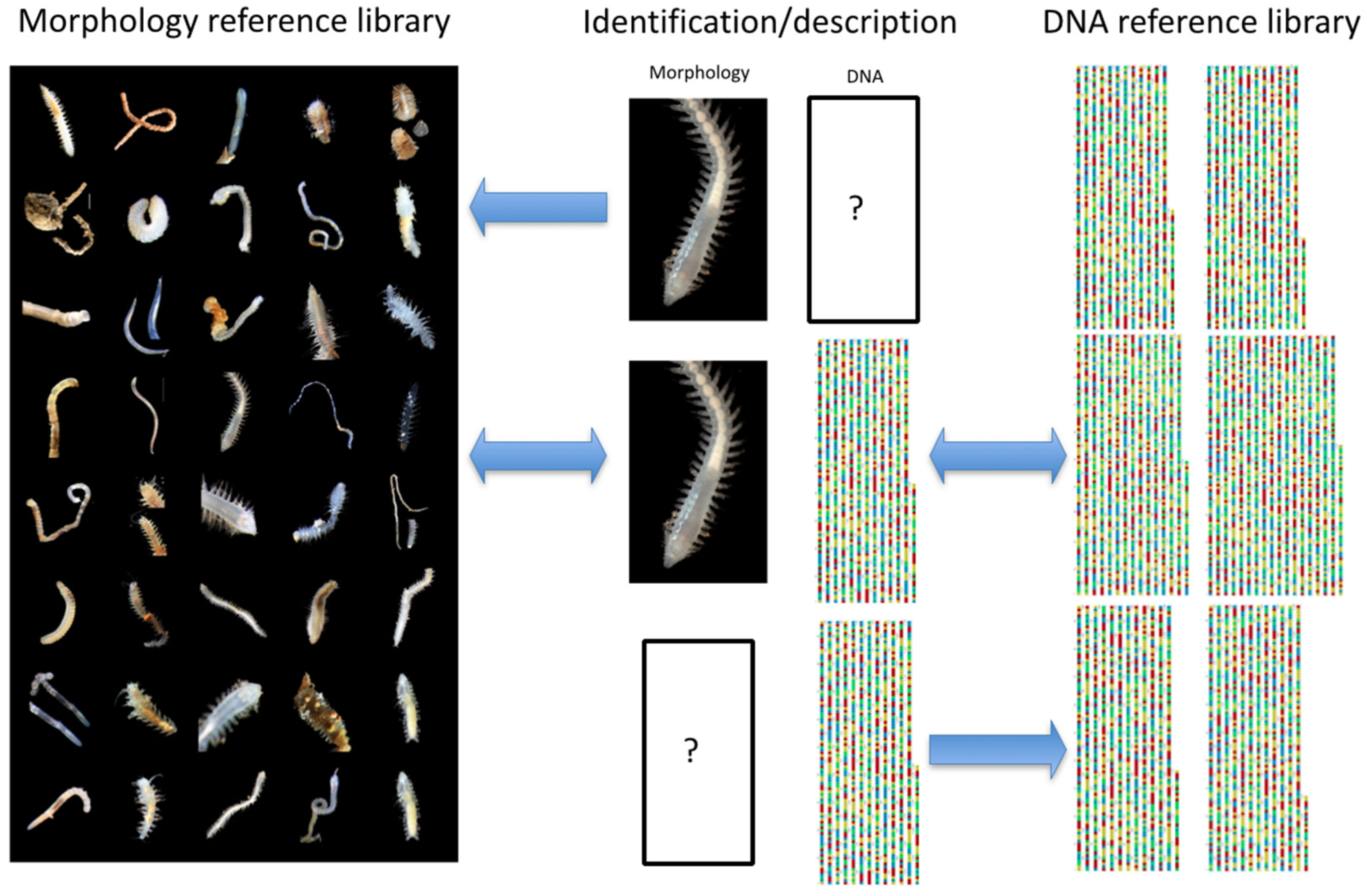
- Statistically well-supported clades where “within-clade” sequence variation is lower than “between-clade” variation are hypothesised as species (Figure 17)
- Morphological data is condensed for each species concept, and a type, series of types or voucher allocated
- Taxonomic species descriptions or records based primarily on genetic data supported by morphological information to aid identification in the field are produced in a taxonomic data paper format (Figure 17)
- Site and associated data are organised into the bioinformatics-standard Darwin Core (DwC) [45] format to support online publication and data management (Figure 17 and see Section 2.4)

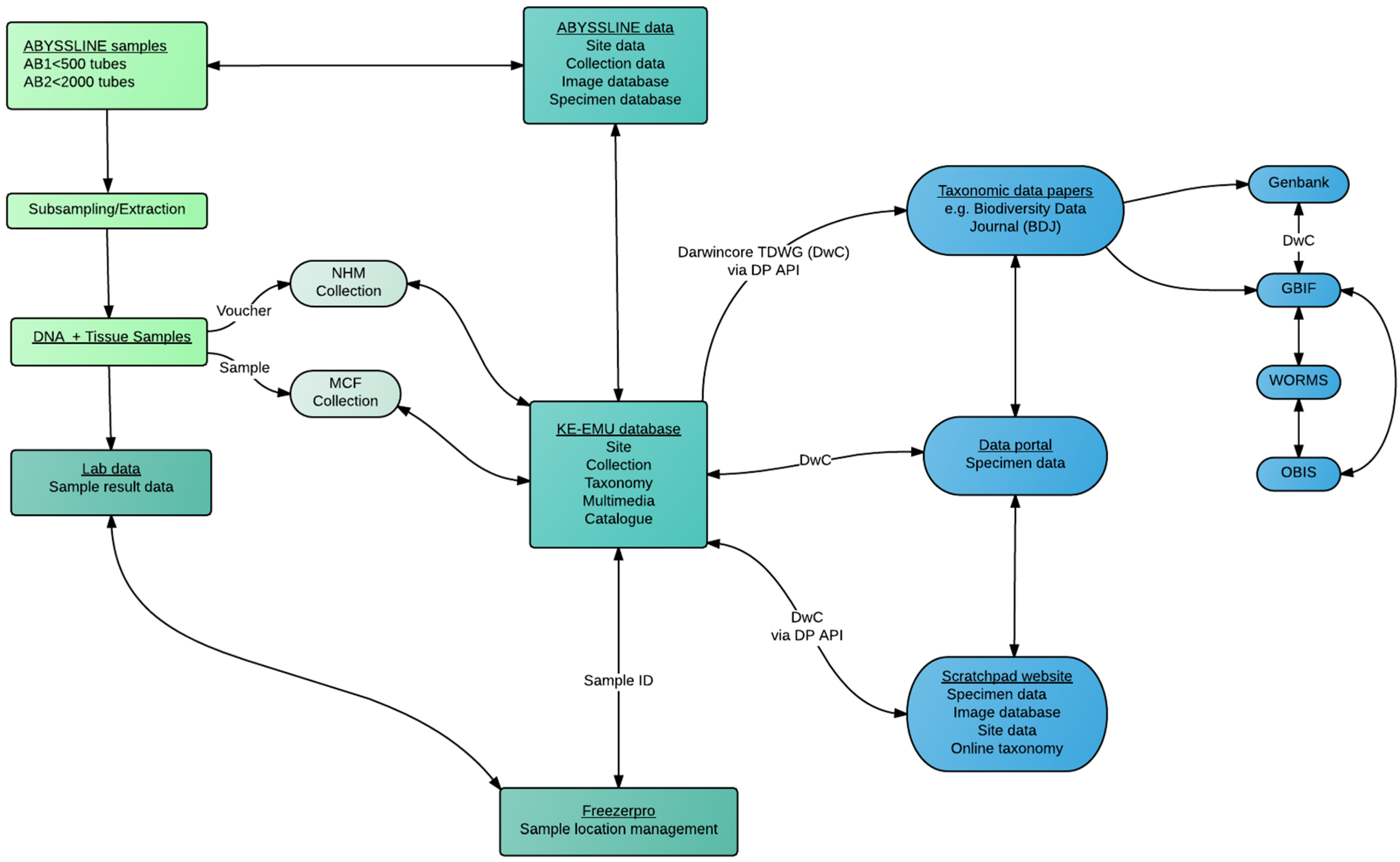
2.4. Data and Sample Management Pipeline
- The data and sample products from the field and laboratory pipelines (which can reach many thousands of physical objects and data points—e.g., Figure 18) are fed into institutional-level databases, in the case of ABYSSLINE this is the KE-EMU database (a central NHM database) and the FreezerPro database (a database for all frozen tissue samples in the Molecular Collection Facility (MCF) of the NHM)
- The DwC outputs are also linked to the taxonomic data papers, in most cases using semantically-enhanced publications such as Biodiversity Data Journal [50]
- The physical samples are stored in two long-term collections—the NHM national collection for ethanol-fixed specimens and the NHM Molecular Collection Facility where tissue samples and DNA template is stored at −80 °C freezers with backup power systems. These samples are maintained by the NHM curatorial staff, and accessible to researchers (Figure 19)
- The DwC outputs from the taxonomic data papers (e.g., Biodiversity Data Journal) are automatically made available to external databases including GBIF and OBIS, with DNA sequences available on GenBank
- DwC outputs can also be used for a project-level (multiple institution) public-facing or project website to enhance collaboration and data sharing amongst project members during the project phase, implemented in the Scratchpad
- All external use of the data or samples is linked to the peer-reviewed data paper which allow data users a stable DOI citation to use as well as contact details for the authors, and associated data


3. Discussion and Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- International Seabed Authority Home Page. Available online: http://www.isa.org.jm (accessed on 14 December 2015).
- Nimmo, M.; Morgan, C.; Banning, D. Clarion-Clipperton Zone Project, Pacific Ocean: Updated NI 43–101 Technical Report; 127631013–003-R-Rev1; Golder Associates Pty Ltd.: Milton, Australia, 2013. [Google Scholar]
- ISA. Recommendations for the Guidance of Contractors for the Assessment of the Possible Environmental Impacts Arising from Exploration for Polymetallic Nodules in the Area; International Seabed Authority: Kingston, Jamaica, 2010. [Google Scholar]
- Seascape Consultants. Review of Implementation of the Environmental Management Plan for the Clarion-Clipperton Zone, Report to the International Seabed Authority, final version; Seascape Consultants Ltd.: Romsey, UK, 2014. [Google Scholar]
- ISA. Report of the Chair of the Legal and Technical Commission on the Work of the Commission during the Twenty-First Session of the International Seabed Authority; ISBA/21/C/16; International Seabed Authority: Kingston, Jamaica, 2015. [Google Scholar]
- ICZN. In International Code for Zoological Nomenclature; The International Trust for Zoological Nomenclature: London, UK, 1999.
- Summers, M.M.; Al-Hakim, I.I.; Rouse, G.W. Turbo-taxonomy: 21 New species of Myzostomida (Annelida). Zootaxa 2014, 3873, 301–344. [Google Scholar] [CrossRef] [PubMed]
- Davey, J.W.; Hohenlohe, P.A.; Etter, P.D.; Boone, J.Q.; Catchen, J.M.; Blaxter, M.L. Genome-wide genetic marker discovery and genotyping using next-generation sequencing. Nat. Rev. Genet. 2011, 12, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Willette, D.A.; Allendorf, F.W.; Barber, P.H.; Barshis, D.J.; Carpenter, K.E.; Crandall, E.D.; Cresko, W.A.; Fernandez-Silva, I.; Matz, M.V.; Meyer, E.; et al. So, you want to use next-generation sequencing in marine systems? Insight from the pan-pacific advanced studies institute. Bull. Mar. Sci. 2014, 90, 79–122. [Google Scholar] [CrossRef]
- NOAA. Volumes of the World’s Oceans from Etopo1. Available online: http://www.ngdc.noaa.gov/mgg/global/etopo1_ocean_volumes.html (accessed on 14 December 2015).
- Rex, M.A.; McClain, C.R.; Johnson, N.A.; Etter, R.J.; Allen, J.A.; Bouchet, P.; Warén, A. A source-sink hypothesis for abyssal biodiversity. Am. Nat. 2005, 165, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Lutz, M.; Dunbar, R.; Caldeira, K. Regional variability in the vertical flux of particulate organic carbon in the ocean interior. Glob. Biogeochem. Cycle 2002, 16. [Google Scholar] [CrossRef]
- Smith, C.R.; Paterson, G.L.J.; Lambshead, P.J.D.; Glover, A.G.; Rogers, A.D.; Gooday, A.; Kitazato, H.; Sibuet, M.; Galeron, J.; Menot, L. Biodiversity, Species Ranges, and Gene Flow in the Abyssal Pacific Nodule Province: Predicting and Managing the Impacts of Deep Seabed Mining: Report; ISA Technical Study No.3; International Seabed Authority: Kingston, Jamaica, 2008. [Google Scholar]
- Glover, A.G.; Smith, C.R.; Paterson, G.L.J.; Wilson, G.D.F.; Hawkins, L.; Sheader, M. Polychaete species diversity in the central pacific abyss: Local and regional patterns, and relationships with productivity. Mar. Ecol. Prog. Ser. 2002, 240, 157–169. [Google Scholar] [CrossRef]
- Smith, C.R.; Berelson, W.; Demaster, D.J.; Dobbs, F.C.; Hammond, D.; Hoover, D.J.; Pope, R.H.; Stephens, M. Latitudinal variations in benthic processes in the abyssal equatorial pacific: Control by biogenic particle flux. Deep Sea Res. II 1997, 44, 2295–2317. [Google Scholar] [CrossRef]
- OBIS. Global Biodiversity Indices from the Ocean Biogeographic Information System. Intergovernmental Oceanographic Commission of UNESCO. Available online: http://www.iobis.org (accessed on 14 December 2015).
- ISA. Rationale and Recommendations for the Establishment of Preservation Reference Areas for Nodule Mining in the Clarion-Clipperton Zone; ISBA/14/LTC/2*; International Seabed Authority: Kingston, Jamaica, 2008. [Google Scholar]
- Smith, C.R.; Gaines, S.; Friedlander, A.; Morgan, C.; Thurnherr, A.; Mincks, S.; Watling, L.; Rogers, A.D.; Clark, M.; Baco-Taylor, A.; et al. Preservation Reference Areas for Nodule Mining in the Clarion-Clipperton Zone: Rationale and Recommendations to the International Seabed Authority; International Seabed Authority: Kingston, Jamaica, 2008. [Google Scholar]
- Wedding, L.M.; Friedlander, A.M.; Kittinger, J.N.; Watling, L.; Gaines, S.D.; Bennett, M.; Hardy, S.M.; Smith, C.R. From principles to practice: A spatial approach to systematic conservation planning in the deep sea. Proc. R. Soc. B 2013, 280, 20131684. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.R.; Dahlgren, T.G.; Drazen, J.C.; Glover, A.G.; Gooday, A.; Kurras, G.; Martinez-Arbizu, P.; Shulse, C.; Spickermann, R.; Sweetman, A.K.; et al. Abyssal Baseline Study (ABYSSLINE) Cruise Report. Available online: http://abyssline.info/content/abyssline-cruises (accessed on 18 December 2015).
- Smith, C.R.; Church, M.; Dahlgren, T.G.; Drazen, J.C.; Glover, A.G.; Gooday, A.; Martinez-Arbizu, P.; Sweetman, A.K.; Vetter, E. The abyssal baseline study (ABYSSLINE). Unpublished work. 2015. [Google Scholar]
- Barnett, P.R.O.; Watson, J.; Connelly, D. A multiple corer for taking virtually undisturbed samples from shelf, bathyal and abyssal sediments. Oceanol. Acta 1984, 7, 399–408. [Google Scholar] [CrossRef]
- Hessler, R.R.; Jumars, P.A. Abyssal community analysis from replicate box cores in the central north pacific. Deep Sea Res. 1974, 21, 185–209. [Google Scholar]
- Hessler, R.R.; Sanders, H.L. Faunal diversity in the deep-sea. Deep Sea Res. 1967, 14, 65–78. [Google Scholar] [CrossRef]
- Brenke, N. An epibenthic sledge for operations on marine soft bottom and bedrock. Mar. Technol. Soc. J. 2005, 39, 10–21. [Google Scholar] [CrossRef]
- Priede, I.G.; Bagley, P.M.; Smith, A.; Creasey, S.; Merrett, N.R. Scavenging deep demersal fishes of the porcupine seabight, north-east atlantic: Observations by baited camera, trap and trawl. J. Mar. Biol. Assoc. UK 1994, 74, 481–498. [Google Scholar] [CrossRef]
- ISA. Workshop on Taxonomic Methods and Standardization of Macrofauna in the Clarion-Clipperton Fracture Zone. Available online: http://www.isa.org.jm/workshop/workshop-taxonomic-methods-and-standardization-macrofauna-clarion-clipperton-fracture-zone (accessed on 14 December 2015).
- Lejzerowicz, F.; Esling, P.; Pillet, L.; Wilding, T.A.; Black, K.D.; Pawlowski, J. High-throughput sequencing and morphology perform equally well for benthic monitoring of marine ecosystems. Sci. Rep. 2015, 5, 13932. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. Mafft: A novel method for rapid multiple sequence alignment based on fast fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. Mrbayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16s-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef]
- Nygren, A.; Sundberg, P. Phylogeny and evolution of reproductive modes in Autolytinae (Syllidae, Annelida). Mol. Phylogenet. Evol. 2003, 29, 235–249. [Google Scholar] [CrossRef]
- Cohen, B.L.; Gawthrop, A.; Cavalier-Smith, T. Molecular phylogeny of brachiopods and phoronids based on nuclear-encoded small subunit ribosomal RNA gene sequences. Philos. Trans. R. Soc. B 1998, 353, 2039–2061. [Google Scholar] [CrossRef]
- Lê, H.L.V.; Lecointre, G.; Perasso, R. A 28s rRNA based phylogeny of the gnathostomes: First steps in the analysis of conflict and congruence with morphologically-based cladograms. Mol. Phylogenet. Evol. 1993, 2, 31–51. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hueh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit i from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Bely, A.E.; Wray, G.A. Molecular phylogeny of naidid worms (Annelida: Clitellata) based on cytochrome oxidase I. Mol. Phylogenet. Evol. 2004, 30, 50–63. [Google Scholar] [CrossRef]
- Carr, C.M.; Hardy, S.M.; Brown, T.M.; Macdonald, T.; Hebert, P.D.N. A tri-oceanic perspective: DNA barcoding reveals geographic structure and cryptic diversity in Canadian polychaetes. PLoS ONE 2011, 2011, e22232. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R. Nucleic acid II: The polymerase chain reaction. In Molecular Systematics; Hillis, D.M., Moritz, G., Mable, B., Eds.; Sinauer Associates: Sunderland, MA, USA, 1996; pp. 205–247. [Google Scholar]
- Sjölin, E.; Erséus, C.; Källersjö, M. Phylogeny of Tubificidae (Annelida, Clitellata) based on mitochondrial and nuclear sequence data. Mol. Phylogenet. Evol. 2005, 2005, 431–441. [Google Scholar] [CrossRef] [PubMed]
- De Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, M.J. A critique of the biological species concept and recommendations for a phylogenetic alternative. Bryologist 1985, 88, 172–181. [Google Scholar] [CrossRef]
- Nygren, A. Cryptic polychaete diversity: A review. Zool. Scr. 2014, 43, 172–183. [Google Scholar] [CrossRef]
- GBIF. Darwin core Archives—How-to Guide, Version 1, Released on 1 March 2011; Global Biodiversity Information Facility: Copenhagen, Denmark, 2010. [Google Scholar]
- Glover, A.G.; Wiklund, H.; Rabone, M.; Amon, D.J.; Smith, C.R.; O'Hara, T.; Mah, C.; Dahlgren, T.G. Abyssal fauna of the uk-1 polymetallic nodule exploration claim, clarion-clipperton zone, central pacific ocean: Echinodermata. Biodivers. Data J. 2015. submitted for publication. [Google Scholar]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. Genbank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [PubMed]
- Global Biodiversity Information Facility. Available online: http://www.gbif.org (accessed on 14 December 2015).
- Boxshall, G.A.; Mees, J.; Costello, M.J.; Hernandez, F.; Gofas, S.; Hoeksema, B.W.; Klautau, M.; Kroh, A.; Paulay, G.; Poore, G.; et al. World Register of Marine Species (Worms); Society for the Management of Electronic Biodivirsity Data (SMEBD): Dublin, UK, 2014. [Google Scholar]
- Smith, V.; Georgiev, T.; Stoev, P.; Biserkov, J.; Miller, J.; Livermore, L.; Baker, E.; Mietchen, D.; Couvreur, T.L.; Mueller, G.; et al. Beyond dead trees: Integrating the scientific process in the biodiversity data journal. Biodivers. Data J. 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Scott, B.; Smith, V. Natural History Museum Data Portal. Available online: http://data.nhm.ac.uk (accessed on 14 December 2015).
- Smith, V.S.; Rycroft, S.; Scott, B.; Baker, E.; Livermore, L.; Heaton, A.; Bouton, K.; Koureas, D.N.; Roberts, D. Scratchpads 2.0: A virtual Research Environment Infrastructure for Biodiversity Data. Available online: http://scratchpads.eu (accessed on 14 December 2015).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glover, A.G.; Dahlgren, T.G.; Wiklund, H.; Mohrbeck, I.; Smith, C.R. An End-to-End DNA Taxonomy Methodology for Benthic Biodiversity Survey in the Clarion-Clipperton Zone, Central Pacific Abyss. J. Mar. Sci. Eng. 2016, 4, 2. https://doi.org/10.3390/jmse4010002
Glover AG, Dahlgren TG, Wiklund H, Mohrbeck I, Smith CR. An End-to-End DNA Taxonomy Methodology for Benthic Biodiversity Survey in the Clarion-Clipperton Zone, Central Pacific Abyss. Journal of Marine Science and Engineering. 2016; 4(1):2. https://doi.org/10.3390/jmse4010002
Chicago/Turabian StyleGlover, Adrian G., Thomas G. Dahlgren, Helena Wiklund, Inga Mohrbeck, and Craig R. Smith. 2016. "An End-to-End DNA Taxonomy Methodology for Benthic Biodiversity Survey in the Clarion-Clipperton Zone, Central Pacific Abyss" Journal of Marine Science and Engineering 4, no. 1: 2. https://doi.org/10.3390/jmse4010002
APA StyleGlover, A. G., Dahlgren, T. G., Wiklund, H., Mohrbeck, I., & Smith, C. R. (2016). An End-to-End DNA Taxonomy Methodology for Benthic Biodiversity Survey in the Clarion-Clipperton Zone, Central Pacific Abyss. Journal of Marine Science and Engineering, 4(1), 2. https://doi.org/10.3390/jmse4010002