Harmful Fouling Communities on Fish Farms in the SW Mediterranean Sea: Composition, Growth and Reproductive Periods

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field and Laboratory Methods
2.3. Data Analysis
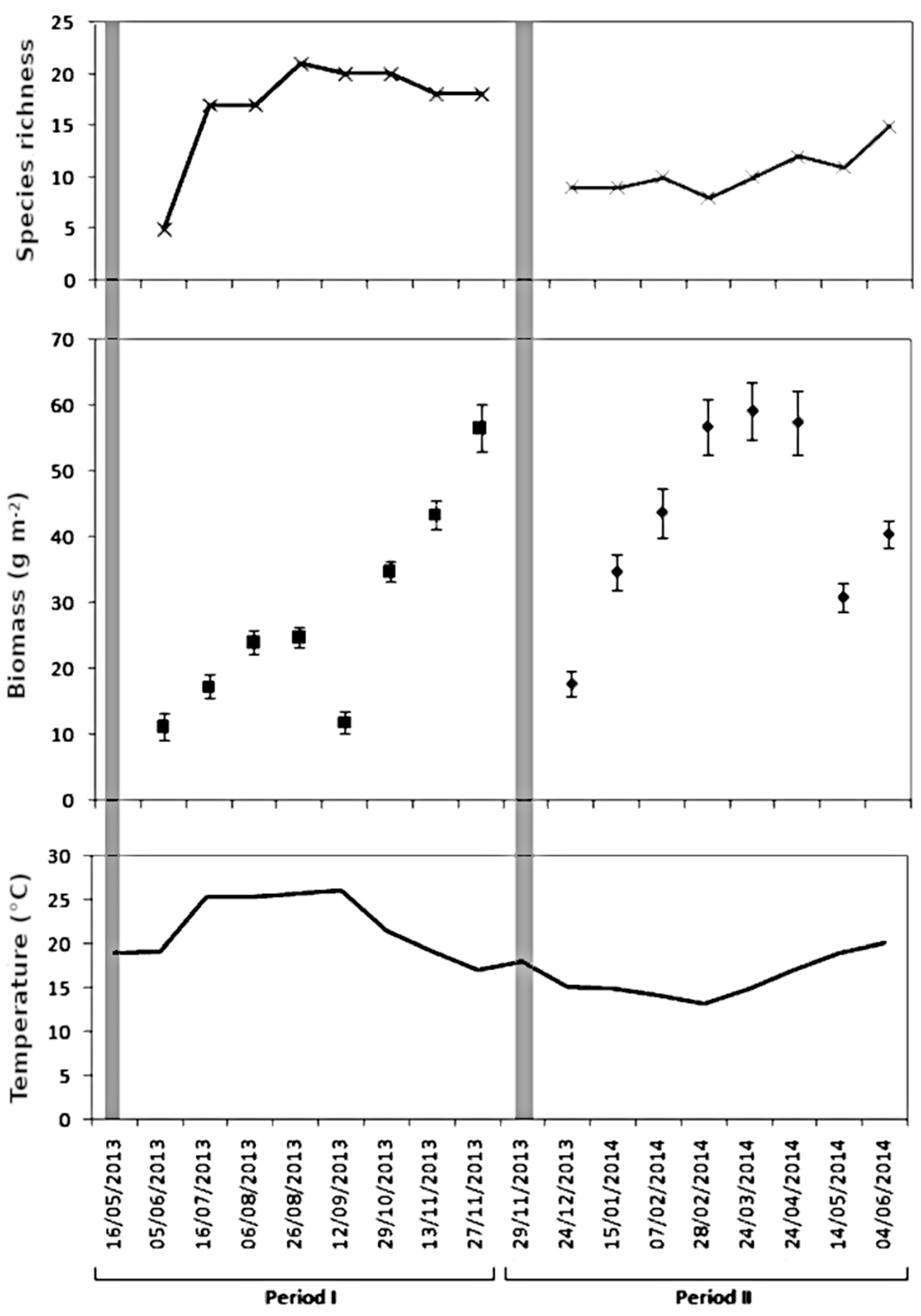
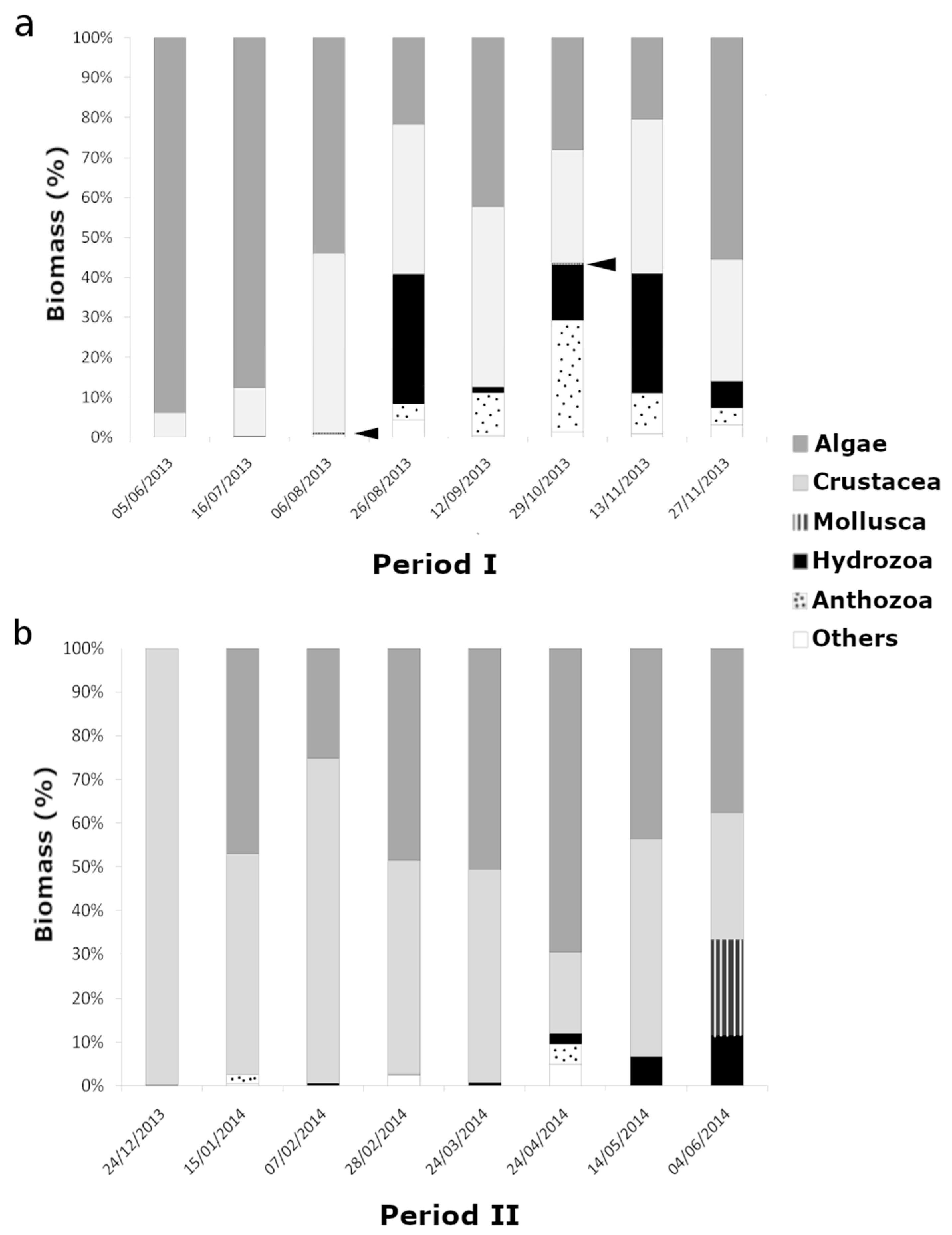
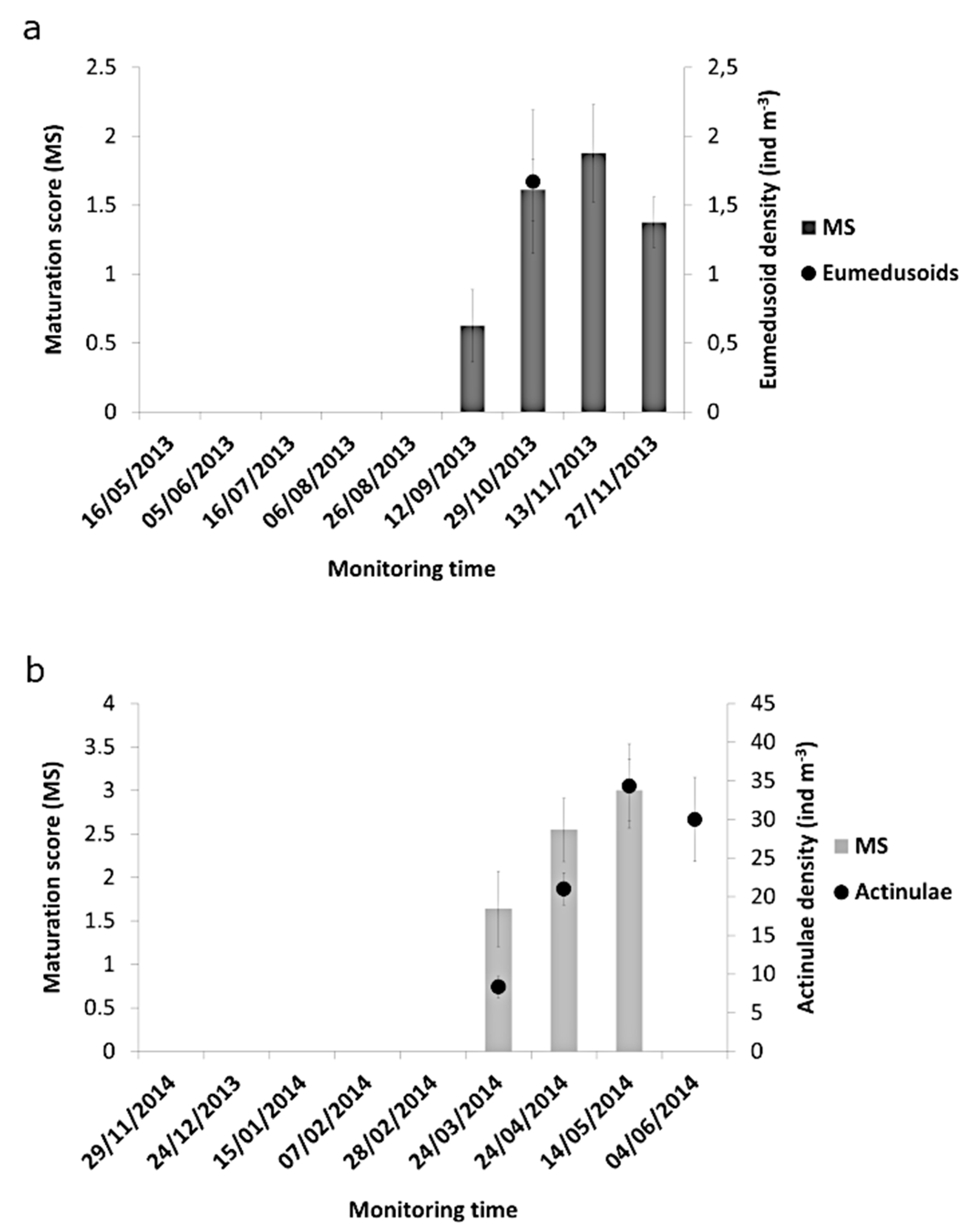
3. Results
3.1. Species Richness, Biomass and Community Composition
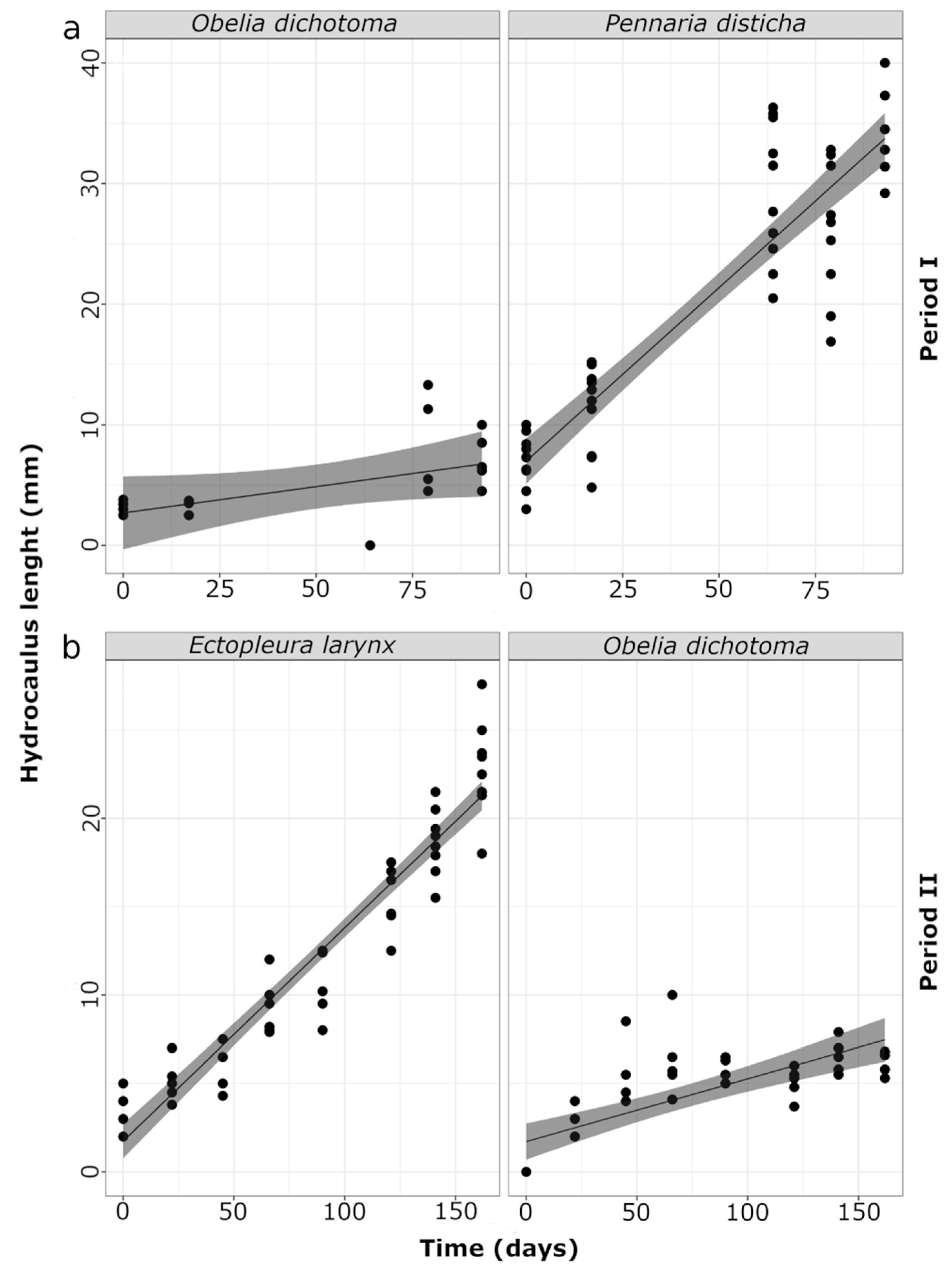
3.2. Hydroids and Colony Growth Rates
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethical Statement
References
- Bi, C.-W.; Zhao, Y.-P.; Dong, G.-H.; Wu, Z.-M.; Zhang, Y.; Xu, T.-J. Drag on and flow through the hydroid-fouled nets in currents. Ocean Eng. 2018, 161, 195–204. [Google Scholar] [CrossRef]
- Braithwaite, R.A.; McEvoy, L.A. Marine biofouling on fish farms and its remediation. Adv. Mar. Biol. 2005, 47, 215–252. [Google Scholar] [PubMed]
- Baxter, E.J.; Sturt, M.M.; Ruane, N.M.; Doyle, K.; Mcallen, R.; Rodger, H.D. Biofouling of the hydroid Ectopleura larynx on aquaculture nets in Ireland: Implications for finfish health. Fish Vet. J. 2012, 13, 17–29. [Google Scholar]
- Bloecher, N.; Olsen, Y.; Guenther, J. Variability of biofouling communities on fish cage nets: A 1-year field study at a Norwegian salmon farm. Aquaculture 2013, 416–417, 302–309. [Google Scholar] [CrossRef]
- Greene, J.K.; Grizzle, R.E. Successional development of fouling communities on open ocean aquaculture fish cages in the western Gulf of Maine, USA. Aquaculture 2007, 262, 289–301. [Google Scholar] [CrossRef]
- Fitridge, I.; Dempster, T.; Guenther, J.; de Nys, R. The impact and control of biofouling in marine aquaculture: A review. Biofouling 2012, 28, 649–669. [Google Scholar] [CrossRef] [PubMed]
- Fitridge, I.; Keough, M.J. Ruinous resident: The hydroid Ectopleura crocea negatively affects suspended culture of the mussel Mytilus galloprovincialis. Biofouling 2013, 29, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gonzalez, V.; Sanchez-Jerez, P. Fouling assemblages associated with off-coast aquaculture facilities: An overall assessment of the Mediterranean Sea. Mediterr. Mar. Sci. 2017, 18, 87–96. [Google Scholar] [CrossRef]
- Leblanc, A.; Landry, T.; Miron, G. Fouling organisms of the blue mussel Mytilus edulis: Their effect on nutrient uptake and release. J. Shellfish Res. 2003, 22, 633–638. [Google Scholar]
- Martell, L.; Bracale, R.; Carrion, S.A.; Purcell, J.E.; Lezzi, M.; Gravili, C.; Piraino, S.; Boero, F. Successional dynamics of marine fouling hydroids (Cnidaria: Hydrozoa) at a finfish aquaculture facility in the Mediterranean Sea. PLoS ONE 2018, 13, e0195352. [Google Scholar]
- Guenther, J.; Misimi, E.; Sunde, L.M. The development of biofouling, particularly the hydroid Ectopleura larynx, on commercial salmon cage nets in Mid-Norway. Aquaculture 2010, 300, 120–127. [Google Scholar] [CrossRef]
- Bosch-Belmar, M.; Azzurro, E.; Pulis, K.; Milisenda, G.; Fuentes, V.; Kéfi-Daly Yahia, O.; Micallef, A.; Deidun, A.; Piraino, S. Jellyfish blooms perception in Mediterranean finfish aquaculture. Mar. Policy 2017, 76, 1–7. [Google Scholar] [CrossRef]
- Baxter, E.J.; Rodger, H.D.; McAllen, R.; Doyle, T.K. Gill disorders in marine-farmed salmon: Investigating the role of hydrozoan jellyfish. Aquac. Environ. Interact. 2011, 1, 245–257. [Google Scholar] [CrossRef]
- Bosch-Belmar, M.; Milisenda, G.; Girons, A.; Taurisano, V.; Accoroni, S.; Totti, C.; Piraino, S.; Fuentes, V. Consequences of Stinging Plankton Blooms on Finfish Mariculture in the Mediterranean Sea. Front. Mar. Sci. 2017, 4, 240. [Google Scholar] [CrossRef]
- Östman, C.; Piraino, S.; Kem, W. Nematocysts of the Mediterranean hydroid Halocordyle disticha. Hydrobiologia 1991, 216–217, 607–613. [Google Scholar] [CrossRef]
- Östman, C.; Myrdal, M.; Nyvall, P.; Lindström, J.; Segura, A.A. Nematocysts in Tubularia larynx (Cnidaria, Hydrozoa) from Scandinavia and the northern coast of Spain. Sci. Mar. 1995, 59, 165–179. [Google Scholar]
- Tezcan, Ö.D.; Sarp, S. An unusual marine envenomation following a rope contact: A report on nine cases of dermatitis caused by Pennaria disticha. Toxicon 2013, 61, 125–128. [Google Scholar] [CrossRef]
- Di Camillo, C.G.; Giordano, G.; Bo, M.; Betti, F.; Mori, M.; Puce, S.; Bavestrello, G. Seasonal patterns in the abundance of Ectopleura crocea (Cnidaria: Hydrozoa) on a shipwreck in the Northern Adriatic. Mar. Ecol. 2013, 34, 25–32. [Google Scholar] [CrossRef]
- Boero, F.; Brotz, L.; Gibbons, M.; Piraino, S.; Zampardi, S. Ocean Warming 3.10 Impacts and Effects of Ocean Warming on Jellyfish. In Explaining Ocean Warming: Causes, Scale, Effects and Consequences; Laffoley, D., Baxter, J.M., Eds.; IUCN: Gland, Switzerland, 2016; p. 460. [Google Scholar]
- Schuchert, P. The European athecate hydroids and their medusae (Hydrozoa, Cnidaria): Capitata Part 1. Rev. Suisse Zool. 2006, 113, 337–555. [Google Scholar] [CrossRef]
- Allman, G.J. A monograph of the gymnoblastic or tubularian hydroids. In Conclusion of Part I, and Part II, Containing Descriptions of the Genera and Species of Gymnoblastea; Ray Society: London, UK, 1872; pp. 155–450. [Google Scholar]
- Schuchert, P. The European athecate hydroids and their medusae (Hydrozoa, Cnidaria): Capitata Part 2. Rev. Suisse Zool. 2010, 117, 337–555. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bouillon, J.; Medel, M.D.; Pagès, F.; Gili, J.-M.; Boero, F.; Gravili, C. Fauna of the Mediterranean Hydrozoa. Sci. Mar. 2004, 68, 5–438. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information—Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Bavestrello, G.; Puce, S.; Cerrano, C.; Zocchi, E.; Boero, N. The problem of seasonality of benthic hydroids in temperate waters. Chem. Ecol. 2006, 22, S197–S205. [Google Scholar] [CrossRef]
- González-Duarte, M.M.; Megina, C.; Piraino, S.; Cervera, J.L. Hydroid assemblages across the Atlantic-Mediterranean boundary: Is the Strait of Gibraltar a marine ecotone? Mar. Ecol. 2013, 34, 33–40. [Google Scholar] [CrossRef]
- Marcos-López, M.; Mitchell, S.O.; Rodger, H.D. Pathology and mortality associated with the mauve stinger jellyfish Pelagia noctiluca in farmed Atlantic salmon Salmo salar L. J. Fish Dis. 2016, 39, 111–115. [Google Scholar] [CrossRef]
- Doyle, T.K.; De Haas, H.; Cotton, D.; Dorschel, B.; Cummins, V.; Houghton, J.D.R.; Davenport, J.; Hays, G.C. Widespread occurrence of the jellyfish Pelagia noctiluca in Irish coastal and shelf waters. J. Plankton Res. 2008, 30, 963–968. [Google Scholar] [CrossRef]
- Purcell, J.E.; Baxter, E.J.; Fuentes, V.L. Jellyfish as Products and Problems of Aquaculture. In Advances in Aquaculture Hatchery Technology; Allan, G., Burnell, G., Eds.; Woodhead Publishing: Oxford, UK, 2013; pp. 404–430. ISBN 9780857097460. [Google Scholar]
- Båmstedt, U.; Fosså, J.H.; Martinussen, M.B.; Fosshagen, A. Mass occcurrence of the physonect siphophore Apolemia Uvaria (Lesueur) in Norwegian waters. Sarsia 1998, 83, 79–85. [Google Scholar] [CrossRef]
- Mitchell, S.O.; Rodger, H.D. A review of infectious gill disease in marine salmonid fish. J. Fish Dis. 2011, 34, 411–432. [Google Scholar] [CrossRef]
- Rajagopal, S.; Venugopalan, V.P.; Nair, K.V.K.; van der Velde, G.; Jenner, H.A.; den Hartog, C. Reproduction, growth rate and culture potential of the green mussel, Perna viridis (L.) in Edaiyur backwaters, east coast of India. Aquaculture 1998, 162, 187–202. [Google Scholar] [CrossRef]
- Folino-Rorem, N.C.; Renken, C.J. Effects of salinity on the growth and morphology of the invasive, euryhaline hydroid Cordylophora (Phylum Cnidaria, Class Hydrozoa). Invertebr. Biol. 2018, 137, 78–90. [Google Scholar] [CrossRef]
- Piraino, S.; Fanelli, G. Keystone Species: What Are We Talking About? Conserv. Ecol. 1999, 3, 10–12. [Google Scholar] [CrossRef]
- Piraino, S.; Fanelli, G.; Boero, F. Variability of species’ roles in marine communities: Change of paradigms for conservation priorities. Mar. Biol. 2002, 140, 1067–1074. [Google Scholar]
- Folino-Rorem, N.C. Phylum Cnidaria. In Ecology and General Biology: Thorp & Covich’s Freshwater Invertebrates; Thorp, J., Rogers, D.C., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 159–179. [Google Scholar]
- Boero, F.; Bouillon, J.; Piraino, S.; Schmid, V. Asexual Reproduction in the Hydrozoa (Cnidaria). In Reproductive Biology of Invertebrates—Progress in Asexual Reproduction; Hughes, R.N., Ed.; Oxford & IBH Publishing: New Delhi, India, 2002; pp. 141–158. ISBN 0471489689. [Google Scholar]
- Carl, C.; Guenther, J.; Sunde, L.M. Larval release and attachment modes of the hydroid Ectopleura larynx on aquaculture nets in Norway. Aquac. Res. 2011, 42, 1056–1060. [Google Scholar] [CrossRef]
- Boero, F. Fluctuations and variations in coastal marine environments. Mar. Ecol. 1994, 15, 3–25. [Google Scholar] [CrossRef]
- Boero, F.; Belmonte, G.; Fanelli, G.; Piraino, S.; Rubino, F. The continuity of living matter and the discontinuities of its constituents: Do plankton and benthos really exist? Trends Ecol. Evol. 1996, 11, 177–180. [Google Scholar] [CrossRef]
- Boero, F.; Bouillon, J.; Gravili, C.; Piraino, S. Who cares about the Hydrozoa of the Mediterranean Sea? An essay on the zoogeography of inconspicuous groups. Biogeographia 2003, 24, 101–113. [Google Scholar] [CrossRef]
- Boero, F.; Bouillon, J.; Gravili, C.; Miglietta, M.P.; Parsons, T.; Piraino, S. Gelatinous plankton: Irregularities rule the world (sometimes). Mar. Ecol. Prog. Ser. 2008, 356, 299–310. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage of Development | Ectopleura larynx | Pennaria disticha |
|---|---|---|
| 0 | No reproductive structures | No reproductive structures |
| 1 | Small gonophores with no distinct structures fixed as sporosacs above hydranth tentacles | Small gonophores oblong medusoids arising on short pedicels just above whorl of long filiform tentacles |
| 2 | Gonophores oval to spherical. Female gonophores with red spadix that can protrude out of sporosac opening (opening is terminal) | Developed eumedusoid with four radial canals and four marginal bulbs, with small velum, without ocelli, tentacles normally absent |
| 3 | Mature female gonophores with four tentacle-like tubercles around opening at distal end. Female sporosacs filled with numerous small cells forming an egg-like mass | Not applicable |
| 4 | Form of processes very variable, occasionally reduced or absent, but usually increase in size with the enlarging gonophores. Visible developed actinula inside the sporosac | Not applicable |
| Species | Average Period I | Average Period II | Contribution % | Cumulative % |
|---|---|---|---|---|
| Anthozoa | 0.32 | 0 | 21.82 | 21.82 |
| Nemertea | 0.07 | 0 | 18.54 | 40.36 |
| Polychaeta | 0.005 | 0 | 17.31 | 57.68 |
| Mollusca | 0.006 | 0.17 | 16.66 | 74.34 |
| Hydrozoa | 0.81 | 0.18 | 15.08 | 89.42 |
| Period I | Period II | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t1 | t2 | t3 | t4 | t5 | t6 | t7 | t8 | t9 | t10 | t11 | t12 | t13 | t14 | t15 | t16 | |
| Crustacea | ||||||||||||||||
| S.O. Caprellida | X | X | X | X | X | X | X | X | X | X | X | X | X | X | ||
| S.O. Gammaridea | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X |
| S.O. Tanaidomorpha | X | X | X | X | X | X | X | |||||||||
| Polychaeta | ||||||||||||||||
| Fam. Nereidae | X | X | X | X | X | X | X | X | ||||||||
| Fam. Syllidae | X | X | ||||||||||||||
| Nematoda | ||||||||||||||||
| Unidentified | X | X | X | X | X | X | X | X | X | X | X | X | ||||
| Nemertea | ||||||||||||||||
| Unidentified | X | X | X | X | X | X | X | X | ||||||||
| Planaria | ||||||||||||||||
| Unidentified | X | X | X | |||||||||||||
| Mollusca | ||||||||||||||||
| Mytilus galloprovincialis | X | X | X | X | X | X | X | X | ||||||||
| Musculus sp. | X | X | X | X | ||||||||||||
| Arca sp. | X | |||||||||||||||
| Irus sp. | X | |||||||||||||||
| Doto sp. | X | X | X | X | ||||||||||||
| Fam. Hiatellidae | X | |||||||||||||||
| Chlamys sp. | X | |||||||||||||||
| Briozoa | ||||||||||||||||
| Or. Cyclostomatida | X | X | ||||||||||||||
| Anthozoa | ||||||||||||||||
| Or. Actiniaria | X | X | X | X | X | X | X | X | X | |||||||
| Hydrozoa | ||||||||||||||||
| Pennaria disticha | X | X | X | X | X | X | ||||||||||
| Halecium pusillum | X | X | X | X | ||||||||||||
| Obelia dichotoma | X | X | X | X | X | X | X | X | X | X | X | |||||
| Ectopleura larynx | X | X | X | X | X | X | X | X | ||||||||
| Coryne eximia | X | X | X | X | ||||||||||||
| Sertularella ellisii | X | |||||||||||||||
| Eudendrium racemosum | X | |||||||||||||||
| Echinodermata | ||||||||||||||||
| Arbacea sp. | X | |||||||||||||||
| Algae | ||||||||||||||||
| Ceramium sp. | X | X | X | X | X | X | ||||||||||
| Antithamnionella sp. | X | X | X | X | X | X | X | |||||||||
| Polysiphonia sp. | X | X | X | X | X | X | X | X | X | X | X | |||||
| Giraudia sp. | X | |||||||||||||||
| Hincksia sp. | X | X | X | X | X | X | ||||||||||
| Jania sp. | X | X | X | X | X | |||||||||||
| Laurencia sp. | X | |||||||||||||||
| Trichleocarpa sp. | X | X | X | X | ||||||||||||
| Bryopsis sp. (2) | X | X | X | X | X | X | ||||||||||
| Chaetomorpha sp. | X | X | X | X | ||||||||||||
| Cladophora sp. | X | X | ||||||||||||||
| Period | Model | AICc | p | R2 |
|---|---|---|---|---|
| I | M1 | 420 | 2.2 × 10−16 | 0.86 |
| I | M2 | 469 | 2.2 × 10−16 | 0.72 |
| II | M1 | 473 | 2.2 × 10−16 | 0.92 |
| II | M2 | 561 | 2.2 × 10−16 | 0.81 |
| Period I | Df | Sum Sq | Mean Sq | F Value | Pr (>F) |
|---|---|---|---|---|---|
| Species | 1 | 3099.9 | 3099.9 | 167.787 | <2.2−16 |
| Time | 1 | 4001.7 | 4001.7 | 216.600 | <2.2−16 |
| Species × time | 1 | 1290.2 | 1290.2 | 69.834 | 4.932−12 |
| Residuals | 68 | 1256.3 | 18.5 | ||
| Period II | |||||
| Species | 1 | 1894.25 | 1894.25 | 427.25 | <2.2−16 |
| Time | 1 | 2706.73 | 2706.73 | 610.51 | <2.2−16 |
| Species × time | 1 | 600.98 | 600.98 | 135.55 | <2.2−16 |
| Residuals | 104 | 461.09 | 4.43 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bosch-Belmar, M.; Escurriola, A.; Milisenda, G.; Fuentes, V.L.; Piraino, S. Harmful Fouling Communities on Fish Farms in the SW Mediterranean Sea: Composition, Growth and Reproductive Periods. J. Mar. Sci. Eng. 2019, 7, 288. https://doi.org/10.3390/jmse7090288
Bosch-Belmar M, Escurriola A, Milisenda G, Fuentes VL, Piraino S. Harmful Fouling Communities on Fish Farms in the SW Mediterranean Sea: Composition, Growth and Reproductive Periods. Journal of Marine Science and Engineering. 2019; 7(9):288. https://doi.org/10.3390/jmse7090288
Chicago/Turabian StyleBosch-Belmar, Mar, Agnés Escurriola, Giacomo Milisenda, Verónica L. Fuentes, and Stefano Piraino. 2019. "Harmful Fouling Communities on Fish Farms in the SW Mediterranean Sea: Composition, Growth and Reproductive Periods" Journal of Marine Science and Engineering 7, no. 9: 288. https://doi.org/10.3390/jmse7090288
APA StyleBosch-Belmar, M., Escurriola, A., Milisenda, G., Fuentes, V. L., & Piraino, S. (2019). Harmful Fouling Communities on Fish Farms in the SW Mediterranean Sea: Composition, Growth and Reproductive Periods. Journal of Marine Science and Engineering, 7(9), 288. https://doi.org/10.3390/jmse7090288