Rhodolith Beds Heterogeneity along the Apulian Continental Shelf (Mediterranean Sea)

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
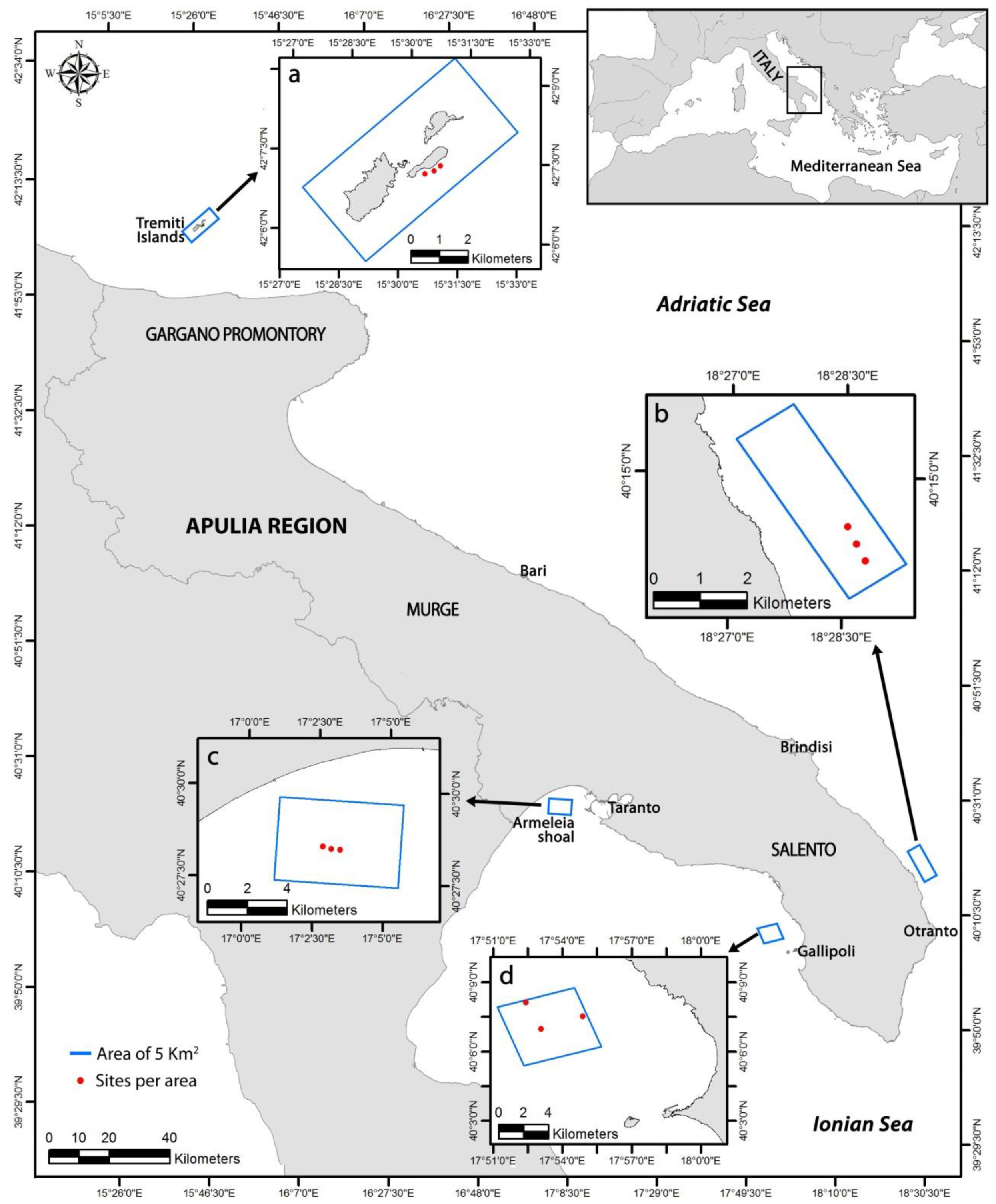
2.1. Study Areas
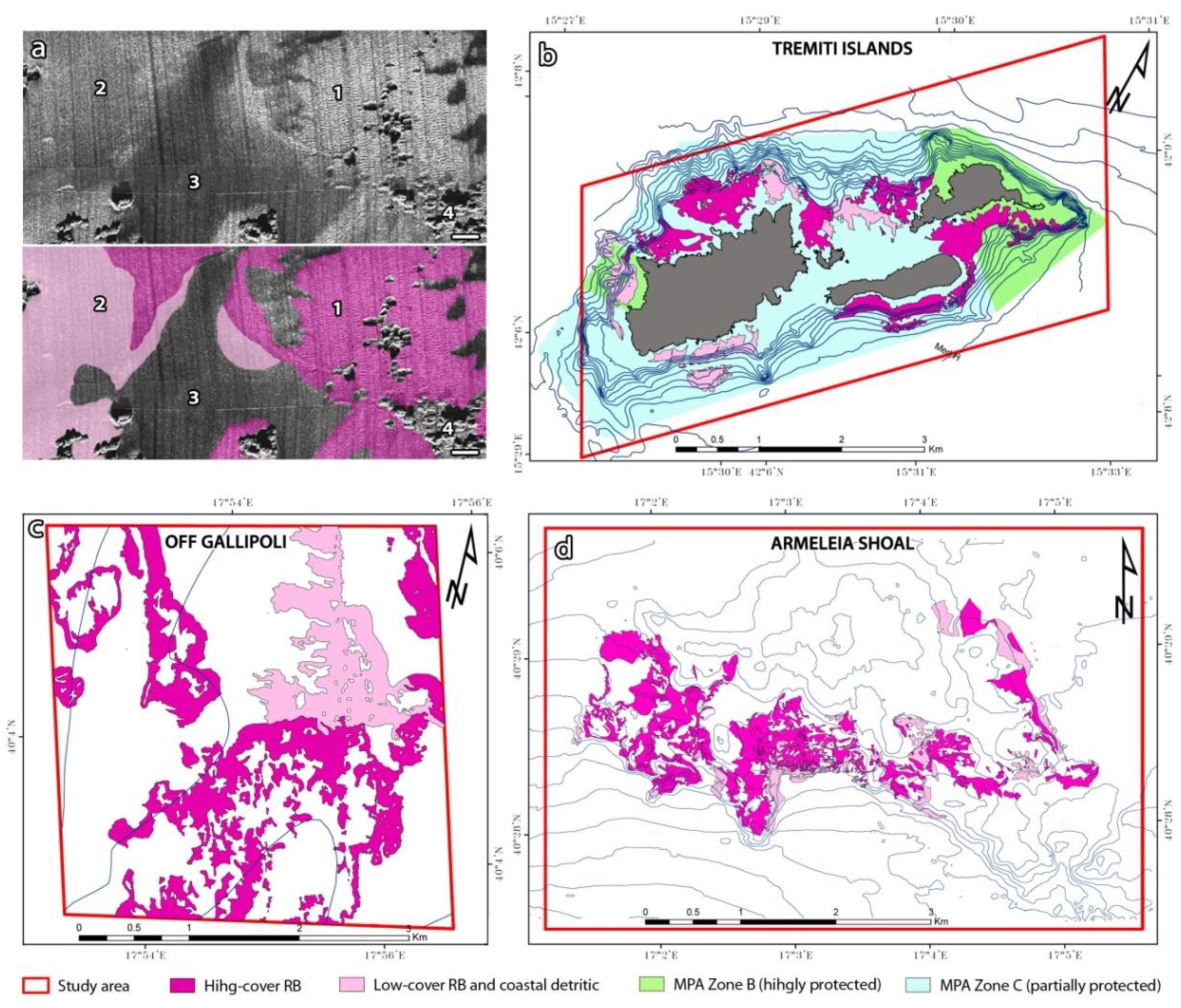
2.2. Habitat Mapping
2.3. Data Collection
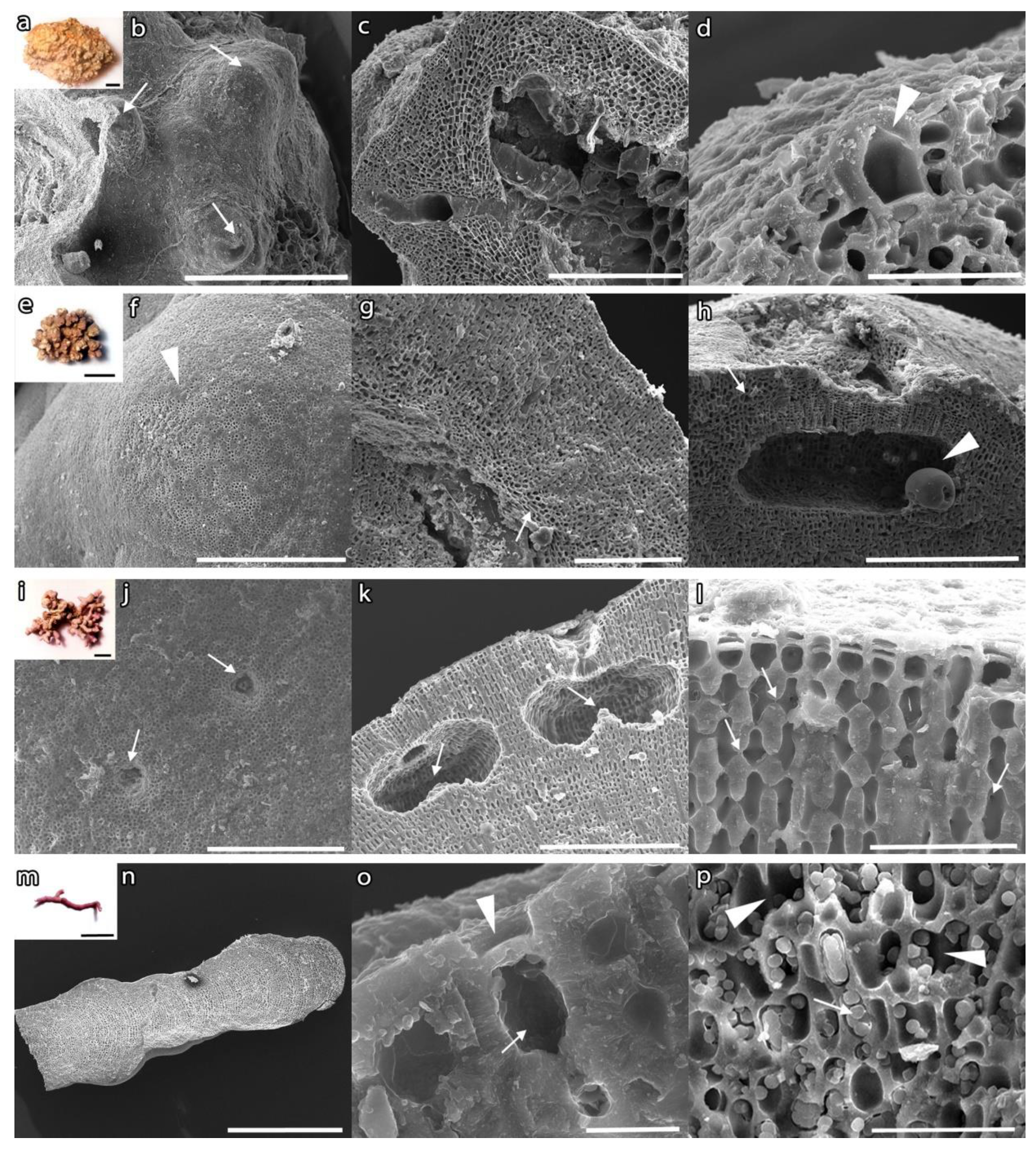
2.4. Taxonomic Identification
2.5. Statistical Analysis
3. Results
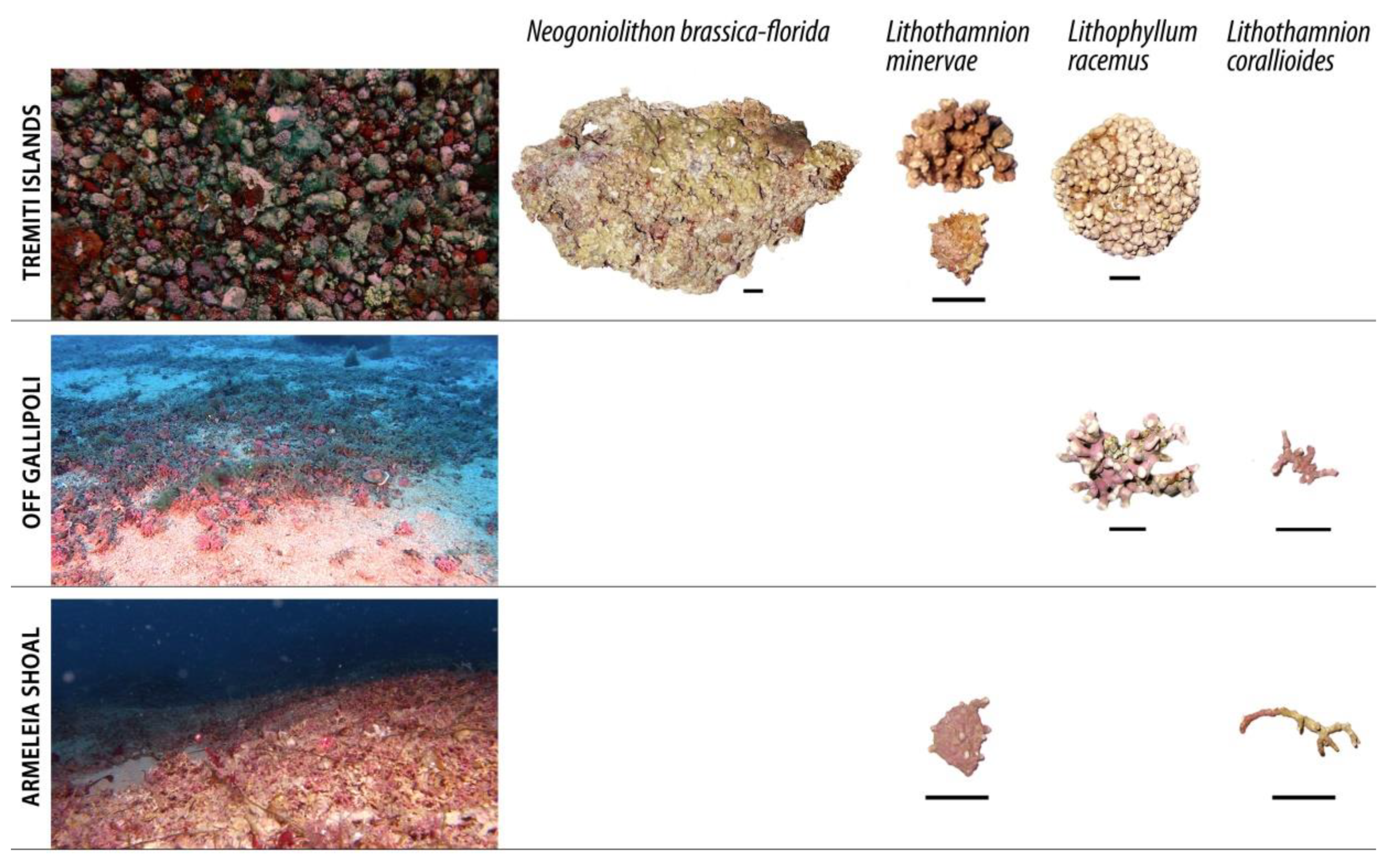
3.1. Mapping and Description of Rhodolith Beds
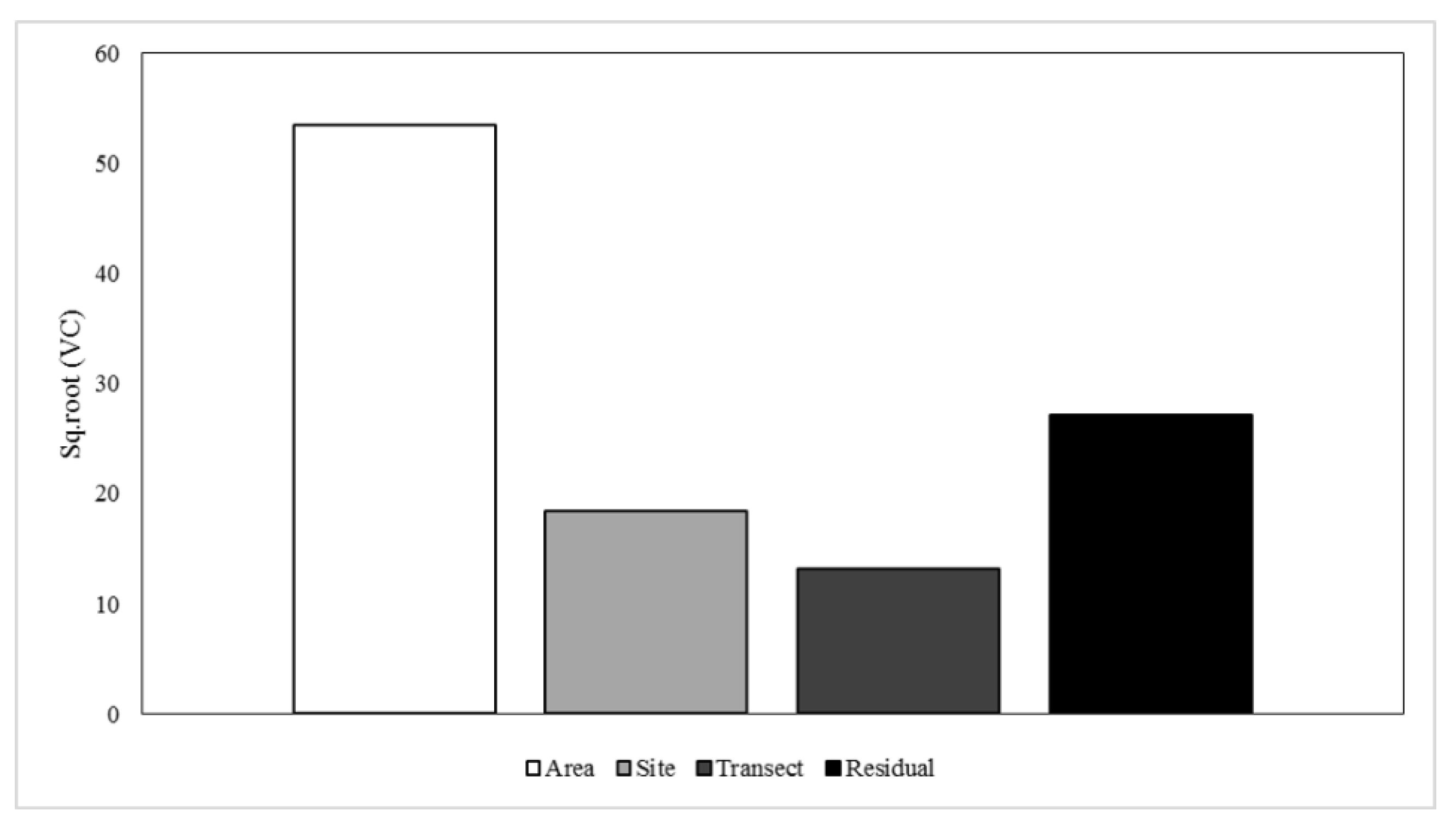
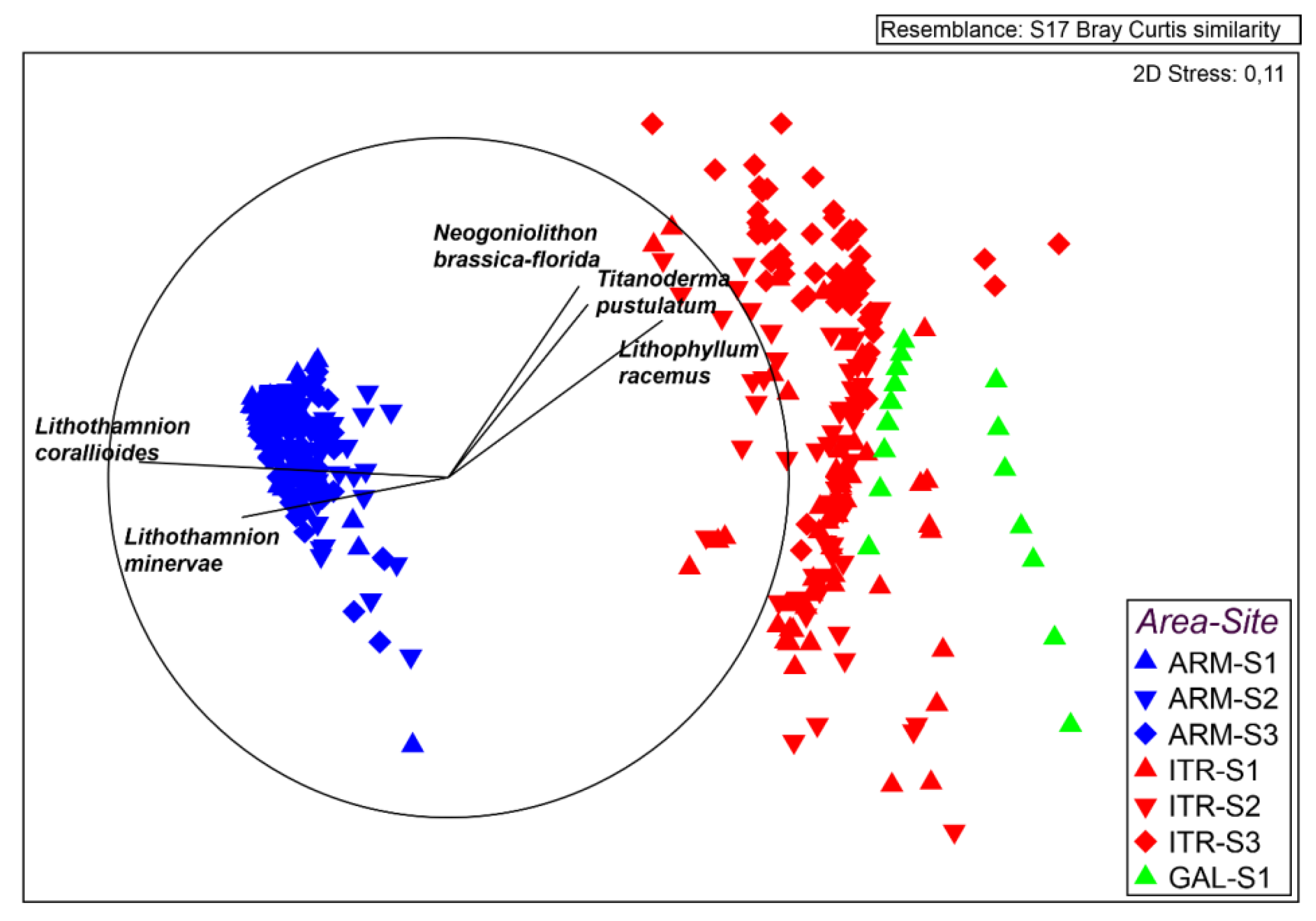
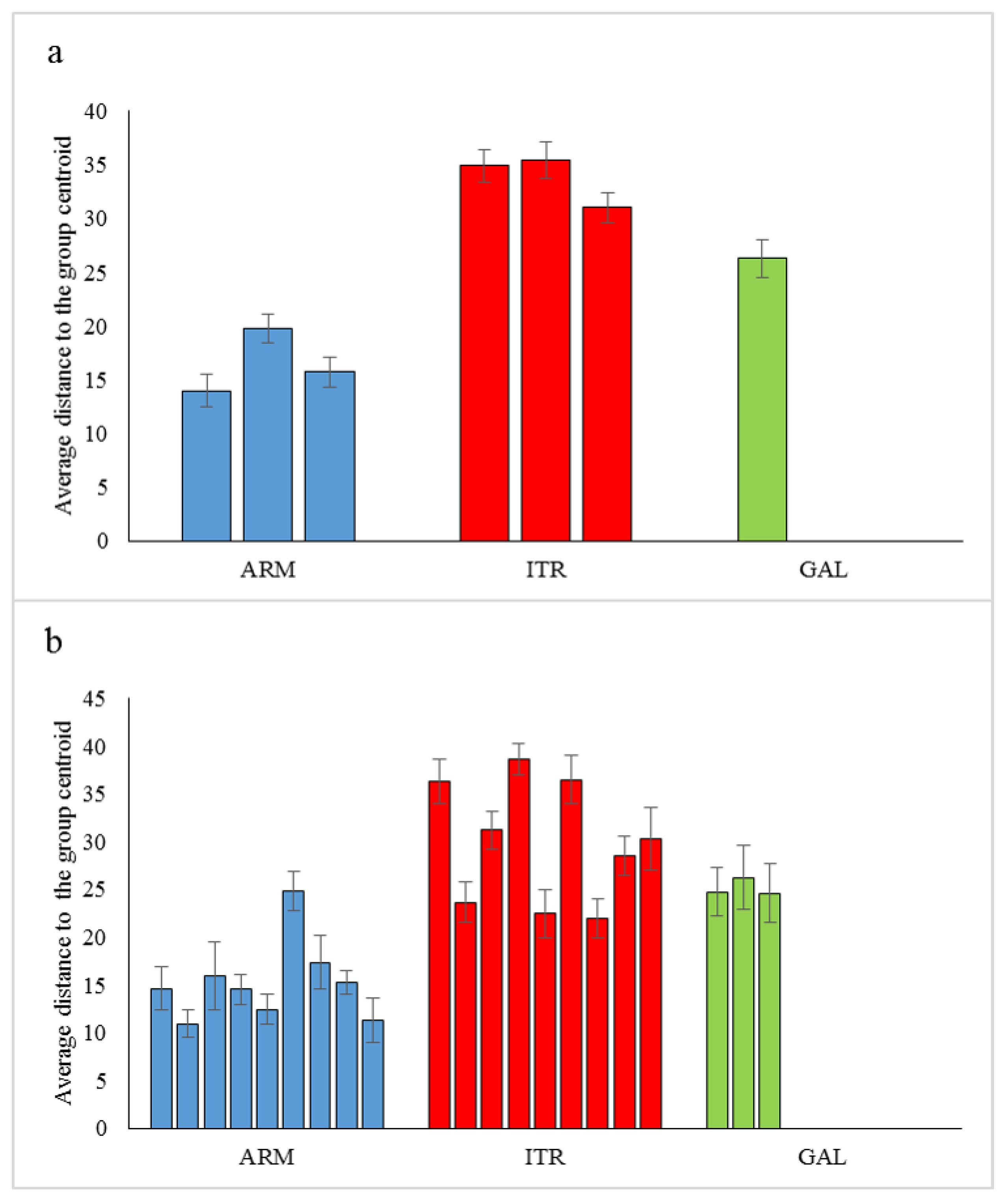
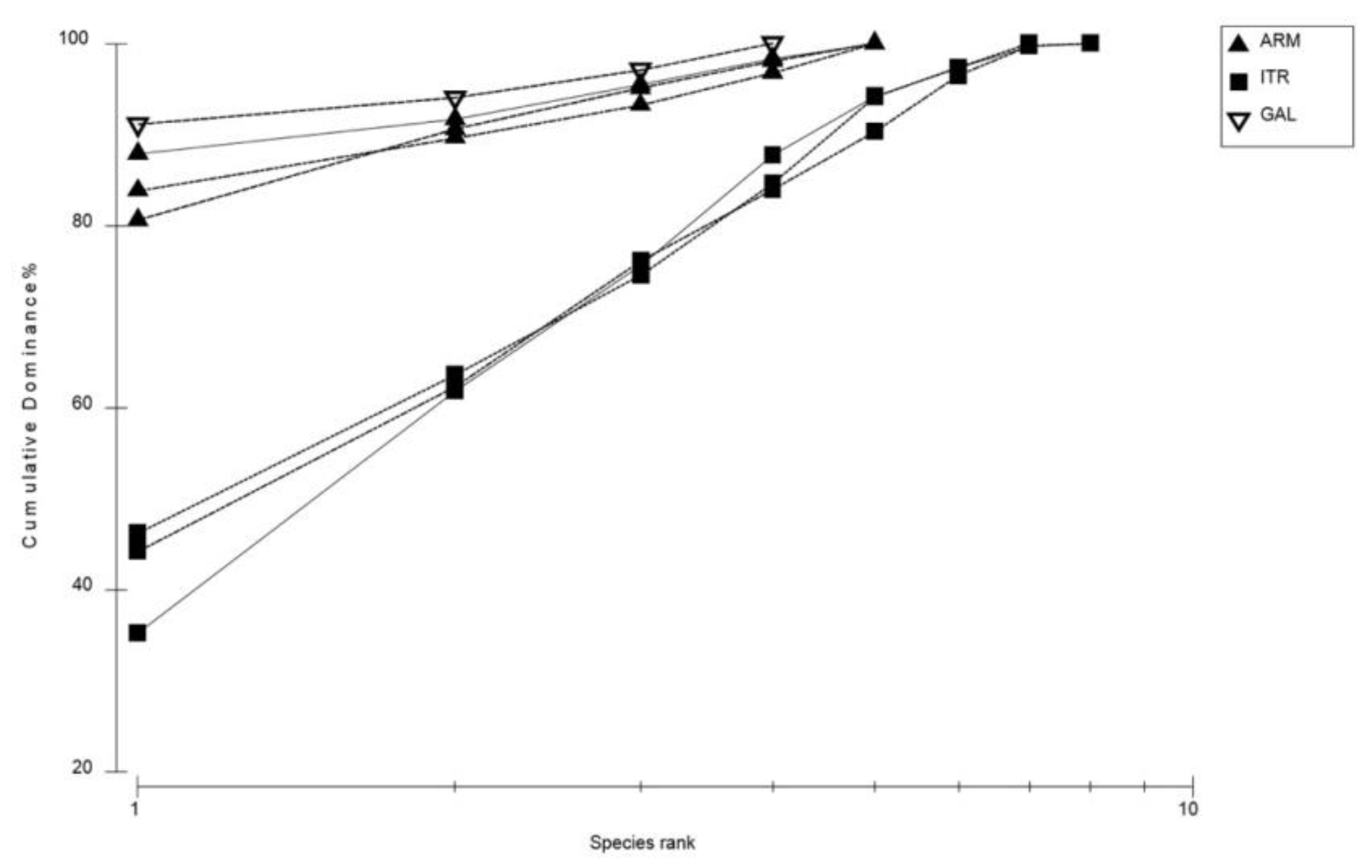
3.2. Heterogeneity of Rhodolith Beds
4. Discussion
4.1. Rhodolith Beds Variability from Small to Large Scale
4.2. Rhodolith Morphotypes and Main Hydrodynamic Regime
4.3. Threats and Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Crain, M.C.; Kroeker, K.; Halpern, B.S. Interactive and cumulative effects of multiple human stressors in marine systems. Ecol. Lett. 2008, 11, 1304–1315. [Google Scholar]
- Vinebrooke, R.D.; Cottingham, K.L.; Norberg, J.; Scheffer, M.; Dodson, S.I.; Maberly, S.C.; Sommer, U. Impacts of multiple stressors on biodiversity and ecosystem functioning: The role of species co-tolerance. Oikos 2004, 104, 451–457. [Google Scholar]
- Chimienti, G.; Bo, M.; Mastrototaro, F. Know the distribution to assess the changes: Mediterranean cold-water coral bioconstructions. Rend. Lincei Sci. Fis. Nat. 2018, 29, 583–588. [Google Scholar]
- Zajac, R.M.; Vozarik, J.M.; Gibbons, B.R. Spatial and temporal patterns in macrofaunal diversity components relative to sea floor landscape structure. PLoS ONE 2013, 8, e65823. [Google Scholar]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar]
- Steller, D.L.; Riosmena-Rodriguez, R.; Foster, M.S.; Roberts, C.A. Rhodolith bed diversity in the Gulf of California: The importance of rhodolith structure and consequences of disturbance. Aquat. Conserv. 2003, 13, S5–S20. [Google Scholar]
- Ingrosso, G.; Cecchi, L.; Bertolino, M.; Bevilacqua, S.; Bianchi, C.N.; Bo, M.; Boscari, E.; Cardone, F.; Cattaneo-Vietti, R.; Cau, A.; et al. Mediterranean Bioconstructions along the Italian Coast. Adv. Mar. Biol. 2018, 79, 61–136. [Google Scholar]
- Sañé, E.; Chiocci, F.L.; Basso, D.; Martorelli, E. Environmental factors controlling the distribution of rhodoliths: An integrated study based on seafloor sampling, ROV and side scan sonar data, offshore the W-Pontine Archipelago. Cont. Shelf Res. 2016, 129, 10–22. [Google Scholar]
- Grall, J.; Glemarec, M. Biodiversite des fonds de maërl en Bretagne: Approache fonctionnelle et impacts anthropogeniques. Vie Milieu 1997, 47, 339–349. [Google Scholar]
- Foster, M.S.; Filho, G.M.A.; Kamenos, K.A.; Riosmena-Rodríguez, R.; Steller, D.L. Rhodoliths and rhodolith beds. In Research and Discoveries: The Revolution of Science Through SCUBA; American Academy of Underwater Sciences: Mobile, AL, USA, 2013; pp. 143–155. [Google Scholar]
- Ordines, F.; Bauzá, M.; Sbert, M.; Roca, P.; Gianotti, M.; Massutí, E. Red algal beds increase the condition of nekto-benthic fish. J. Sea Res. 2015, 95, 115–123. [Google Scholar]
- Riosmena-Rodríguez, R.; Nelson, W.; Aguirre, J. (Eds.) Rhodolith/Maërl Beds: A Global Perspective; Coastal Research Library 15; Springer: Cham, Switzerland, 2017; pp. 1–362. [Google Scholar]
- Jacquotte, R. Etude des fonds de maerl de Méditerranée. Recueil Travaux StationMarine d’Endoume 1962, 26, 141–216. [Google Scholar]
- Hall-Spencer, J.M.; Grall, J.; Moore, P.G.; Atkinson, R.J.A. Bivalve fishing and maerl-bed conservation in France and the UK-retrospect and prospect. Aquat. Conserv. 2003, 13, S33–S41. [Google Scholar]
- Barberá, C.; Moranta, J.; Ordines, F.; Ramón, M.; de Mesa, A.; Díaz-Valdés, M.; Grau, A.M.; Massutí, E. Biodiversity and habitat mapping of Menorca Channel (western Mediterranean): Implications for conservation. Biodivers. Conserv. 2012, 21, 701–728. [Google Scholar]
- Cavalcanti, G.S.; Gregoraci, G.B.; Santos, E.O.; Silveira, C.B.; Meirelles, P.M.; Longo, L.; Gotoh, K.; Nakamura, S.; Iida, T.; Sawabe, T.; et al. Physiologic and metagenomic attributes of the rhodoliths forming the largest CaCO3 bed in the South Atlantic Ocean. ISME J. 2014, 8, 52–62. [Google Scholar]
- Basso, D.; Babbini, L.; Kaleb, S.; Bracchi, V.; Falace, A. Monitoring deep Mediterranean rhodolith beds. Aquat. Conserv. 2016, 26, 549–561. [Google Scholar]
- Krayesky-Self, S.; Schmidt, W.; Phung, D.; Henry, C.; Sauvage, T.; Camacho, O.; Felgenhauer, B.E.; Fredericq, S. Eukaryotic life inhabits rhodolith-forming coralline algae (Hapalidiales, Rhodophyta), remarkable marine benthic microhabitats. Sci. Rep. 2017, 7, 45850. [Google Scholar]
- Coletti, G.; Basso, D.; Frixa, A. Economic importance of coralline carbonates. In Rhodolith/Maërl Beds: A Global Perspective; Riosmena-Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Coastal Research Library—Springer: Cham, Switzerland, 2017; Volume 15, pp. 87–101. [Google Scholar]
- Schubert, N.; Schoenrock, K.M.; Aguirre, J.; Kamenos, N.A.; Silva, J.; Horta, P.A.; Hofmann, L.C. Coralline algae: Globally distributed ecosystem engineers. Front. Mar. Sci. 2020, 7, 352. [Google Scholar]
- Peña, V.; Barbara, I. Maerl community in the north-west Iberian Peninsula: A review of floristic studies and long-term changes. Aquat. Conserv. 2008, 18, 339–366. [Google Scholar]
- Sciberras, M.; Rizzo, M.; Mifsud, J.R.; Camilleri, K.; Borg, J.A.; Lanfranco, E.; Schembri, P.J. Habitat structure and biological characteristics of a maerl bed off the northeastern coast of the Maltese Islands (central Mediterranean). Mar. Biodivers. 2009, 39, 251–264. [Google Scholar]
- Di Geronimo, R.; Giaccone, G. Le alghe calcaree nel detritico costiero di Lampedusa (Isole Pelagie). Boll. Accad. Gioenia Sci. Nat. 1994, 27, 5–25. [Google Scholar]
- Falace, A.; Kaleb, S.; Agnesi, S.; Annunziatellis, A.; Salvati, E.; Tunesi, L. Macroalgal composition of rhodolith beds in a pilot area of the Tuscan Archipelago (Tyrrhenian Sea): Primary elements to evaluate the degree of conservation of this habitat. In Proceedings of the 2nd Mediterranean Symposium on the conservation of Coralligenous & other Calcareous Bio-Concretions, Portorož, Slovenia, 29–30 October 2014; pp. 213–214. [Google Scholar]
- Basso, D.; Babbini, L.; Ramos-Esplá, A.A.; Salomidi, M. Mediterranean Rhodolith Beds. In Rhodolith/Maërl Beds: A Global Perspective; Riosmena-Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Coastal Research Library—Springer: Cham, Switzerland, 2017; Volume 15, pp. 281–298. [Google Scholar]
- Rendina, F.; Kaleb, S.; Caragnano, A.; Ferrigno, F.; Appolloni, L.; Donnarumma, L.; Russo, G.F.; Sandulli, R.; Roviello, V.; Falace, A. Distribution and characterization of deep rhodolith beds off the Campania coast (SW Italy, Mediterranean Sea). Plants 2020, 9, 985. [Google Scholar] [CrossRef]
- European Parliament. Council of the European Union Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 establishing a framework for Community action in the field of marine environmental policy (Marine Strategy Framework Directive). Off. J. Eur. Union 2008, L164, 19. [Google Scholar]
- European Commission. Commission decision of 1 September 2010 on criteria and methodological standards on good environmental status of marine waters. Off. J. Eur. Union 2010, L232, 14. [Google Scholar]
- European Parliament. Council directive 1992/43/EC Conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, L206, 7–50. [Google Scholar]
- European Community. Council Decision 82/72/EEC of 3 December 1981 concerning the conclusion of the Convention on the conservation of European wildlife and natural habitats (Bern Convention). Off. J. Eur. Union 1982, L38, 1–2. [Google Scholar]
- European Parliament. Council Regulation No. 1967/2006 concerning management measures for the sustainable exploitation of fishery resources in the Mediterranean Sea, amending Regulation (EEC) No 2847/93 and repealing Regulation (EC) No 1626/94. Off. J. Eur. Union 2006, L409, 11. [Google Scholar]
- United Nations Environmental Program. Proposal of Standard Methods for Inventorying and Monitoring Coralligenous and Rhodoliths Populations; UNEP(DEPI)/MED WG.359/10; RAC/SPA: Tunis, Tunisia, 2011; pp. 1–23. [Google Scholar]
- Steller, D.L.; Foster, M.S. Environmental factors influencing distribution and morphology of rhodoliths in Bahía Concepción, B.C.S., México. J. Exp. Mar. Biol. Ecol. 1995, 194, 201–212. [Google Scholar]
- Steller, D.L.; Hernández-Ayón, J.M.; Riosmena-Rodríguez, R.; Cabello-Pasini, A. Effect of temperature on photosynthesis, growth and calcification rates of the free-living coralline alga Lithophyllum margaritae. Ciencias Mar. 2007, 33, 441–456. [Google Scholar]
- Nelson, W.; Neill, K.; Farr, T.; Barr, N.; D’Archino, R.; Miller, S.; Stewart, R. Rhodolith Beds in Northern New Zealand: Characterisation of Associated Biodiversity and Vulnerability to Environmental Stressors; New Zealand Aquatic Environment and Biodiversity Report; Ministry for Primary Industries: Wellington, New Zeland, 2012; pp. 1–106. [Google Scholar]
- Barberá, C.; Bordehore, C.; Borg, J.A.; Glemarec, M.; Grall, J.; Hall-Spencer, J.M.; De la Huz, C.; Lanfranco, E.; Lastra, M.; Moore, P.G.; et al. Conservation and management of northeast Atlantic and Mediterranean maërl beds. Aquat. Conserv. 2003, 13, S65–S76. [Google Scholar]
- Peña, V.; Barbara, I. Seasonal patterns in the mäerl community of shallow European Atlantic beds and their use as a baseline for monitoring studies. Eur. J. Phycol. 2010, 45, 327–342. [Google Scholar]
- Pardo, C.; Lopez, L.; Peña, V.; Hernández-Kantún, J.; Le Gall, L.; Bárbara, I.; Barreiro, R. A multilocus species delimitation reveals a striking number of species of coralline algae forming maërl in the OSPAR Maritime Area. PLoS ONE 2014, 9, e104073. [Google Scholar]
- Thrush, S.F.; Schneider, D.C.; Legendre, P.; Whitlatch, R.B.; Dayton, P.K.; Hewitt, J.E.; Hines, A.H.; Cummings, V.J.; Lawrie, S.M.; Grantet, J.; et al. Scaling-up from experiments to complex ecological systems: Where to next? J. Exp. Mar. Biol. Ecol. 1997, 216, 243–254. [Google Scholar]
- Wootton, J.T. Local interactions predict large-scale pattern in empirically derived cellular automata. Nature 2001, 413, 841–843. [Google Scholar]
- Irving, A.D.; Connell, S.D.; Gillanders, B.M. Local complexity in patterns of canopy-benthos associations produces regional patterns across temperate Australasia. Mar. Biol. 2004, 144, 361–368. [Google Scholar]
- Fraschetti, S.; Bianchi, C.N.; Terlizzi, A.; Fanelli, G.; Morri, C.; Boero, F. Spatial variability and human disturbance in shallow subtidal hard substrate assemblages: A regional approach. Mar. Ecol. Prog. Ser. 2001, 212, 1–12. [Google Scholar]
- Fraschetti, S.; Terlizzi, A.; Benedetti-Cecchi, L. Patterns of distribution of marine assemblages from rocky shores: Evidence of relevant scales of variation. Mar. Ecol. Prog. Ser. 2005, 296, 13–29. [Google Scholar]
- Steneck, R.S. The ecology of coralline algal crusts: Convergent patterns and adaptive strategies. Ann. Rev. Ecol. Syst. 1986, 17, 273–303. [Google Scholar]
- Lanfranco, E.; Rizzo, M.; Hall-Spencer, J.; Borg, J.A.; Schembri, P.J. Maerl-forming coralline algae and associated phytobenthos from the Maltese Islands. Cent. Mediterr. Nat. 1999, 3, 1–6. [Google Scholar]
- Ballesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. Oceanogr. Mar. Biol. 2006, 44, 123–195. [Google Scholar]
- Chimienti, G.; Stithou, M.; Dalle Mura, I.; Mastrototaro, F.; D’Onghia, G.; Tursi, A.; Izzi, C.; Fraschetti, S. An explorative assessment of the importance of mediterranean coralligenous habitat to local economy: The case of recreational diving. J. Environ. Account. Manag. 2017, 5, 315–325. [Google Scholar]
- Piazzi, L.; Kaleb, S.; Ceccherelli, G.; Montefalcone, M.; Falace, A. Deep coralligenous outcrops of the Apulian continental shelf: Biodiversity and spatial variability of sediment-regulated assemblages. Cont. Shelf. Res. 2019, 172, 50–56. [Google Scholar]
- Pérès, J.M.; Picard, J. Nouveau manuel de bionomie benthique de la Mer Méditerranée. Rec. Travl St. Mar. Endoume 1964, 31, 5–137. [Google Scholar]
- Bedulli, D.; Bianchi, C.N.; Zurlini, G.; Morri, C. Caratterizzazione biocenotica e strutturale del macrobenthos delle coste pugliesi. In Indagine Ambientale del Sistema Marino Costiero della Regione Puglia; Viel, M., Zurlini, G., Eds.; ENEA: Rome, Italy, 1986; pp. 227–255. [Google Scholar]
- Damiani, V.; Bianchi, C.N.; Ferretti, O.; Bedulli, D.; Morri, C.; Viel, M.; Zurlini, G. Risultati di una ricerca ecologica sul sistema marino costiero pugliese. Thalassia Salentina 1988, 18, 153–169. [Google Scholar]
- Matarrese, A.; Panza, M.; Mastrototaro, F.; Costantino, G. Preliminare rappresentazione cartografica dei fondali dell’arcipelago delle Isole Tremiti (Mar Adriatico). Biol. Mar. Mediterr. 2000, 7, 590–593. [Google Scholar]
- Tropeano, M.; Spalluto, L. Present-day temperate-type carbonate sedimentation on Apulia shelves (southern Italy). GeoActa 2006, 5, 129–142. [Google Scholar]
- Martin, C.; Giannoulaki, M.; De Leo, F.; Scardi, M.; Salomidi, M.; Knittweis, L.; Pace, M.L.; Garofalo, G.; Gristina, M.; Ballesteros, E. Coralligenous and maërl habitats: Predictive modelling to identify their spatial distributions across the Mediterranean Sea. Sci. Rep. 2015, 4, 5073. [Google Scholar]
- Cushman-Roisin, B.; Gacic, M.; Poulain, P.M.; Artegiani, A. Physical Oceanography of the Adriatic Sea: Past, Present and Future; Springer: Dordrecht, The Netherlands, 2001; pp. 1–304. [Google Scholar]
- Millot, C.; Taupier-Letage, I. Circulation in the Mediterranean Sea. In The Mediterranean Sea. Handbook of Environmental Chemistry; Sailot, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 5K, pp. 29–66. [Google Scholar]
- Chimienti, G.; De Padova, D.; Mossa, M.; Mastrototaro, F. A mesophotic black coral forest in the Adriatic Sea. Sci. Rep. 2020, 10, 1–15. [Google Scholar]
- Aiello, G.; Budillon, F. Lowstand prograding wedges as fourth-order glacio-eustatic cycles in the Pleistocene continental shelf of Apulia (southern Italy). In Cyclostratigraphy: Approaches and Case Histories; D’Argenio, B., Fisher, A.G., Premoli Silva, I., Weissert, H., Ferreri, V., Eds.; Society for Sedimentary Geology: Tusla, OK, USA, 2014; pp. 215–230. [Google Scholar]
- Senatore, M.R. Terrazzi deposizionali sommersi lungo il margine ionico della Puglia. Mem. Descr. Carta Geol. d’It. 2004, 58, 141–146. [Google Scholar]
- Poulain, P.M. Adriatic Sea surface circulation as derived from drifter data between 1990 and 1999. J. Mar. Syst. 2001, 29, 3–32. [Google Scholar]
- Artale, V.; Zoccolotti, L. Alcuni aspetti della circolazione dell’Adriatico e formazione di acque dense. In Indagine Ambientale del Sistema Marino Costiero della Regione Puglia; Viel, M., Zurlini, G., Eds.; ENEA: Rome, Italy, 1986; pp. 87–99. [Google Scholar]
- Mosetti, F. Caratteristiche fondamentali dell’idrologia dell’Adriatico. Boll. Ocean. Teor. Appl. 1984, 2, 169–194. [Google Scholar]
- Artegiani, A.; Paschini, E.; Russo, A.; Bregnt, D.; Raicich, F.; Pinardi, N. The Adriatic Sea general circulation. Part II: Baroclinic circulation structure. J. Phys. Oceanogr. 1997, 27, 1515–1532. [Google Scholar]
- Cattaneo, A.; Correggiari, A.; Langone, L.; Trincardi, F. The late-Holocene Gargano subaqueous delta, Adriatic shelf: Sediment pathways and supply fluctuations. Mar. Geol. 2003, 193, 61–91. [Google Scholar]
- Caldara, M.; Centenaro, E.; Mastronuzzi, G.; Sansò, P.; Sergio, A. Features and present evolution of Apulian coast (Southern Italy). J. Coast. Res. 1998, SI26, 55–64. [Google Scholar]
- Gasparini, G.P.; Griffa, A.L. Studio delle condizioni dinamiche nel Golfo di Taranto. In Indagine Ambientale del Sistema Marino Costiero della Regione Puglia; Viel, M., Zurlini, G., Eds.; ENEA: Rome, Italy, 1986; pp. 101–125. [Google Scholar]
- Savini, A.; Basso, D.; Bracchi, V.A.; Corselli, C.; Pennetta, M. Maerl-bed mapping and carbonate quantification on submerged terraces offshore the Cilento peninsula (Tyrrhenian Sea, Italy). Geodiversitas 2012, 34, 77–98. [Google Scholar]
- Chimienti, G.; Angeletti, L.; Rizzo, L.; Tursi, A.; Mastrototaro, F. ROV vs trawling approaches in the study of benthic communities: The case of Pennatula rubra (Cnidaria: Pennatulacea). J. Mar. Biol. Assoc. 2018, 98, 1859–1869. [Google Scholar]
- Weinberg, S. The minimal area problem in invertebrate communities of Mediterranean rocky substrata. Mar. Biol. 1978, 49, 33–40. [Google Scholar]
- Chimienti, G. Vulnerable forests of the pink sea fan Eunicella verrucosa in the Mediterranean Sea. Diversity 2020, 12, 176. [Google Scholar] [CrossRef]
- Kaleb, S.; Alongi, G.; Falace, A. Coralline algae preparation for scanning electron microscopy and optical microscopy. In Protocols for Macroalgae Research; Charrier, B., Wichard, T., Reddy, C.R.K., Eds.; Taylor and Franciss Group: Boca Raton, FL, USA, 2018; pp. 413–429. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2020. Available online: http://www.algaebase.org (accessed on 20 July 2020).
- Clarke, K.R.; Gorley, R.H. PRIMER V6: User Manual/Tutorial; Primer-e: Plymouth, UK, 2001. [Google Scholar]
- Anderson, M. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- Anderson, M.J.; ter Braak, C.J.F. Permutation tests for multi-factorial analysis of variance and regression. J. Stat. Comput. Simul. 2003, 73, 85–113. [Google Scholar]
- Anderson, M.J.; Diebel, C.E.; Blom, W.M.; Landers, T.J. Consistency and variation in kelp holdfast assemblages: Spatial patterns of biodiversity for the major phyla at different taxonomic resolutions. J. Exp. Mar. Biol. Ecol. 2005, 320, 35–56. [Google Scholar]
- Anderson, M.J. Distance-based tests for homogeneity of multivariate dispersion. Biometrics 2006, 62, 245–253. [Google Scholar]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar]
- Lambshead, P.J.D.; Piatt, H.M.; Shaw, K.M. The detection of differences among assemblages of marine benthic species based on an assessment of dominance and diversity. J. Nat. Hist. 1983, 17, 859–974. [Google Scholar]
- Rocha, G.A.; Bastos, A.C.; Amado-Filho, G.M.; Boni, G.C.; Moura, R.L.; Oliveira, N. Heterogeneity of rhodolith beds expressed in backscatter data. Mar. Geol. 2020, 423, 106136. [Google Scholar]
- Le Bas, T.P.; Huvenne, V.A.I. Acquisition and processing of backscatter data for habitat mapping—Comparison of multibeam and sidescan systems. Appl. Acoust. 2009, 70, 1248–1257. [Google Scholar]
- Otero-Ferrer, F.; Cosme, M.; Tuya, F.; Espino, F.; Haroun, R. Effect of depth and seasonality on the functioning of rhodolith seabeds. Estuar. Coast. Shelf. Sci. 2020, 235, 106579. [Google Scholar]
- Thrush, S.F.; Pridmore, R.D.; Hewitt, J.E. Impacts on soft-sediment macrofauna: The effects of spatial variation on temporal trends. Ecol. Appl. 1994, 4, 31–41. [Google Scholar]
- Coleman, R.A.; Browne, M.; Theobalds, T. Small-scale spatial variability in intertidal and subtidal turfing algal assemblages and the temporal generality of these patterns. J. Exp. Mar. Biol. Ecol. 2002, 267, 53–74. [Google Scholar]
- Schubert, N.; Salazar, V.W.; Rich, W.A.; Bercovich, M.V.; Saá, A.A.; Fadigas, S.D.; Silva, J.; Horta, P.A. Rhodolith primary and carbonate production in a changing ocean: The interplay of warming and nutrients. Sci. Total Environ. 2019, 676, 455–468. [Google Scholar]
- Carvalho, V.F.; Assis, J.; Serrão, E.A.; Nunes, J.M.; Anderson, A.B.; Batista, M.B.; Barufi, J.B.; Silva, J.; Pereira, S.M.B.; Horta, P.A. Environmental drivers of rhodolith beds and epiphytes community along the South Western Atlantic coast. Mar. Environ. Res. 2020, 154, 104827. [Google Scholar]
- Agnesi, S.; Annunziatellis, A.; Inghilesi, R.; Orasi, A. The contribution of wind-wave energy at sea bottom to the modelling of rhodolith beds distribution in an off-shore continental shelf. Mediterr. Mar. Sci. 2020, 21, 433–441. [Google Scholar]
- Bosellini, A.; Ginsburg, R. Form and internal structure of recent algal nodules (Rhodolites) from Bermuda. J. Geol. 1971, 79, 669–682. [Google Scholar]
- Crocetta, F.; Riginella, E.; Lezzi, M.; Tanduo, V.; Balestrieri, L.; Rizzo, L. Bottom-trawl catch composition in a highly polluted coastal area reveals multifaceted native biodiversity and complex communities of fouling organisms on litter discharge. Mar. Environ. Res. 2020, 155, 104875. [Google Scholar]
- Rendina, F.; Ferrigno, F.; Appolloni, L.; Donnarumma, L.; Sandulli, R.; Fulvio, G. Anthropic pressure due to lost fishing gears and marine litter on different rhodolith beds off the Campania Coast (Tyrrhenian Sea, Italy). Ecol. Quest. 2020, 31, 1–17. [Google Scholar]
- Barberá, C.; Mallol, S.; Vergés, A.; Cabanellas-Reboredo, M.; Díaz, D.; Goñi, R. Maerl beds inside and outside a 25-year-old no-take area. Mar. Ecol. Prog. Ser. 2017, 572, 77–90. [Google Scholar]
- Gennaro, P.; Piazzi, L. Synergism between two anthropic impacts: Caulerpa racemosa var. cylindracea invasion and seawater nutrient enrichment. Mar. Ecol. Prog. Ser. 2011, 427, 59–70. [Google Scholar]
- Klein, J.; Verlaque, M. The Caulerpa racemosa invasion: A critical review. Mar. Pollut. Bull. 2008, 56, 205–225. [Google Scholar]
- Piazzi, L.; Balata, D.; Bulleri, F.; Gennaro, P.; Ceccherelli, G. The invasion of Caulerpa cylindracea in the Mediterranean: The known, the unknown and the knowable. Mar. Biol. 2016, 163, 161. [Google Scholar]
- Rizzo, L.; Pusceddu, A.; Bianchelli, S.; Fraschetti, S. Potentially combined effect of the invasive seaweed Caulerpa cylindracea (Sonder) and sediment deposition rates on organic matter and meiofaunal assemblages. Mar. Environ. Res. 2020, 159, 104966. [Google Scholar]
- Rizzo, L.; Pusceddu, A.; Stabili, L.; Alifano, P.; Fraschetti, S. Potential effects of an invasive seaweed (Caulerpa cylindracea, Sonder) on sedimentary organic matter and microbial metabolic activities. Sci. Rep. 2017, 7, 12113. [Google Scholar]
- Martin, S.; Gattuso, J.P. Response of Mediterranean coralline algae to ocean acidification and elevated temperature. Glob. Chang. Biol. 2009, 15, 2089–2100. [Google Scholar]
- McCoy, S.J.; Ragazzola, F. Skeletal trade-offs in coralline algae in response to ocean acidification. Nat. Clim. Chang. 2014, 4, 719–723. [Google Scholar]
- Rindi, F.; Braga, J.; Martin, S.; Peña, V.; Le Gall, L.; Caragnano, A.; Aguirre, J. Coralline algae in a changing Mediterranean Sea: How can we predict their future, if we do not know their present? Front. Mar. Sci. 2019, 6, 723. [Google Scholar]
- Martin, S.; Cohu, S.; Vignot, C.; Zimmerman, G.; Gattuso, J.P. One-year experiment on the physiological response of the Mediterranean crustose coralline alga, Lithophyllum cabiochae, to elevated pCO2 and temperature. Ecol. Evol. 2013, 3, 676–693. [Google Scholar]
- Vásquez-Elizondo, R.M.; Enríquez, S. Coralline algal physiology is more adversely affected by elevated temperature than reduced pH. Sci. Rep. 2016, 6, 19030. [Google Scholar]
- Martin, S.; Hall-Spencer, J.M. Effects of ocean warming and acidification on rhodolith/maërl beds. In Rhodolith/Maërl Beds: A Global Perspective; Riosmena-Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 55–85. [Google Scholar]
- Chimienti, G.; Bo, M.; Taviani, M.; Mastrototaro, F. Occurrence and Biogeography of Mediterranean Cold-Water Corals. In Mediterranean Cold-Water Corals: Past, Present and Future; Orejas, C., Jimennez, C., Eds.; Springer International Publishing: Berlin, Germany, 2019; Volume 9, pp. 213–243. [Google Scholar]
- Morato, T.; González-Irusta, J.M.; Dominguez-Carrió, C.; Wei, C.L.; Davies, A.; Sweetman, A.K.; Taranto, G.H.; Beazley, L.; García-Alegre, A.; Grehan, A.; et al. Climate-induced changes in the suitable habitat of cold-water corals and commercially important deep-sea fishes in the North Atlantic. Glob. Chang. Biol. 2020, 26, 2181–2202. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Northwest Otranto | Tremiti Islands | Off Gallipoli | Armeleia Shoal |
|---|---|---|---|---|
| Lithophyllum racemus (Lamarck) Foslie | <2 | 18.59 ± 16.09 | 22.25 ± 13.31 | |
| Lithothamnion corallioides (P. Crouan & H. Crouan) P. Crouan & H. Crouan | 0.73 ± 0.45 | 59.19 ± 20.68 | ||
| Lithothamnion crispatum Hauck | 2.55 ± 2.71 | 0.73 ± 0.45 | 1.78 ± 1.72 | |
| Lithothamnion minervae Basso | 0.22 ± 0.47 | 2.64 ± 2.43 | ||
| Lithothamnion valens Foslie | 0.11 ± 0.33 | |||
| Mesophyllum sp. | 1.30 ± 1.16 | 1.75 ± 1.34 | ||
| Neogoniolithon brassica-florida (Harvey) Setchell & Mason | 12.36 ± 14.06 | |||
| Spongites fruticulosus Kützing | 4.91 ± 5.54 | |||
| Titanoderma pustulatum (J.V. Lamourou) Nägeli | 3.23 ± 3.59 | 0.74 ± 0.44 | 0.11 ± 0.48 | |
| Cover % | 43.27 ± 30.53 | 24.44 ± 14.09 | 65.59 ± 19.57 | |
| Live/Dead rhodolith ratio | 78.25 ± 11.45 | 73.18 ± 9.54 | 50.95 ± 24.89 |
| Proportion of Living Thalli | Rhodolith Cover | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Source | df | MS | Pseudo-F | p (perm) | Variation Components | MS | Pseudo-F | p (perm) | Variation Components |
| Ar | 2 | 35,194.0 | 2.955 | 0.0943 | 185.52 | 43,479.0 | 1.919 | 0.1619 | 165.92 |
| Si(Ar) | 4 | 12,397.0 | 11.162 | 0.0002 * | 188.14 | 23,582.0 | 11.426 | 0.0002 * | 358.70 |
| Tr(Si(Ar)) | 14 | 1097.0 | 5.811 | 0.0001 * | 45.99 | 2038.5 | 6.644 | 0.0001 * | 87.68 |
| Res | 394 | 188.8 | 188.77 | 306.8 | 306.82 | ||||
| Total | 414 | ||||||||
| Source | df | MS | Pseudo-F | p (perm) | Variation Components |
|---|---|---|---|---|---|
| Ar | 2 | 382,000 | 16.25 | * | 2858.60 |
| Si(Ar) | 4 | 24,471.00 | 5.80 | *** | 337.68 |
| Tr(Si(Ar)) | 14 | 4166.70 | 5.64 | *** | 173.55 |
| Res | 394 | 739.04 | 739.04 | ||
| Total | 414 |
| ARM | ITR | GAL | ARM vs. ITR 94.70% | ARM vs. GAL 96.86% | ITR vs. GAL 64.33% | ||||
|---|---|---|---|---|---|---|---|---|---|
| Taxa | Av. | Av. | Av. | Av.Diss | Contrib% | Av.Diss | Contrib% | Av.Diss | Contrib% |
| L. corallioides | 59.19 | 0.00 | 0.73 | 51.78 | 54.68 | 61.42 | 63.41 | 1.17 | 1.82 |
| L. racemus | 0.00 | 18.59 | 22.25 | 14.26 | 15.05 | 23.33 | 24.08 | 26.09 | 40.55 |
| C. cylindracea | 4.58 | 8.12 | 0.0 | 7.50 | 7.92 | 4.70 | 4.85 | 7.50 | 11.66 |
| N. brassica-florida | 0.00 | 12.37 | 0.00 | 9.24 | 9.76 | 0.00 | 0.00 | 15.11 | 23.49 |
| S. fruticulosus | 0.00 | 4.91 | 0.00 | 3.61 | 3.81 | 0.00 | 0.00 | 5.77 | 8.97 |
| L. minervae | 2.64 | 0.00 | 0.00 | 2.59 | 2.74 | 3.18 | 3.28 | 0.00 | 0.00 |
| T. pustulatum | 0.00 | 3.23 | 0.73 | 2.50 | 2.64 | 0.78 | 0.81 | 3.17 | 4.93 |
| L. crispatum | 1.78 | 2.55 | 0.73 | 1.93 | 2.04 | 1.34 | 1.39 | 3.41 | 5.29 |
| Mesophyllum sp. | 1.75 | 1.30 | 0.00 | 1.10 | 1.16 | 2.12 | 2.19 | 1.86 | 2.88 |
| C. prolifera | 0.00 | 0.40 | 0.00 | 0.20 | 0.21 | 0.00 | 0.00 | 0.26 | 0.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chimienti, G.; Rizzo, L.; Kaleb, S.; Falace, A.; Fraschetti, S.; Giosa, F.D.; Tursi, A.; Barbone, E.; Ungaro, N.; Mastrototaro, F. Rhodolith Beds Heterogeneity along the Apulian Continental Shelf (Mediterranean Sea). J. Mar. Sci. Eng. 2020, 8, 813. https://doi.org/10.3390/jmse8100813
Chimienti G, Rizzo L, Kaleb S, Falace A, Fraschetti S, Giosa FD, Tursi A, Barbone E, Ungaro N, Mastrototaro F. Rhodolith Beds Heterogeneity along the Apulian Continental Shelf (Mediterranean Sea). Journal of Marine Science and Engineering. 2020; 8(10):813. https://doi.org/10.3390/jmse8100813
Chicago/Turabian StyleChimienti, Giovanni, Lucia Rizzo, Sara Kaleb, Annalisa Falace, Simonetta Fraschetti, Francesco De Giosa, Angelo Tursi, Enrico Barbone, Nicola Ungaro, and Francesco Mastrototaro. 2020. "Rhodolith Beds Heterogeneity along the Apulian Continental Shelf (Mediterranean Sea)" Journal of Marine Science and Engineering 8, no. 10: 813. https://doi.org/10.3390/jmse8100813
APA StyleChimienti, G., Rizzo, L., Kaleb, S., Falace, A., Fraschetti, S., Giosa, F. D., Tursi, A., Barbone, E., Ungaro, N., & Mastrototaro, F. (2020). Rhodolith Beds Heterogeneity along the Apulian Continental Shelf (Mediterranean Sea). Journal of Marine Science and Engineering, 8(10), 813. https://doi.org/10.3390/jmse8100813