
Effects of Exposure of Pink Shrimp, Farfantepenaeus duorarum, Larvae to Macondo Canyon 252 Crude Oil and the Corexit Dispersant


Abstract
:1. Introduction
2. Experimental Section
2.1. Animals
2.2. Solution Preparation
2.3. PAH Analysis
2.4. Acute Toxicity Bioassays
2.4.1. Survival (Determination of LC50 Values)
2.4.2. Behavioral Responses
2.5. Sub-Lethal Toxicity Bioassays
3. Results
3.1. PAH Analysis
3.2. Acute Toxicity Bioassays
3.2.1. Survival (Nominal LC50 Values)
3.2.2. Survival (Determined PAH and TPH LC50 Values)
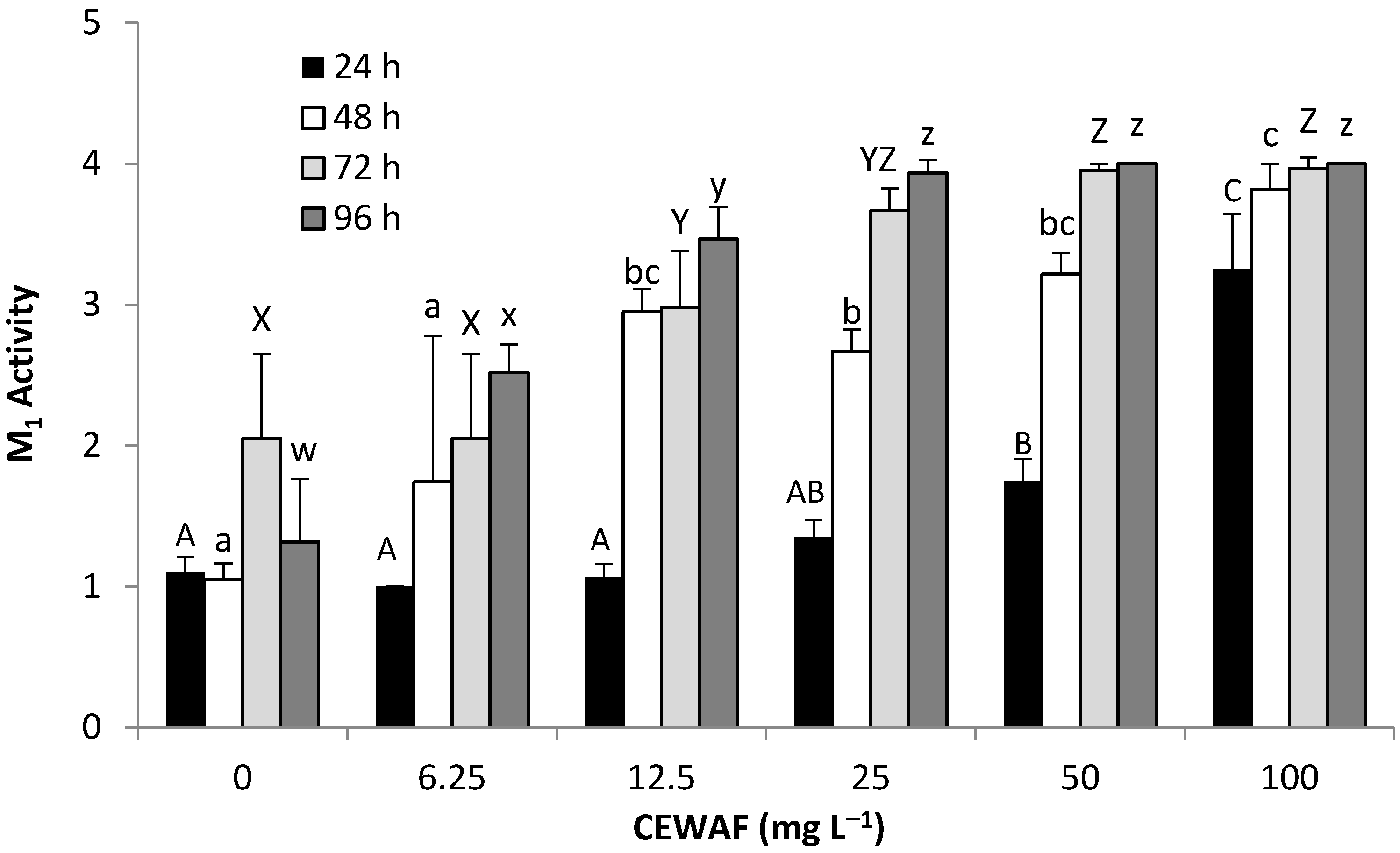
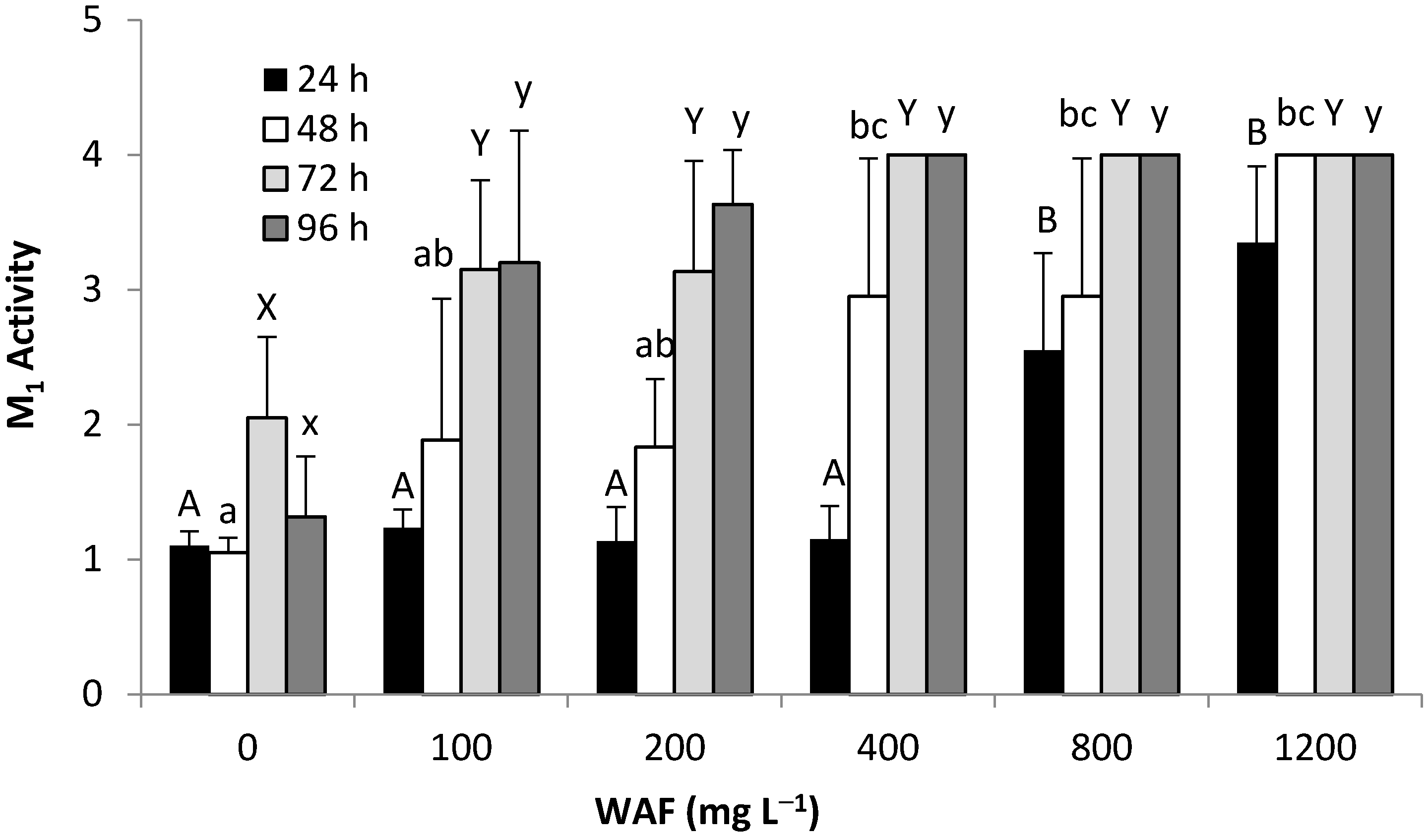
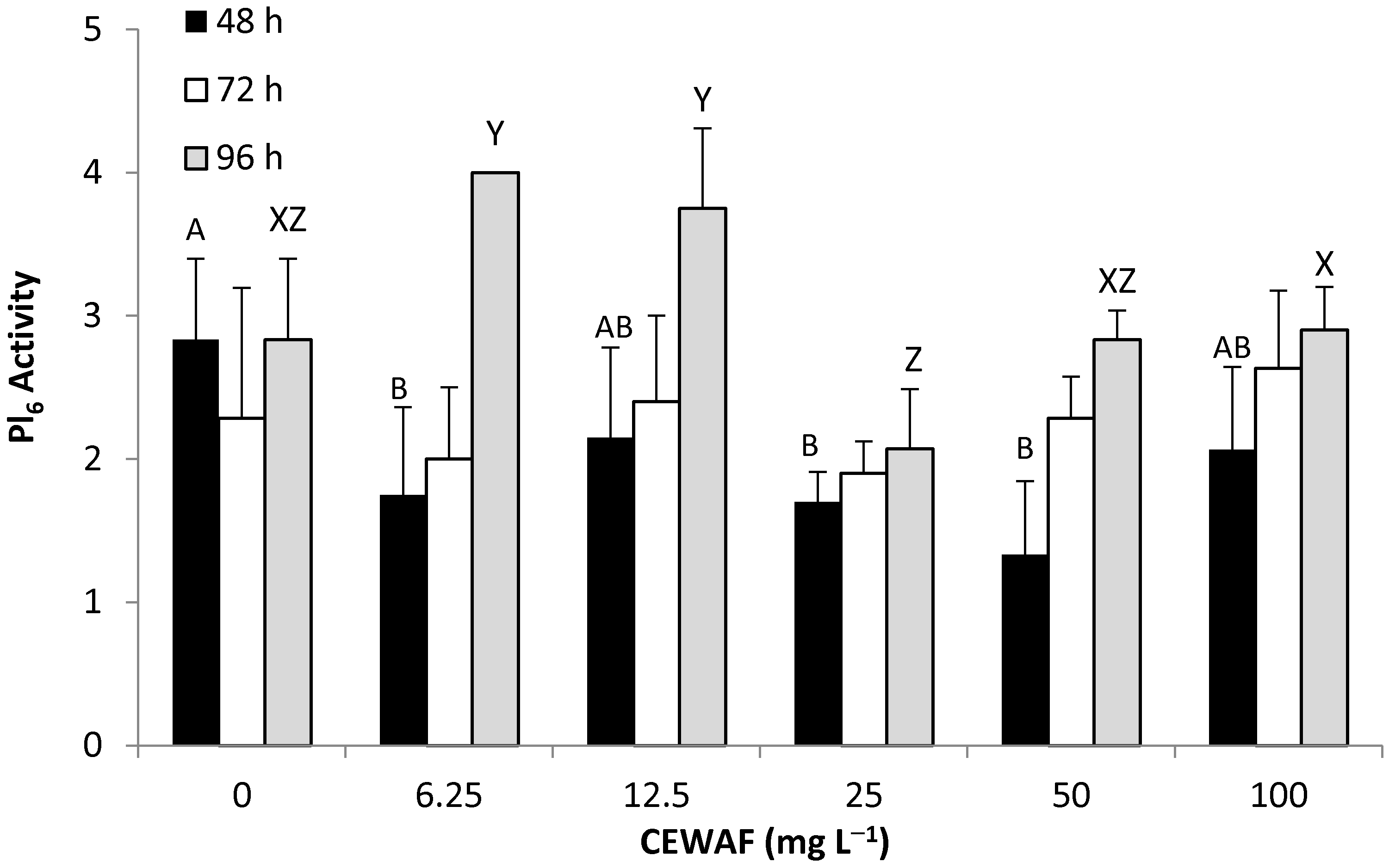
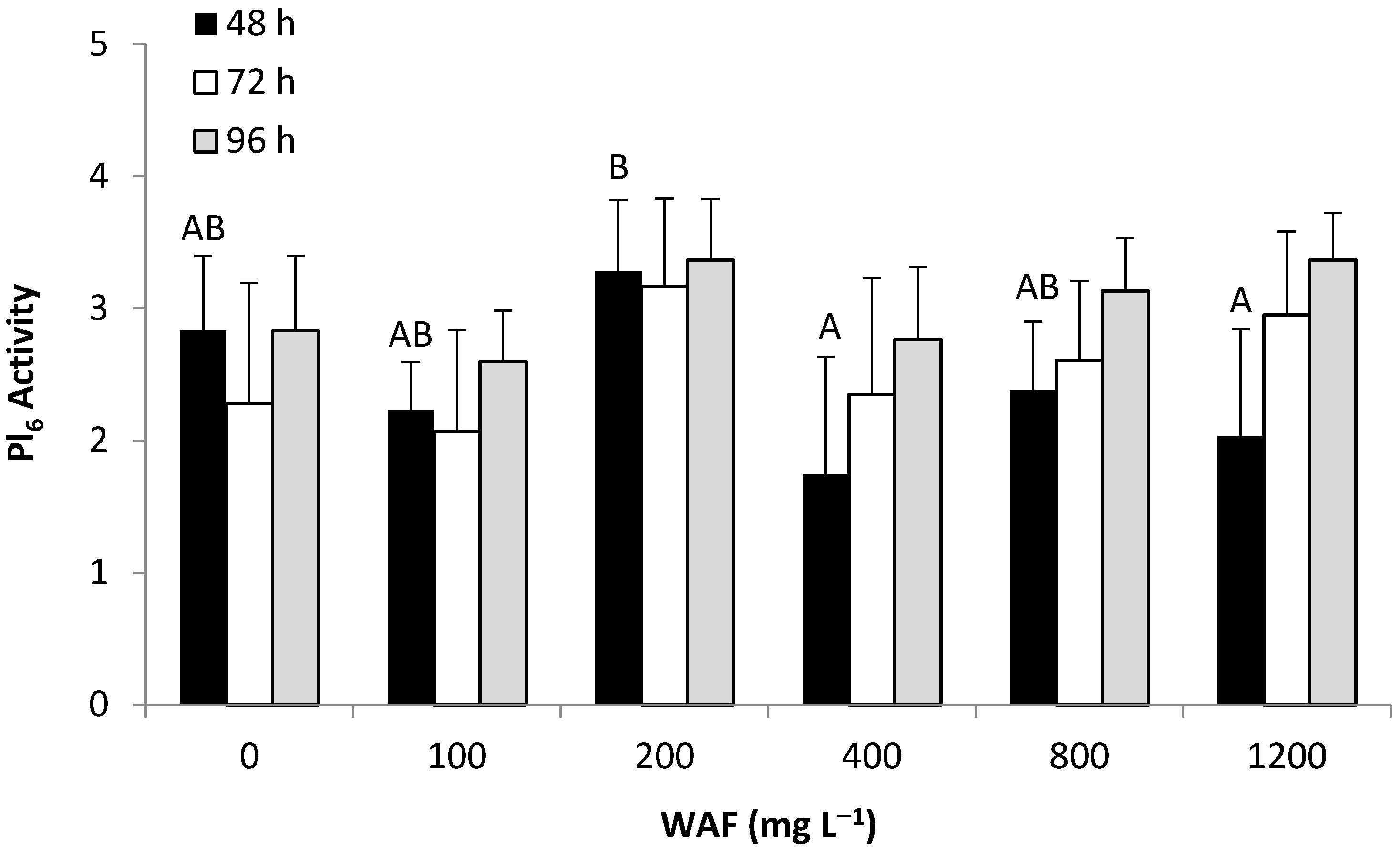
3.2.3. Behavioral Response—WAF & CEWAF (M1,Pl6)
3.2.4. Behavioral Response—CEWAF (N5, Z1, Z3, M2)
3.3. Long Term Sublethal Effects
3.3.1. Survival
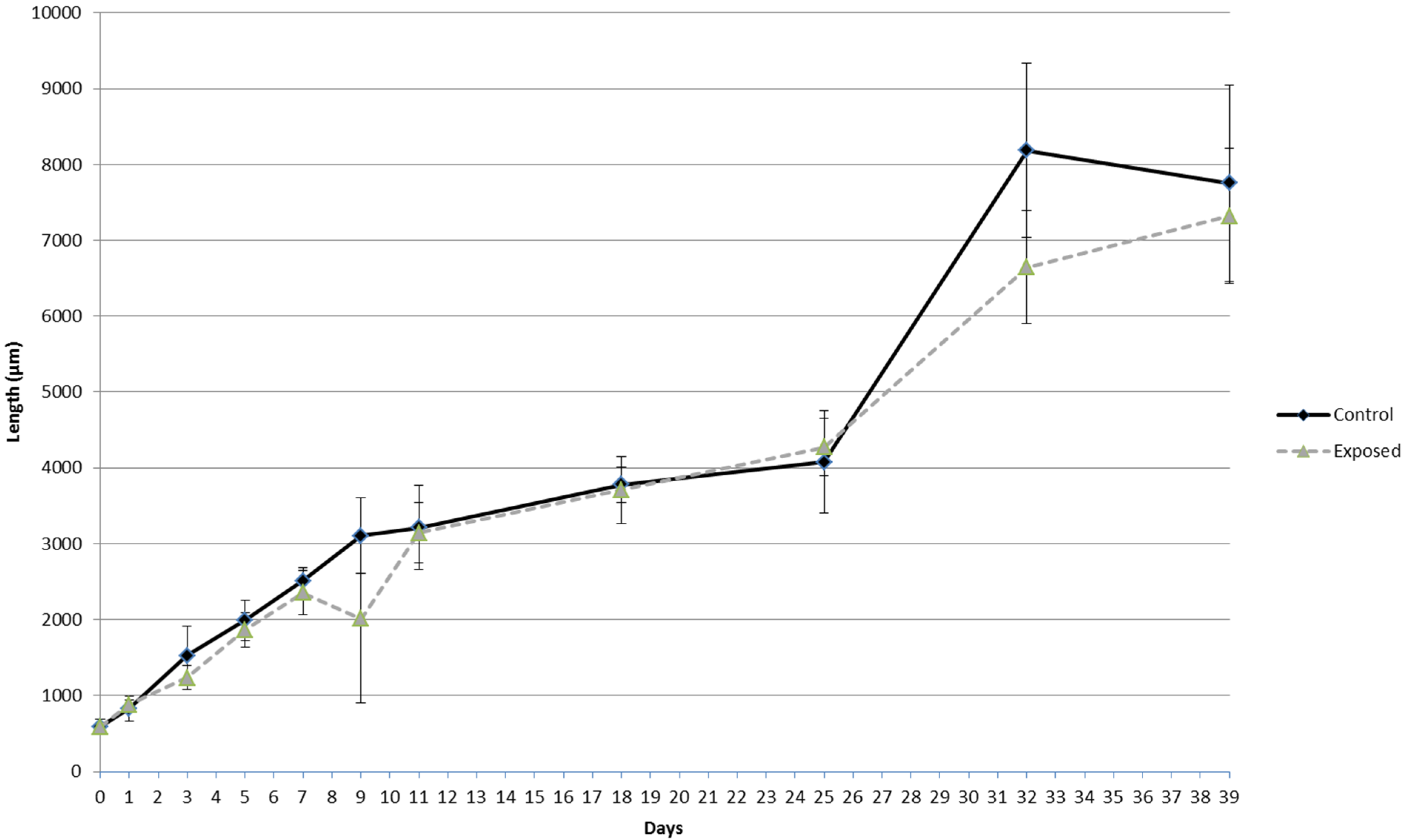
3.3.2. Growth
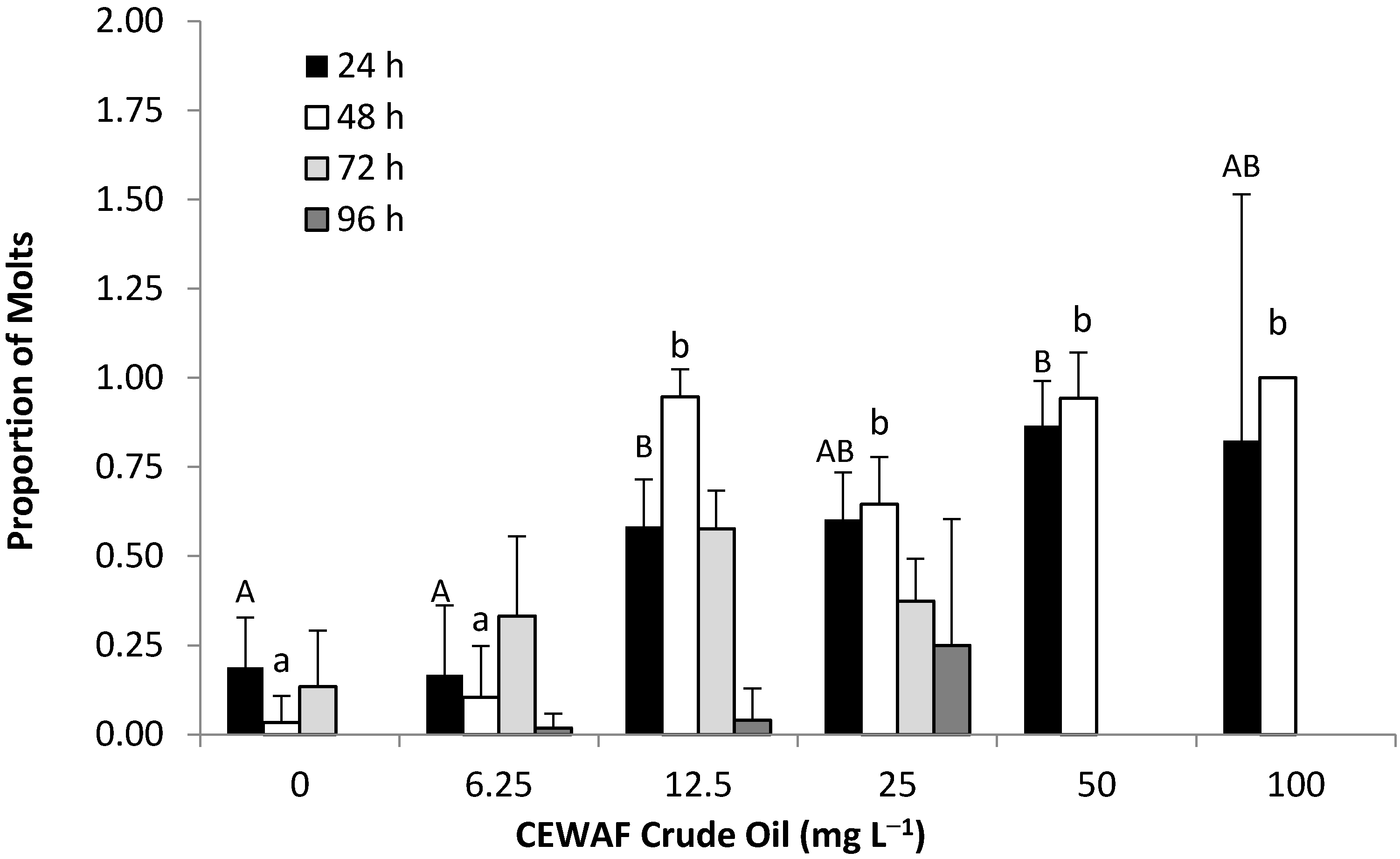
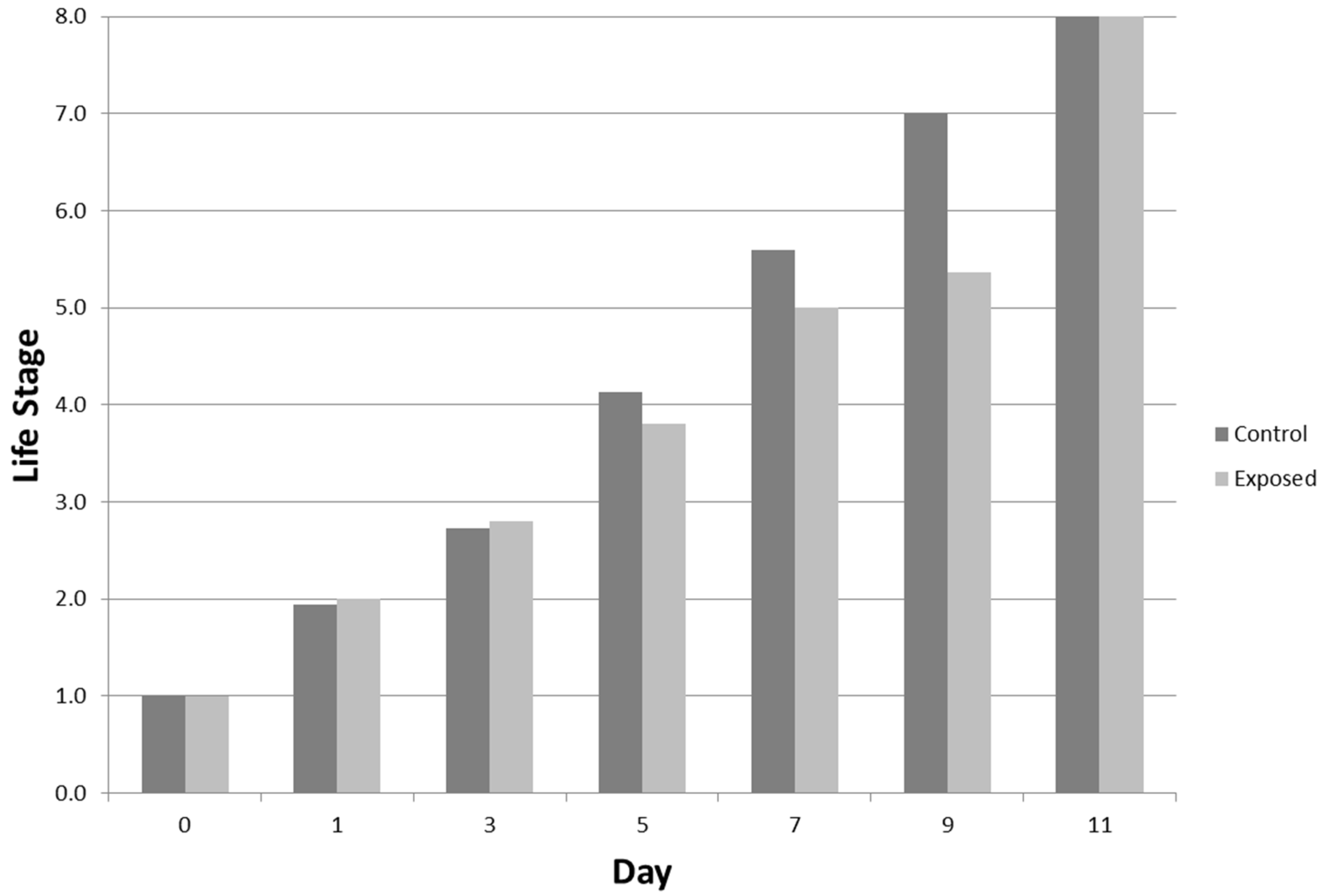
3.3.3. Developmental Stages
4. Discussion
4.1. Acute Toxicity Effects
4.1.1. Survival
4.1.2. Behavior
4.1.3. Molting
4.2. Sublethal Effects
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- National Marine Fisheries Service (NMFS). Fisheries of the United States 2003. In National Marine Fisheries Service, Office of Science and Technology; NMFS: Silver Spring, MD, USA, 2004. [Google Scholar]
- National Marine Fisheries Service (NMFS). Fisheries of the United States 2010. In National Marine Fisheries Service, Office of Science and Technology; NMFS: Silver Spring, MD, USA, 2011. [Google Scholar]
- Zimmerman, R.J.; Minello, T.J.; Rozas, L.P. Saltmarsh linkages to productivity of penaeid shrimps and blue crabs in the Northern Gulf of Mexico. In Concepts and Controversies in Tidal Marsh Ecology; Weinstein, M.P., Kreeger, D.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 293–314. [Google Scholar]
- Linder, M.J.; Anderson, W.W. Growth, Migration, Spawning and Size Distribution of Shrimp: Penaeus setiferus; US Government Printing Office: Washington, DC, USA, 1956; Volume 56, pp. 555–645. [Google Scholar]
- Renfro, W.C. Life History Stages of Gulf of Mexico Brown Shrimp; United States Bureau of Commercial Fisheries: Galveston, TX, USA, 1964; pp. 94–98. [Google Scholar]
- Cummings, W.C. Maturation and spawning of the pink shrimp, Penaeus duorarum Burkenroad. Trans. Am. Fish. Soc. 1961, 90, 462–468. [Google Scholar] [CrossRef]
- National Commission on the BP Deepwater Horizon Oil Spill and Offshore Drilling. Deep Water: The Gulf oil disaster and the future of offshore drilling. 2011; Report to the President. Available online: http://docs.lib.noaa.gov/noaa_documents/NOAA_related_docs/oil_spills/DWH_report-to-president.pdf (accessed on 16 November 2011). [Google Scholar]
- Kelly, T.R. Environmental health insights into the 2010 Deepwater Horizon (BP) oil blowout. Environ. Health Insights 2010, 4, 61–63. [Google Scholar] [CrossRef]
- Federal Interagency Solutions Group. Oil Budget Calculator: Deepwater Horizon, November 2010. Available online: http://www.restorethegulf.gov/sites/default/files/documents/pdf/OilBudgetCalc_Full_HQ-Print_111110.pdf (accessed on 16 November 2011).
- George Aryes, A.; Clark, J.R. Aquatic toxicity of two Corexit dispersants. Chemosphere 2000, 40, 897–906. [Google Scholar] [CrossRef]
- Fucik, K.W.; Carr, K.A.; Balcom, B.J. Toxicity of oil and dispersed oil to the eggs and larvae of seven marine fish and invertebrates from the Gulf of Mexico. In The Use of Chemicals in Oil Spill Response; ASTM STP 1252; Lane, P., Ed.; American Society for Testing and Materials: Philadelphia, PA, USA, 1995; pp. 135–171. [Google Scholar]
- Le Hir, M.; Hily, C. First observations in a high rocky-shore community after the Erika oil spill (December 1999, Brittany, France). Mar. Pollut. Bull. 2002, 44, 1243–1252. [Google Scholar] [CrossRef]
- Liu, B.; Romaire, R.P.; Delaune, R.D.; Lindau, C.W. Field investigation on the toxicity of Alaska North Slope crude oil (ANSC) and dispersed ANSC crude to Gulf killifish, Eastern oyster and white shrimp. Chemosphere 2006, 62, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Saco-Alvarez, L.; Bellas, J.; Nieto, O.; Bayona, J.M.; Albaiges, J.; Beiras, R. Toxicity and phototoxicity of water-accommodated fraction obtained from Prestige fuel oil and Marine fuel oil evaluated by marine bioassays. Sci. Total Environ. 2008, 394, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Laramore, S.; Krebs, W.; Garr, A. Effects of Macondo Canyon 252 oil (naturally and chemically dispersed) on larval Crassostrea virginica (Gmelin, 1791). J. Shell. Res. 2014, 33, 709–718. [Google Scholar] [CrossRef]
- Temara, A.; Gulec, I.; Holdway, D.A. Oil induced disruption of foraging behavior of the asteroid keystone predator, Coscinasterias muricata (Echinodermata). Mar. Biol. 1999, 133, 501–507. [Google Scholar] [CrossRef]
- Fernandez, N.; Cesar, A.; Salamanca, M.J.; DelValls, T.A. Toxicological characterization of the aqueous soluble phase of Prestige fuel oil using the sea urchin embryo bioassay. Ecotoxicology 2006, 15, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Beiras, R.; Saco Alvarez, L. Toxicity of seawater and sand affected by the Prestige fuel oil spill using bivalve and sea urchin embryogenesis bioassays. Water Air Soil Poll. 2006, 177, 457–466. [Google Scholar] [CrossRef]
- Martínez-Jerónimo, F.; Villaseñor, R.; Ríos, G.; Espinosa-Chavez, F. Toxicity of the crude oil water soluble fraction and kaolin-adsorbed crude oil on Daphnia magna (Crustacea: Anomopoda). Arch. Environ. Contam. Toxicol. 2005, 48, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Roth, A.M.F.; Baltz, D.M. Short-term effects of an oil spill on marsh edge fishes and decapod crustacean. Estuar. Coasts 2009, 32, 565–572. [Google Scholar] [CrossRef]
- Bechmann, R.K.; Larsen, B.K.; Taban, I.C.; Hellgren, L.I.; Moller, P.; Sanni, S. Chronic exposure of adults and embryos of Pandalus borealis to oil causes PAH accumulation, initiation of biomarker responses and an increase in larval mortality. Mar. Poll. Bull. 2010, 60, 2087–2098. [Google Scholar] [CrossRef] [PubMed]
- Rozas, L.P.; Minello, T.J.; Miles, M.S. Effect of Deepwater Horizon oil on growth rates of juvenile penaeid shrimps. Estuar. Coasts 2014, 37, 1403–1414. [Google Scholar] [CrossRef]
- Swedmark, M.; Granmo, A.; Kollberg, S. Effects of oil dispersants and oil emulsions on marine animals. Water Res. 1973, 7, 1649–1692. [Google Scholar] [CrossRef]
- Wells, P.G. The toxicity of oil dispersants to marine organisms: A current perspective. In Oil Spill Chemical Dispersants: Research, Experience and Recommendations; ASTM International: Philadelphia, PA, USA, 1984; pp. 177–202. [Google Scholar]
- Singer, M.M.; Aurand, D.; Coelho, G.; Bragin, G.E.; Clark, J.R.; Sowby, M.L.; Tjeerdema, R.S. Making, measuring and using water-accommodated fractions of petroleum for toxicity testing. In Proceedings of the 2001 International Oil Spill Conference, American Petroleum Institute, Tampa, FL, USA, 26–29 March 2001; pp. 1269–1274.
- Fuller, C.; Bonner, J. Comparative toxicity of oil, dispersant and dispersed oil to Texas marine species. In Proceedings of the 2001 International Oil Spill Conference, American Petroleum Institute, Tampa, FL, USA, 26–29 March 2001; pp. 1243–1248.
- Wu, R.S.S.; Lam, P.K.S.; Zou, B.S. Effects of two oil dispersants on phototaxis and swimming behavior of barnacle larvae. Hydrobiologia 1997, 352, 9–16. [Google Scholar] [CrossRef]
- Barron, M.G.; Carl, M.G.; Short, J.W.; Rice, S.D. Photoenhanced toxicity of aqueous phase and chemically dispersed weathered Alaska North Slope crude oil to Pacific herring eggs and larvae. Environ. Toxicol. Chem. 2003, 22, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Georgiades, E.T.; Holdway, D.E.; Brennan, S.E.; Butty, J.S.; Temera, A. The impact of oil derived products on the behavior and biochemistry of the eleven armed asteroid Coscinasterias muracata, Echinodermata. Mar. Environ. Res. 2003, 55, 257–276. [Google Scholar] [CrossRef]
- Lopez, B.A.; Lopez, D.A. Moulting frequency and behavioural responses to salinity and diesel oil in Austromegabalanus psittacus (Molina) (Cirripedia: Balanidae). Mar. Freshw. Behav. Physiol. 2005, 38, 249–258. [Google Scholar] [CrossRef]
- Greco, G.; Corra, C.; Garaventa, F.; Chelossi, E.; Faimali, M. Standardization of laboratory bioassays with Balanus amphitrite larvae for preliminary oil dispersants toxicological characterization. Chem. Ecol. 2006, 22, 163–172. [Google Scholar] [CrossRef]
- Clark, J.R.; Bragin, G.E.; Febbo, R.J.; Letinski, D.J. Toxicity of physically and chemically dispersed oils under continuous and environmentally realistic exposure conditions: Applicability to dispersant use decisions in spill response planning. In Proceedings of the 2001 International Oil Spill Conference, American Petroleum Institute, Tampa, FL, USA, 26–29 March 2001; pp. 1249–1255.
- Barron, M.G.; Ka'aihue, L. Critical evaluation of CROSERF test methods for oil dispersant toxicity testing under subarctic conditions. Mar. Pollut. Bull. 2003, 46, 1191–1199. [Google Scholar] [CrossRef]
- National Research Council (NRC). Committee on understanding oil spill dispersants. In Oil Spill Dispersants Efficacy and Effects; National Academies Press: Washington, DC, USA, 2005; p. 400. [Google Scholar]
- Fincas, M. A Review of Literature Related to Oil Spill Dispersants 1997–2008; Prince William Sound Regional Citizens Advisory Council: Anchorage, AK, USA, 2008; p. 155. [Google Scholar]
- Trudel, K. Environmental risks and trade-offs in Prince William Sound. In Proceedings of the Dispersant Application in Alaska: A Technical Update, Anchorage, AK, USA, 1998; pp. 159–188.
- Treece, G.D.; Fox, J.M. Design, Operation and Training Manual for an Intensive Culture Shrimp Hatchery; Publication TAMU-SG-93–505; Texas A&M University, Sea Grant College Program: Galveeston, TX, USA, 1993; p. 187. [Google Scholar]
- Cohen, A.M.; Gagnon, M.M.; Nugegoda, D. Alterations of metabolic enzymes in Australian bass, Macquaria novemaculeata, after exposure to petroleum hydrocarbons. Arch. Environ. Contam. Toxicol. 2005, 49, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Gulec, I.; Leonard, B.; Holdway, D.A. Oil and dispersed oil toxicity to amphipods and snails. Spill Sci. Technol. Bull. 1997, 4, 1–6. [Google Scholar] [CrossRef]
- Fuller, C.; Bonner, J.; Page, C.; Ernest, A.; McDonald, T.; McDonald, S. Comparative toxicity of oil, dispersant and oil plus dispersant to several marine species. Environ. Toxicol. Chem. 2004, 23, 2941–2949. [Google Scholar] [CrossRef] [PubMed]
- Perkins, R.A.; Rhoton, S.; Behr-Andres, C. Toxicity of dispersed and undispersed, fresh and weathered oil to larvae of a cold water species, Tanner crab (C. bairdi) and standard warm water test species. Cold Reg. Sci. Technol. 2003, 36, 129–140. [Google Scholar] [CrossRef]
- Rhoton, S.; Perkins, R.A.; Braddock, J.F.; Behr-Andres, C. A cold weather species response to chemically dispersed fresh and weathered Alaska North Slope crude oil. In Proceedings of the 2001 International Oil Spill Conference, American Petroleum Institute, Tampa, FL, USA, 26–29 March 2001; pp. 1231–1236.
- Wetzel, D.L.; Van Fleet, E.S. Cooperative studies on the toxicity of dispersants and dispersed oil to marine organisms: A 3 year Florida study. In Proceedings of the 2001 International Oil Spill Conference, American Petroleum Institute, Tampa, FL, USA, 26–29 March 2001; pp. 1237–1241.
- Singer, M.M.; George, S.; Lee, I.; Jacobson, S.; Weetman, L.L.; Blondina, G.; Tjeerdema, R.S.; Aurand, D.; Sowby, M.L. Effects of dispersant treatment on the acute aquatic toxicity of petroleum hydrocarbons. Environ. Contam. Toxicol. 1998, 34, 177–187. [Google Scholar] [CrossRef]
- Boehm, P.D.; Cook, L.L.; Murray, K.J. Aromatic hydrocarbon concentrations in seawater: Deepwater Horizon oil spill. In Proceedings of the 2011 International Oil Spill Conference, American Petroleum Institute, Portland, OR, USA, 23–26 May 2011; Volume 1. Abstract 371.
- Echols, B.S.; Smith, A.J.; Gardinali, P.R.; Rand, G.M. Acute aquatic toxicity studies of Gulf of Mexico water samples collected following the Deepwater Horizon incident (2 May 2010 to 11 December 2010). Chemosphere 2015, 120, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Allan, S.E.; Smith, B.W.; Anderson, K.A. Impact of the Deepwater Horizon oil spill on bioavailable polycylic aromatic hydrocarbons in Gulf of Mexico coastal waters. Environ. Sci. Techno. 2012, 46, 2033–2039. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, A. Aquatic behavioral ecotoxicology—Prospects and limitations. Hum. Ecol. Risk Assess. 2007, 13, 481–491. [Google Scholar] [CrossRef]
- Eisler, R.; Carney, G.C.; Lockwood, A.P.M.; Perkins, E.J. Behavioural responses of marine poikilotherms to pollutants (and discussion). Philos. Trans. R. Soc. Lond. B Biol. Sci. 1979, 286, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Takashashi, F.T.; Kittredge, J.S. Sublethal effects of the water soluble component of oil: chemical communication in the marine environment. In The Microbial Degradation of Oil Pollutants; Ahearn, D.G., and Mers, S.P., Eds.; Louisiana State University: Baton Rouge, LA, USA, 1973; pp. 259–264. [Google Scholar]
- McCain, B.B.; Malins, D.C. Effects of petroleum hydrocarbons on selected demersal fishes and crustaceans. In Sediment Toxicity Assessment; Burton, G.A., Ed.; Lewis Publishers: Boca Raton, FL, USA, 1992; pp. 315–325. [Google Scholar]
- Jensen, L.K.; Carroll, J. Experimental studies of reproduction and feeding for two Artic dwelling Calanus species exposed to crude oil. Aquatic. Biol. 2010, 10, 261–271. [Google Scholar] [CrossRef]
- Bigford, TE. Effects of oil on behavioral response to light, pressure and gravity in larvae of the rock crab Cancer irroratus. Mar. Biol. 1977, 43, 137–148. [Google Scholar] [CrossRef]
- Brodersen, C. Rapid narcosis and delayed mortality in larvae of king crabs and kelp shrimp exposed to the water soluble fraction of crude oil. Mar. Environ. Res. 1987, 22, 233–239. [Google Scholar] [CrossRef]
- Rice, S.D.; Short, J.W.; Brodersen, C.C.; Mecklenburg, T.A.; Moles, D.A.; Misch, C.J.; Cheatham, D.L.; Karinen, J.F. Acute toxicity and uptake-depuration studies with Cook Inlet crude oil, Prudhoe Bay crude oil, No. 2 fuel oil, and several subarctic marine organisms. In Northwest Fisheries Center Auke Bay Fisheries Laboratory, Processed Report; National Marine Fisheries Service, NOAA: Juneau, AK, USA, 1976; p. 90. [Google Scholar]
- Van Wezel, A.P.; Opperhuizen, A. Narcosis due to environmental pollutants in aquatic organisms—Residue-based toxicity, mechanisms and membrane burdens. Crit. Rev. Toxicol. 1995, 25, 255–279. [Google Scholar] [CrossRef] [PubMed]
- Barron, M.G.; Carls, M.G.; Heintz, R.; Rice, S.D. Evaluation of fish early life stage toxicity models of chronic embryonic exposures to complex polycyclic aromatic hydrocarbon mixtures. Toxicol. Sci. 2004, 78, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Weis, J.S.; Cristini, A.; Rao, K.R. Effects of pollutants on molting and regeneration in Crustacea. Amer. Zool. 1992, 32, 495–500. [Google Scholar] [CrossRef]
- Johns, D.M.; Pechenik, J.A. Influence of water accommodated fraction of No. 2 fuel oil on energetics of Cancer irroratus larvae. Mar. Biol. 1980, 55, 247–254. [Google Scholar] [CrossRef]
- Karinen, J.F.; Rice, S.D. Effects of Prudhoe bay crude oil on molting tanner crabs, Chionoecetes bairdi. Mar. Fish. Rev. 1974, 36, 31–37. [Google Scholar]
- Cantelmo, A.; Lazell, R.; Mantel, L. The effects of benzene on molting and limb regeneration in juvenile Callinectes sapidus. Mar. Bio. Lett. 1981, 2, 333–343. [Google Scholar]
- Cucci, T.L.; Epifanio, C.E. Long term effects of water soluble fractions of Kuwait crude oil on the larval and juvenile development of the mud crab Eurypanopeus depressus. Mar. Biol. 1979, 55, 215–220. [Google Scholar] [CrossRef]
- Laughlin, R.B., Jr.; Ng, J.; Guard, H.E. Hormesis: A response to low environmental concentrations of petroleum hydrocarbons. Science 1981, 211, 705–707. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, R.B. Jr.; Young, L.G.L.; Neff, J.M. A long term study of the effects of water soluble fractions of No. 2 fuel oil on the survival, development rate and growth of the mud crab Rhithropanopeus harrisii. Mar. Biol. 1978, 47, 87–95. [Google Scholar] [CrossRef]
- Bang, H.W.; Lee, W.; Kwak, L.S. Detecting points as developmental delay based on the life-history development and urosome deformity of the harpacticoid copepod, Tigriopus japonicus sensu lato, following exposure to benzo(a)pyrene. Chemosphere 2009, 76, 1435–1439. [Google Scholar] [CrossRef] [PubMed]
- Worboys, M.A.; Leung, K.M.Y.; Grist, E.P.M.; Crane, M. Time should be considered in developmental ecotoxicity test. Mar. Poll. Bull. 2002, 45, 92–99. [Google Scholar] [CrossRef]
- Laughlin, R.B., Jr.; Neff, J.M. Ontogeny of respiratory and growth responses of larval mud crabs Rhithropanopeus harrisii exposed to different temperatures, salinities and naphthalene concentrations. Mar. Ecol. Prog. Ser. 1981, 5, 319–332. [Google Scholar] [CrossRef]
- Gunderson, D.T.; Kristanto, S.W.; Curtis, L.R.; AL-Yakoob, S.N.; Metwally, M.; Al-Aljimi, D. Subacute toxicity of the water soluble fractions of Kuwait crude oil and partially combusted crude oil on Menidia beryllina and Palaemontes pugio. Arch. Environ. Contam. Toxicol. 1996, 31, 1–8. [Google Scholar] [CrossRef]
- Van der Ham, J.L.; de Musert, K. Abundance and size of Gulf shrimp in Louisiana’s coastal areas following the Deepwater Horizon oil spill. PLOS ONE 2014, 9, e108884. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Compounds | C rings | 2 ppt CEWAF | 2ppt WAF |
|---|---|---|---|
| µg L−1 | µg L−1 | ||
| Napthalene (C0-C4) | 2 | 925.96 | 377.66 |
| Acenaphthylene | 2 | 6.40 | 0.05 |
| Acenaphthene | 2 | 0.61 | 0.67 |
| Fluorene (C0-C4) | 3 | 102.92 | 14.65 |
| Anthracene (C0-C4) | 3 | 235.38 | 30.14 |
| Phenanthrene | 3 | 34.79 | 8.36 |
| Fluoranthene | 3 | 1.53 | 0.18 |
| Chrysene (C0-C4) | 4 | 32.6 | 4.02 |
| Pyrene (C0-C4) | 4 | 61.5 | 6.71 |
| Benzo[A]anthracence | 4 | 0.21 | 0.14 |
| Napthobenzothiophene (C0-C4) | 4 | 1.32 | 0.16 |
| Dibenzothiophene (C0-C4) | 5 | 5.47 | 5.6 |
| Benzo[B]fluorene | 5 | 0.72 | 0.09 |
| Benzo[B]fluoranthene | 5 | 0.00 | 0.07 |
| Benzo[K]fluoranthene | 5 | 0.43 | 0.00 |
| Benzo[E]pyrene | 5 | 0.61 | 0.11 |
| Benzo[A]pyrene | 5 | 0.00 | 0.02 |
| Perylene | 5 | 0.77 | 0.13 |
| Dibenzo[A,H]anthracene | 5 | 0.00 | 0.01 |
| Indeno[1,2,3-Cd]pyrene | 6 | 0.01 | 0.00 |
| Benzo[G,H,I]perylene | 6 | 0.00 | 0.02 |
| Total PAH in µg L−1 | 1428.64 | 451.92 | |
| Total Petroleum Hydrocarbon C9-C42 in µg L−1 | 62,613.50 | 2466.57 |
| Time | WAF LC50 | PAH | TPH | CEWAF LC50 | PAH | TPH | Corexit LC50 |
|---|---|---|---|---|---|---|---|
| 24 h | |||||||
| Nauplii | >100 | NC | NC | 81.5 (75.3, 88.1) | 58 | 2,551 | 33.3 (31.8, 34.9) |
| Zoea 1 | 67.4 (39, 100) | 15 | 83 | 15.4 (11.6, 20.4) | 11 | 470 | 3.1 (0.7, 13.7) |
| Mysis 1 | >100 | NC | NC | 84.6 (74.9, 95.7) | 60 | 2,649 | 20.9 (19.2, 22.7) |
| PL 6 | >400 | NC | NC | 99.7 (77.3, 100) | 71 | 3,121 | 28.4 (24.2, 33.3) |
| 48 h | |||||||
| Nauplii | >100 | NC | NC | 41.5 (36.3, 47.5) | 30 | 1,299 | 18.6 (16.8, 20.5) |
| Zoea 1 | 25.5 (22.5, 28.9) | 6 | 31 | ND | NC | NC | <2.5 |
| Mysis 1 | >100 | NC | NC | 47.4 (41.3, 54.3) | 34 | 1,484 | 18.3 (16.1, 20.8) |
| PL 6 | >400 | NC | NC | 70.4 (56.4, 87.9) | 50 | 2,204 | 26.5 (22.4, 31.3) |
| 72 h | |||||||
| Nauplii | ND | NC | NC | ND | NC | NC | ND |
| Zoea 1 | 21.2 (17.7, 25.5) | 5 | 26 | ND | NC | NC | ND |
| Mysis 1 | >100 | NC | NC | 31.9 (28.6, 35.7) | 23 | 999 | 8.3 (6.8, 10.1) |
| PL 6 | >400 | NC | NC | 49.2 (40, 60.5) | 35 | 1,002 | 22.4 (20.8, 23.9) |
| 96 h | |||||||
| Nauplii | ND | NC | NC | ND | NC | NC | ND |
| Zoea 1 | 23.3 (20.9, 26) | 5 | 29 | ND | NC | NC | ND |
| Mysis 1 | 29.7 | 7 | 37 | 8.5 (7.1, 10.1) | 6 | 266 | 2.6 (2.2, 3.0) |
| PL 6 | >400 | NC | NC | 44 (36.5, 53.2) | 31 | 1,377 | 22.5 (21.4, 23.8) |
| Larval Stage | Time (h) | Concentration (mg L−1) | Activity Level (1–4) ± S.D. | Feeding (1–4) ± S.D. | % molts | Phototaxic (1–3) ± S.D. | Metamorphosis |
|---|---|---|---|---|---|---|---|
| N5 | 24 | 0 | 1.03±0.05 a | 1.0 ± 0.0 a | - | 1.02 ± 0.05 a | N5-Z1 |
| 12.5 | 2.13 ± 0.05 b | 3.33 ± 0.5 b | - | 2.13 ± 0.05 b | N5-Z1 | ||
| 25 | 2.35 ± 0.35 bc | 3.67 ± 0.5 b | - | 2.27 ± 0.41 b | N5-Z1 | ||
| 50 | 2.28 ± 0.14 bc | 4.0 ± 0.0 b | - | 2.28 ± 0.14 b | N5-Z1 | ||
| 100 | 2.98 ± 0.04 c | 4.0 ± 0.0 b | - | 3.0 ± 0.0 c | N5-Z1 | ||
| 48 | 0 | 1.33 ± 0.52 a | - | - | 1.5 ± 0.58 a | Z1-Z2 | |
| 12.5 | 2.17 ± 0.98 a | - | - | 2.25 ± 0.29 b | Z1-Z2 | ||
| 25 | 2.23 ± 0.74 a | - | - | 2.34 ± 0.45 bc | Z1-Z2 | ||
| 50 | 3.19 ± 0.95 ab | - | - | 2.53 ± 0.67 bc | Z1-Z2 | ||
| 100 | 3.83 ± 0.41 b | - | - | 3.0 ± 0.0 c | Z1 | ||
| Z1 | 24 | 0 | 1.67 ± 0.82 a | - | 4% | 1.67 ± 0.82 a | Z1-Z2 (4:1) |
| 3.125 | 2.29 ± 0.56 ab | - | 12% | 2.33 ± 0.58 ab | Z1 | ||
| 6.25 | 2.54 ± 0.56 b | - | 21% | 2.63 ± 0.38 b | Z1 | ||
| 12.5 | 2.92 ± 0.13 bc | - | 21% | 2.92 ± 0.13 bc | Z1 | ||
| 25 | 3.0 ± 0.0 c | - | 21% | 3.0 ± 0.0 c | Z1 | ||
| 48 | 0 | 2.0 ± 1.55 | - | - | 1.67 ± 1.03 a | Z1-Z2 | |
| 3.125 | 2.17 ± 1.47 | - | - | 1.83 ± 0.98 a | Z1-Z2 | ||
| 6.25 | 3.0 ± 1.55 | - | - | 2.5 ± 0.77 ab | Z1-Z2 | ||
| 12.5 | 3.17 ± 1.33 | - | - | 2.5 ± 0.84 ab | Z1-Z2 | ||
| 25 | 4.0 ± 0.0 | - | - | 3.0 ± 0.0 b | - | ||
| 72 | 0 | 2.5 ± 1.64 a | - | - | 2.0 ± 1.1 | Z2 | |
| 3.125 | 2.75 ± 1.47 a | - | - | 2.33 ± 1.03 | Z1-Z2 | ||
| 6.25 | 3.75 ± 0.61 ab | - | - | 2.75 ± 0.61 | Z1-Z2 | ||
| 12.5 | 4.0 ± 0.0 b | - | - | 3.0 ± 0.0 | - | ||
| 25 | 4.0 ± 0.0 b | - | - | 3.0 ± 0.0 | - | ||
| Z3 | 24 | 0 | 1.0 ± 0.0 a | 1.0 ± 0.0 | 0.80% | 1.0 ± 0.0 a | - |
| 3.125 | 1.0 ± 0.0 a | 1.0 ± 0.0 | 1.60% | 1.0 ± 0.0 a | - | ||
| 6.25 | 1.0 ± 0.0 a | 1.0 ± 0.0 | 0% | 1.0 ± 0.0 a | - | ||
| 12.5 | 2.0 ± 0.0 b | 1.0 ± 0.0 | 2.80% | 3.0 ± 0.0 b | - | ||
| 25 | 2.0 ± 0.0 b | 1.0 ± 0.0 | 1.60% | 3.0 ± 0.0 b | - | ||
| 48 | 0 | 1.02 ± 0.04 | 1.0 ± 0.0 | 9.1% | - | Z3-M1 (3:2) | |
| 3.125 | 1.0 ± 0.0 | 1.0 ± 0.0 | 15% | - | Z3-M1 (1:4) | ||
| 6.25 | 1.0 ± 0.0 | 1.0 ± 0.0 | 19.0% | - | Z3-M1 (1:4) | ||
| 12.5 | 1.0 ± 0.0 | 1.0 ± 0.0 | 13.3% | - | M1 | ||
| 25 | 1.02 ± 0.04 | 1.0 ± 0.0 | 14.1% | - | Z3-M1 (2:3) | ||
| 72 | 0 | 1.13 ± 0.31 | - | 0% | 1.29 ± 0.71 ab | - | |
| 3.125 | 1.7 ± 1.1 | - | 0% | 1.7 ± 1.1 ab | - | ||
| 6.25 | 1.03 ± 0.05 | - | 0% | 1.03 ± 0.05 a | - | ||
| 12.5 | 1.12 ± 0.12 | - | 0% | 2.39 ± 0.47 b | - | ||
| 25 | 1.07 ± 0.05 | - | 1.60% | 2.03 ± 0.03 b | - | ||
| M2 | 24 | 0 | 1.0 ± 0.0 a | 1.0 ± 0.0 a | 4% | 1.0 ± 0.0 a | M2 |
| 12.5 | 1.2 ± 0.45 a | 1.0 ± 0.0 a | 1% | 1.0 ± 0.0 a | M2 | ||
| 25 | 1.84 ± 0.19 b | 2.2 ± 0.45 b | 28% | 3.0 ± 0.0 b | M2 | ||
| 50 | 1.9 ± 0.12 b | 3.6 ± 0.55 c | 37% | 3.0 ± 0.0 b | M2 | ||
| 100 | 2.0 ± 0.0 b | 4.0 ± 0.0 c | 51% | 3.0 ± 0.0 b | M2 | ||
| 48 | 0 | 1.0 ± 0.0 a | 1.0 ± 0.0 a | 2% | 1.0 ± 0.0 a | - | |
| 12.5 | 1.2 ± 0.45 a | 1.0 ± 0.0 a | 4% | 2.0 ± 0.0 b | - | ||
| 25 | 1.35 ± 0.41 a | 2.0 ± 0.0 b | 28% | 2.0 ± 0.0 b | - | ||
| 50 | 1.98 ± 0.08 b | 2.6 ± 0.55 bc | 2% | 3.0 ± 0.0 c | - | ||
| 100 | 2.28 ± 0.04 b | 3.4 ± 0.55 c | 0% | 3.0 ± 0.0 c | - | ||
| 72 | 0 | 1.8 ± 1.1 a | 1.0 ± 0.0 a | 0% | 1.0 ± 0.0 a | M3 | |
| 12.5 | 2.29 ± 0.40 a | 1.0 ± 0.0 a | 0% | 2.2 ± 0.45 b | - | ||
| 25 | 2.6 ± 0.55 ab | 3.2 ± 1.1 b | 3% | 2.8 ± 0.45 bc | - | ||
| 50 | 3.0 ± 0.0 b | 4.0 ± 0.0 b | 0% | 3.0 ± 0.0 c | - | ||
| 100 | 3.0 ± 0.0 b | 4.0 ± 0.0 b | 0% | 3.0 ± 0.0 c | - |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laramore, S.; Krebs, W.; Garr, A. Effects of Exposure of Pink Shrimp, Farfantepenaeus duorarum, Larvae to Macondo Canyon 252 Crude Oil and the Corexit Dispersant. J. Mar. Sci. Eng. 2016, 4, 24. https://doi.org/10.3390/jmse4010024
Laramore S, Krebs W, Garr A. Effects of Exposure of Pink Shrimp, Farfantepenaeus duorarum, Larvae to Macondo Canyon 252 Crude Oil and the Corexit Dispersant. Journal of Marine Science and Engineering. 2016; 4(1):24. https://doi.org/10.3390/jmse4010024
Chicago/Turabian StyleLaramore, Susan, William Krebs, and Amber Garr. 2016. "Effects of Exposure of Pink Shrimp, Farfantepenaeus duorarum, Larvae to Macondo Canyon 252 Crude Oil and the Corexit Dispersant" Journal of Marine Science and Engineering 4, no. 1: 24. https://doi.org/10.3390/jmse4010024
APA StyleLaramore, S., Krebs, W., & Garr, A. (2016). Effects of Exposure of Pink Shrimp, Farfantepenaeus duorarum, Larvae to Macondo Canyon 252 Crude Oil and the Corexit Dispersant. Journal of Marine Science and Engineering, 4(1), 24. https://doi.org/10.3390/jmse4010024