Shifts in Diatom Dominance Associated with Seasonal Changes in an Estuarine-Mangrove Phytoplankton Community

Abstract
:1. Introduction
2. Materials and Methods
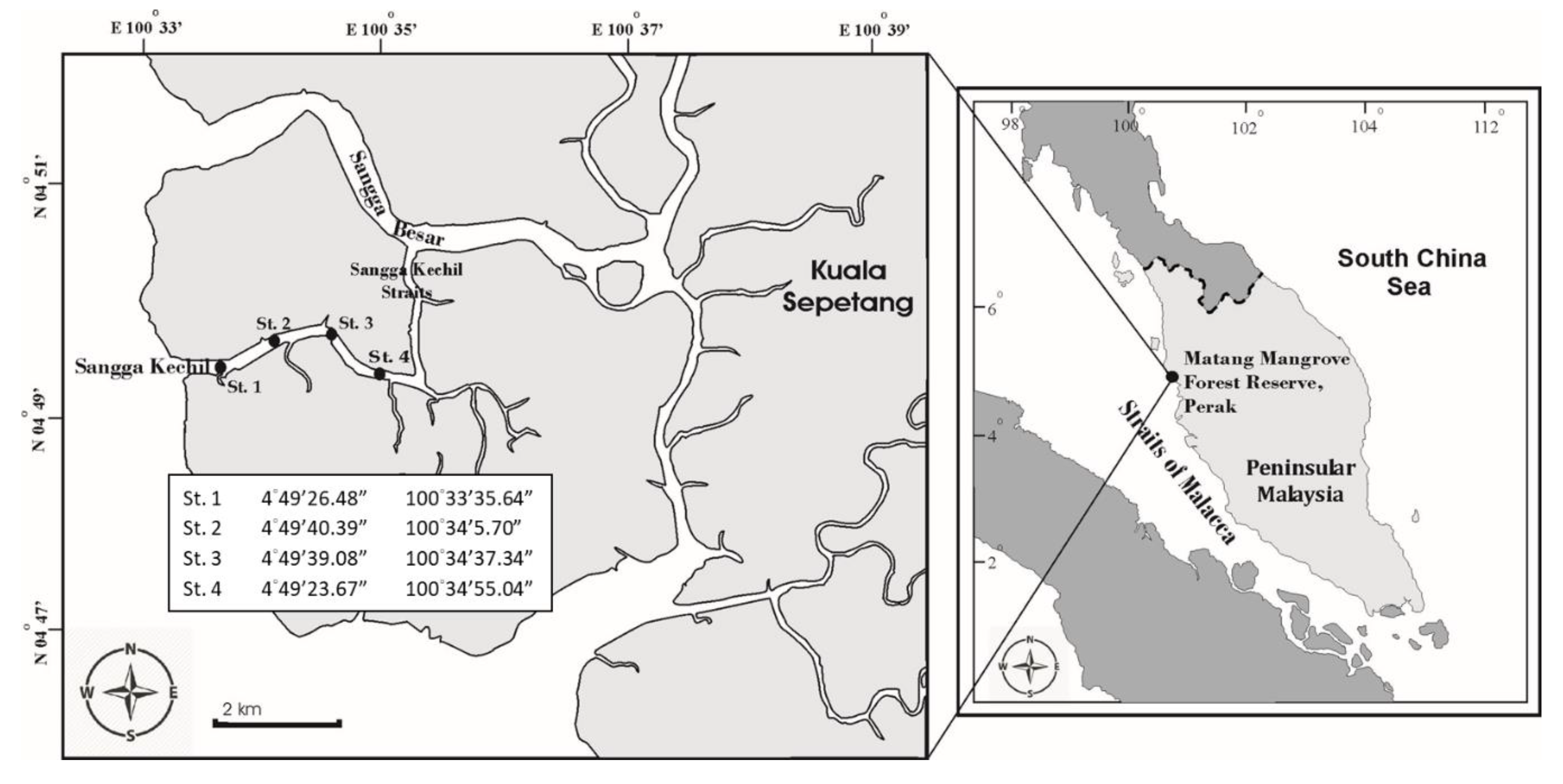
2.1. Study Areas
2.2. Samples and Environmental Data Collection
2.3. Chlorophyll a, Nutrients and Rainfall Data
2.4. Phytoplankton Collection and Enumeration
2.5. Data Analyses
3. Results
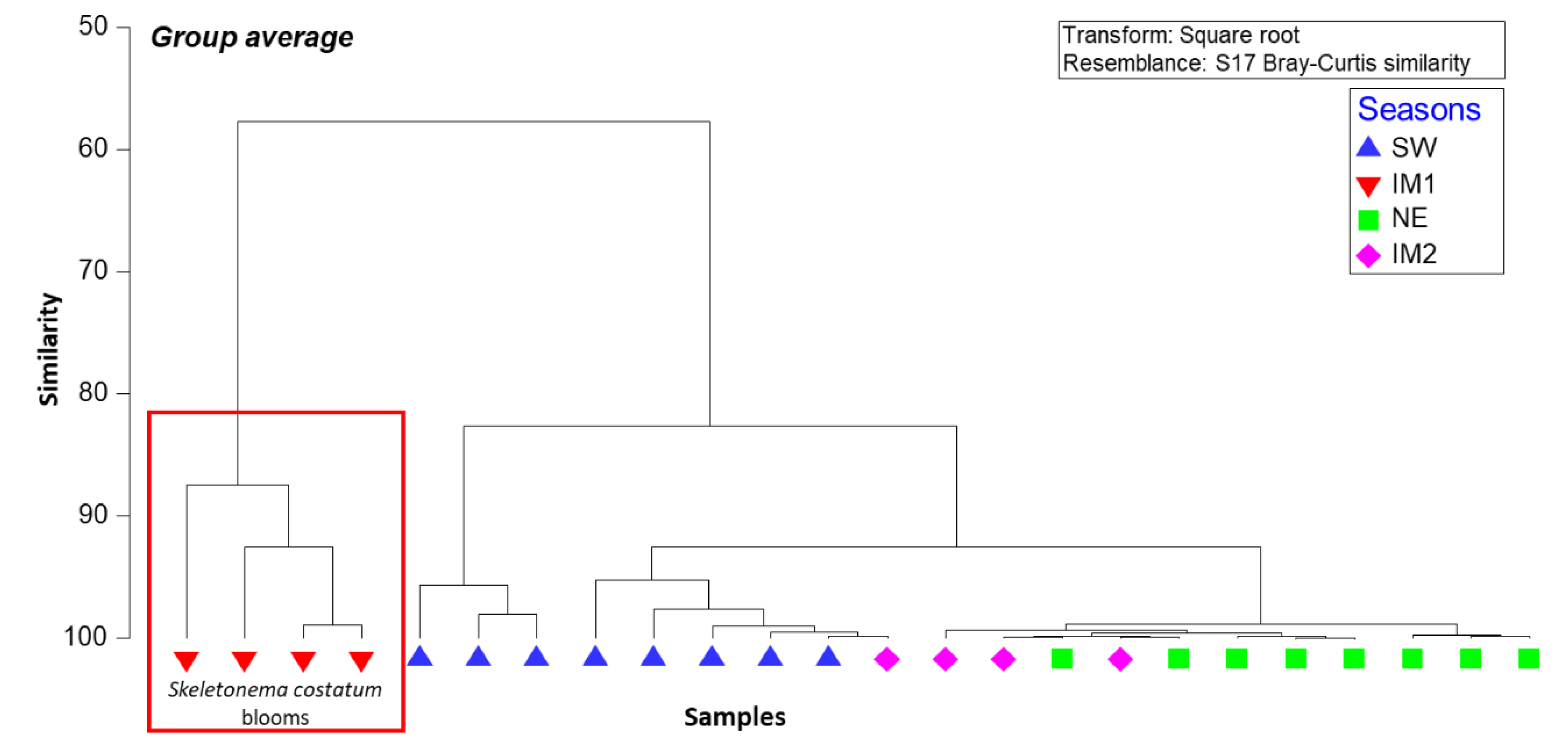
3.1. Diatom Dominated Phytoplankton Population
3.2. Correlation of Physical and Chemical Parameters and CCA Ordination of Dominant Diatoms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Srikanth, S.; Lum, S.K.Y.; Chen, Z. Mangrove root: Adaptations and ecological importance. Tree 2016, 30, 451–465. [Google Scholar] [CrossRef]
- Rovai, A.S.; Twilley, R.R.; Castañeda-Moya, E.; Riul, P.; Cifuentes-Jara, M.; Manrow-Villalobos, M.; Horta, P.A.; Simonassi, J.C.; Fonseca, A.L.; Pagliosa, P.R. Global controls on carbon storage in mangrove soils. Nat. Clim. Chang. 2018, 8, 534–538. [Google Scholar] [CrossRef]
- Woodroffe, C.D.; Rogers, K.; McKee, K.L.; Lovelock, C.E.; Mendelssohn, I.A.; Saintilan, N. Mangrove sedimentation and response to relative sea-level rise. Annu. Rev. Mar. Sci. 2016, 8, 243–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.C.; Shih, S.S.; Ho, Y.S.; Chen, C.P.; Hsieh, H.L. Restoration of shorebird-roosting mudflats by partial removal of estuarine mangroves in northern Taiwan. Restor. Ecol. 2012, 20, 76–84. [Google Scholar] [CrossRef]
- Mitra, A. Mangroves: A natural ecosystem of cultural and religious convergence. In Mangrove Forests in India; Mitra, A., Ed.; Springer: Cham, Switzerland, 2020; pp. 337–352. [Google Scholar]
- Nicholls, P.; Schwarz, A.-M.; Hancock, N. Nutrient enrichment in mangrove ecosystems: A growing concern. Water Atmos. 2004, 12, 16–17. [Google Scholar]
- Lacerda, L.D.; Menezes, M.O.; Molisani, M.M. Changes in mangrove extension at the Pacoti River estuary, CE, NE Brazil due to regional environmental changes between 1958 and 2004. Biota Neotrop. 2007, 7, 67–72. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Ball, M.C.; Martin, K.C.; Feller, I.C. Nutrient enrichment increases mortality of mangroves. PLoS ONE 2009, 4, e5600. [Google Scholar] [CrossRef] [Green Version]
- Postacchini, M.; Romano, A. Dynamics of the coastal zone. J. Mar. Sci. Eng. 2019, 7, 451. [Google Scholar] [CrossRef]
- Rahaman, S.M.; Biswas, S.K.; Rahaman, M.S.; Ghosh, A.K.; Sarder, L.; Siraj, S.M.S.; Islam, S.S. Seasonal nutrient distribution in the Rupsha-Passur tidal river system of the Sundarbans mangrove forest, Bangladesh. Ecol. Process. 2014, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Rassignol, K.L.; Hall, S.N.; Peierls, B.L.; Wetz, M.S. Phytoplankton community indicators of short- and longterm ecological change in the anthropogenically and climatically impacted Neuse River Estuary, North Carolina, USA. Estuar. Coasts. 2010, 33, 485–497. [Google Scholar] [CrossRef]
- Roubeix, V.; Becquevort, S.; Lancelot, C. Influence of bacteria and salinity on diatom biogenic silica dissolution in estuarine systems. Biogeochemistry 2008, 88, 47–62. [Google Scholar] [CrossRef]
- Nixon, S.W.; Olsen, S.B.; Buckley, E.; Fulweiler, R. “Lost to the Tide”—The Importance of Fresh Water Flow to Estuaries; Final Report Submitted to the Coastal Resources Center; Graduate School of Oceanography, University of Rhode Island: Narragansett, RI, USA, 2004; p. 15. [Google Scholar]
- Faridah-Hanum, I.; Yusoff, F.M.; Fitrianto, A.; Ainuddin, N.A.; Gandaseca, S.; Zaiton, S.; Norizah, K.; Nurhidayu, S.; Roslan, M.K.; Hakeem, K.R.; et al. Development of a comprehensive mangrove quality index (MQI) in Matang Mangrove; Assessing mangrove ecosystem health. Ecol. Indic. 2019, 102, 103–117. [Google Scholar] [CrossRef]
- Griffiths, J.R.; Hajdu, S.; Downing, A.S.; Hjerne, O.; Larsson, U.; Winder, M. Phytoplankton community interactions and environmental sensitivity in coastal and offshore habitats. Oikos 2016, 125, 1134–1143. [Google Scholar] [CrossRef]
- Smith, V.H. Responses of estuarine and coastal marine phytoplankton to nitrogen and phosphorus enrichment. Limnol. Oceanogr. 2006, 51, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Rath, A.R.; Mitbavkar, S.; Anil, A.C. Phytoplankton community structure in relation to environmental factors from the New Mangalore Port waters along the southwest coast of India. Environ. Monit. Assess. 2018, 190, 481. [Google Scholar] [CrossRef] [PubMed]
- Wan Maznah, W.O. Perspectives on the use of algae as biological indicators for monitoring and protecting aquatic environments, with special reference to Malaysian freshwater ecosystems. Trop. Life Sci. Res. 2010, 21, 51–67. [Google Scholar]
- Cloern, J.E.; Foster, S.Q.; Kleckner, A.E. Phytoplankton primary production in the world’s estuarine-coastal ecosystems. Biogeosciences 2014, 11, 2477–2501. [Google Scholar] [CrossRef] [Green Version]
- San Martin, E.; Irigoien, X.; Harris, R.P.; López-Urrutia, A.; Zubkov, M.V.; Heywood, J.L. Variation in the transfer energy in marine plankton along a productivity gradient in the Atlantic Ocean. Limnol. Oceanogr. 2006, 51, 2084–2091. [Google Scholar] [CrossRef]
- Armengol, L.; Calbet, A.; Franchy, G.; Rodríguez-Santos, A.; Hernández-León, S. Planktonic food web structure and trophic transfer efficiency along a productivity gradient in the tropical and subtropical Atlantic Ocean. Sci. Rep. 2019, 9, 2044. [Google Scholar] [CrossRef] [Green Version]
- Berdalet, E.; Kudela, R.M.; Banas, N.S.; Bresnan, E.; Burford, M.A.; Davidson, K.; Gobler, C.J.; Karlson, B.; Lim, P.T.; Mackenzie, L.; et al. GlobalHAB: Fostering International Coordination on Harmful Algal Bloom Research in Aquatic Systems. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P., Berdalet, E., Burford, M., Pitcher, G., Zhou, M., Eds.; Ecological Studies (Analysis and Synthesis); Springer: Cham, Switzerland, 2018; pp. 425–447. [Google Scholar]
- Munawar, M.; Fitzpatrick, M.A.J. Eutrophication in three Canadian Areas of Concern: Phytoplankton and major nutrient interactions. Aquat. Ecosyst. Heath 2018, 21, 421–437. [Google Scholar] [CrossRef]
- Boyer, J.N.; Kelble, C.R.; Ortner, P.B.; Rudnick, D.T. Phytoplankton bloom status: Chlorophyll a biomass as an indicator of water quality condition in the southern estuaries of Florida, USA. Ecol. Indic. 2009, 9, S56–S67. [Google Scholar] [CrossRef]
- Rajkumar, M.; Rajasekar, K.T.; Sun, J.; Jenkinson, I.R.; Prabu, V.A.; Perumal, P. Phytoplankton diversity in the Parangipettai coastal waters, southeast coast of India. J. Environ. Biol. 2012, 33, 1115–1125. [Google Scholar]
- Shekhar, T.S.; Kiran, B.R.; Puttaiah, E.T.; Shivaraj, Y.; Mahadevan, K.M. Phytoplankton as index of water quality with reference to industrial pollution. J. Environ. Biol. 2008, 29, 233–236. [Google Scholar] [PubMed]
- Casé, M.; Leça, E.E.; Leitão, S.N.; Sant, E.E.; Schwamborn, R.; de Moraes Junior, A.T. Plankton community as an indicator of water quality in tropical shrimp culture ponds. Mar. Pollut. Bull. 2008, 56, 1343–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, V.R.; Brownlee, C.; Milazzo, M.; Hall-Spencer, J.M. Marine microphytobenthic assemblage shift along a natural shallow-water CO2 gradient subjected to multiple environment stressors. J. Mar. Sci. Eng. 2015, 3, 1425–1447. [Google Scholar] [CrossRef] [Green Version]
- Hilaluddin, F.; Yusoff, F.M.; Natrah, F.M.I.; Lim, P.T. Disturbance of mangrove forests causes alterations in estuarine phytoplankton community structure in Malaysian Matang mangrove forests. Mar. Environ. Res. 2020, 158, 104935. [Google Scholar] [CrossRef]
- Hilaluddin, F.; Leaw, C.P.; Lim, P.T. Fine structure of the diatoms Thalassiosira and Coscinodiscus (Bacillariophyceae): Light and electron microscopy observation. Ann. Microsc. 2010, 10, 28–35. [Google Scholar]
- Hilaluddin, F.; Leaw, C.P.; Lim, P.T. Morphological observation of common pennate diatoms (Bacillariophyceae) from Sarawak estuarine waters. Ann. Microsc. 2011, 11, 12–23. [Google Scholar]
- Manoylov, K.M.; France, Y.E.; Geletu, A.; Dominy, J.N. Algal community membership of estuarine mudflats from the Savannah River, United States. J. Mar. Sci. Eng. 2016, 4, 11. [Google Scholar] [CrossRef]
- Stickley, C.E.; Pike, J.; Leventer, A.; Dunbar, R.; Domack, E.W.; Brachfeld, S.; Manley, P.; McClennan, C. Deglacial ocean and climate seasonality in laminated diatom sediments, Mac. Robertson Shelf, Antarctica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 227, 290–310. [Google Scholar] [CrossRef]
- Silva-Lehmkuhl, A.M.; Tremarin, P.I.; Vercellino, I.S.; Ludwig, T.A. Periphytic diatoms from an oligotrophic lentic system, Piraquara I reservoir, Paraná state, Brazil. Biota Neotrop. 2019, 19, e20180568. [Google Scholar] [CrossRef]
- Kafouris, S.; Smeti, E.; Spatharis, S.; Tsirtsis, G.; Economou-Amilli, A.; Danielidis, D.B. Nitrogen as the main driver of benthic diatom composition and diversity in oligotrophic coastal systems. Sci. Total Environ. 2019, 694, 133773. [Google Scholar] [CrossRef] [PubMed]
- Coenen, J.J.; Dodd, J.P.; Gruetzner, J.; Hall, I.R.; LeVay, L.J.; Lathika, N.; Jimenez, F.J.; Hemming, S.R.; Scherer, R.P. Intermediate water dynamics at the Indian-Atlantic Ocean Gateway during the Pliocene inferred from opal accumulation and diatom assemblages at IODP Site U1475. IODP Expedition 361 Scientists (2019). In Proceedings of the AGU Fall Meeting 2019, San Francisco, CA, USA, 9–13 December 2019. [Google Scholar]
- Hafner, D.; Car, A.; Jasprica, N.; Kapetanović, T.; Dupčić Radić, I. Relationship between marine epilithic diatoms and environmental variables in oligotrophic bay, NE Mediterranean. Mediterr. Mar. Sci. 2018, 19, 223–239. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Chen, M.; Lan, B.; Qi, H.; Zhang, A.; Lan, D.; Fang, Q. Diatom distribution as an environmental indicator in surface sediments of the West Philippine Basin. Chin. J. Oceanol. Limn. 2017, 35, 431–443. [Google Scholar] [CrossRef]
- Muda, A.; Ahmad, Z.M.A.; Lim, K.L. Sustainable management and conservation of the Matang mangroves. In Sustainable Management of Matang Mangroves 100 Years and Beyond; Shaharuddin, M.I., Muda, A., Ujang, R., Budin, K.A., Lim, K.L., Rosli, S., Jalil, M.S., Eds.; Forest Biodiversity Series, 4; Forestry Department Peninsular Malaysia: Kuala Lumpur, Malaysia, 2005; pp. 39–52. [Google Scholar]
- Ooi, A.L.; Chong, V.C. Larval fish assemblages in a tropical mangrove estuary and adjacent coastal waters: Offshore—inshore flux of marine and estuarine species. Cont. Shelf Res. 2011, 31, 1599–1610. [Google Scholar] [CrossRef]
- Chew, L.L.; Chong, V.C.; Tanaka, K.; Sasekumar, A. Phytoplankton fuel the energy flow from zooplankton to small nekton in turbid mangrove waters. Mar. Ecol. Prog. Ser. 2012, 469, 7–24. [Google Scholar] [CrossRef] [Green Version]
- Balqis, S.A.R.; Yusoff, F.M.; Nishikawa, J.; Lindsay, D.; Nishida, S. Influenced of environmental parameters on habitat preference of gelatinous zooplankton in various coastal ecosystems, the Straits of Malacca. Reg. Stud. Mar. Sci. 2019, 30, 100712. [Google Scholar] [CrossRef]
- Hou, W.; Lee, Z.; Weidemann, A.D. Why does the Secchi disk disappear? An imaging perspective. Opt. Express 2007, 15, 2791–2802. [Google Scholar] [CrossRef] [Green Version]
- Kulshreshtha, A.; Shanmugam, P. Estimation of Secchi transparency in turbid coastal waters. Aquat. Procedia 2015, 4, 1114–1118. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: Oxford, UK, 1984; p. 173. [Google Scholar]
- Ebina, J.; Tsutsui, T.; Shirai, T. Simultaneous determination of total nitrogen and total phosphorus in water using peroxodisulfate oxidation. Water Res. 1983, 17, 1721–1726. [Google Scholar] [CrossRef]
- LeGresley, M.; McDermott, G. Counting chamber methods for quantitative phytoplankton analysis-Haemocytometer, Palmer-Maloney cell and Sedgewick-Rafter cell. In Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis; Karlson, B., Cusack, C., Bresnan, E., Eds.; Intergovernmental Oceanographic Commission, UNESCO (IOC Manuals and Guides): Paris, France, 2010; pp. 25–30. [Google Scholar]
- Andersen, P.; Throndsen, J. Estimating cell numbers. In Manual on Harmful Marine Microalgae, 2nd ed.; Hallegreaff, G.M., Anderson, D.M., Cembella, A.D., Enevoldsen, H.O., Eds.; UNESCO: Paris, France, 2003; pp. 99–129. [Google Scholar]
- Omura, T.; Iwataki, M.; Borja, V.M.; Takayama, H.; Fukuyo, Y. Marine Phytoplankton of the Western Pacific; Kouseisha Kouseikaku: Tokyo, Japan, 2012; p. 160. [Google Scholar]
- Tomas, C.R. Identifying Marine Phytoplankton; Academic Press, an Imprint of Elsevier: San Diego, CA, USA, 1997; p. 858. [Google Scholar]
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E Ltd.: Plymouth, UK, 2014. [Google Scholar]
- Aké-Castillo, J.A.; Vázquez, G. Phytoplankton variation and its relation to nutrients and allochthonous organic matter in a coastal lagoon on the Gulf of Mexico. Estuar. Coast. Shelf Sci. 2008, 78, 705–714. [Google Scholar] [CrossRef]
- Rahaman, S.M.B.; Golder, J.; Rahaman, M.S.; Hasanuzzaman, A.F.M.; Huq, K.A.; Begum, S.; Islam, S.S.; Bir, J. Spatial and temporal variations in phytoplankton abundance and species diversity in the Sundarbans mangrove forest of Bangladesh. J. Mar. Sci. Res. Dev. 2013, 3, 1–9. [Google Scholar]
- Tanaka, K.; Choo, P.S. Influence of nutrient outwelling from the mangrove swamp on the distribution of phytoplankton in the Matang mangrove estuary, Malaysia. J. Oceanogr. 2000, 56, 69–78. [Google Scholar] [CrossRef]
- Manna, S.; Chaudhuri, K.; Bhattacharyya, S.; Bhattacharyya, M. Dynamics of Sundarban estuarine ecosystem: Eutrophication induced threat to mangroves. Saline Syst. 2010, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Thillai Rajasekar, K.T.; Perumal, P.; Santhanam, P. Phytoplankton diversity in the Coleroon estuary, southeast coast of India. J. Mar. Biol. Assoc. India 2005, 47, 127–132. [Google Scholar]
- Zhang, Y.; Peng, C.; Wang, J.; Huang, S.; Hu, Y.; Zhang, J.; Li, D. Temperature and silicate are significant driving factors for the seasonal shift of dominant diatoms in a drinking water reservoir. J. Oceanol. Limnol. 2019, 37, 568–579. [Google Scholar] [CrossRef]
- Donald, H.K.; Foster, G.L.; Fröhberg, N.; Swan, G.E.; Poulton, A.J.; Moore, C.M.; Humphreys, M.P. The pH dependency of the boron isotopic composition of diatom opal (Thalassiosira weissflogii). Biogesociences 2020, 17, 2825–2837. [Google Scholar] [CrossRef]
- Mejía, L.M.; Isensee, K.; Méndez-Vicente, A.; Pisonero, J.; Shimizu, N.; González, C.; Monteleone, B.; Stoll, H. B content and Si/C ratios from cultured diatoms (Thalassiosira pseudonana and Thalassiosira weissflogii): Relationship to seawater pH and diatom carbon acquisition. Geochim. Cosmochim. Acta 2013, 123, 322–337. [Google Scholar] [CrossRef] [Green Version]
- Khatoon, H.; Banerjee, S.; Yusoff, F.M.; Shariff, M. Effects of salinity on the growth and proximate composition of selected tropical marine periphytic diatoms and cyanobacteria. Aquac. Res. 2010, 41, 1348–1355. [Google Scholar] [CrossRef]
- Saifullah, A.S.M.; Abu Hena, M.K.; Idris, M.H.; Halimah, A.R.; Johan, I. Composition and diversity of phytoplankton from mangrove estuaries in Sarawak, Malaysia. J. Biol. Sci. 2014, 14, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Nursuhayati, A.S.; Yusoff, F.M.; Shariff, M. Spatial and temporal distribution of phytoplankton in Perak estuary, Malaysia, during monsoon season. J. Fish. Aquat. Sci. 2013, 8, 480–493. [Google Scholar]
- Potapova, M. Patterns of diatom distribution in relation to salinity. In The Diatom World; Seckbach, J., Kociolek, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 313–332. [Google Scholar]
- Rajkumar, M.; Perumal, P.; Ashok Prabu, V.; Vengadesh Perumal, N.; Thillai Rajasekar, K. Phytoplankton diversity in Pichavaram mangrove waters from south-east coast of India. J. Environ. Biol. 2009, 30, 489–498. [Google Scholar] [PubMed]
- Lewandowska, A.; Sommer, U. Climate change and the spring bloom: A mesocosm study on the influence of light and temperature on phytoplankton and mesozooplankton. Mar. Ecol. Prog. Ser. 2010, 405, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhao, L.; Zhu, C.; Wang, J.; Jiang, J.; Yang, S. Response of diatom community in Lugu Lake (Yunnan–Guizhou Plateau, China) to climate change over the past century. J. Paleolimnol. 2014, 51, 357–373. [Google Scholar] [CrossRef]
- Ge, J.; Torres, R.; Chen, C.; Liu, J.; Xu, Y.; Bellerby, R.; Shen, F.; Bruggeman, J.; Ding, P. Influence of suspended sediment front on nutrients and phytoplankton dynamics off the Changjiang Estuary: A FVCOM-ERSEM coupled model experiment. J. Mar. Syst. 2020, 204, 103292. [Google Scholar] [CrossRef]
- Domingues, R.B.; Anselmo, T.P.; Barbosa, A.B.; Sommer, U.; Galvão, H.M. Light as a driver of phytoplankton growth and production in the freshwater tidal zone of a turbid estuary. Estuar. Coast Shelf. Sci. 2011, 91, 526–535. [Google Scholar] [CrossRef]
- Shen, P.P.; Li, G.; Huang, L.M.; Zhang, J.L.; Tan, Y.H. Spatio-temporal variability of phytoplankton assemblages in the Pearl River estuary, with special reference to the influence of turbidity and temperature. Cont. Shelf. Res. 2011, 31, 1672–1681. [Google Scholar] [CrossRef]
- Fan, C.; Glibert, P.M. Effects of light on nitrogen and carbon uptake during a Prorocentrum minimum bloom. Harmful Algae 2005, 4, 629–641. [Google Scholar] [CrossRef]
- Matsubara, T.; Nagasoe, S.; Yamasaki, Y.; Shikata, T.; Shimasaki, Y.; Oshima, Y.; Honjo, T. Effects of temperature, salinity, and irradiance on the growth of the dinoflagellate Akashiwo sanguinea. J. Exp. Mar. Biol. Ecol. 2007, 342, 226–230. [Google Scholar] [CrossRef]
- Yuan, Y.; Jiang, M.; Liu, X.; Yu, H.; Otte, M.L.; Ma, C.; Her, Y.G. Environmental variables influencing phytoplankton communities in hydrologically connected aquatic habitats in the Lake Xingkai basin. Ecol. Indic. 2018, 91, 1–12. [Google Scholar] [CrossRef]
- Sgro, G.V.; Reavie, E.D. Fossil diatoms, geochemistry, and the Anthropocene paleolimnology of Lake Huron. J. Great Lakes Res. 2018, 44, 765–778. [Google Scholar] [CrossRef]
- Zhang, X.; Reed, J.M.; Lacey, J.H.; Francke, A.; Leng, M.J.; Levkov, Z.; Wagner, B. Complexity of diatom response to Lateglacial and Holocene climate and environmental change in ancient, deep and oligotrophic Lake Ohrid (Macedonia and Albania). Biogeosciences 2016, 13, 1351–1365. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Das, S.; Chakraborty, S.; De, T.K. Study on mangrove associated estuarine waters of Northeastern Bay of Bengal reveals potential diatom indicators of dissolved inorganic compounds. Braz. J. Biol. Sci. 2015, 2, 155–168. [Google Scholar]
- Canini, N.D.; Metillo, E.B.; Azanza, R.V. Monsoon-influenced phytoplankton community structure in a Philippine mangrove estuary. Trop. Ecol. 2013, 54, 331–343. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Mean ± SE | 1. | 2. | 3. | 4. | 5. | 6. | 7. | 8. | 9. | 10. | 11. | 12. | 13. | 14. | 15. | 16. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Temp. (°C) | 30.24 ± 0.21 | 1 | |||||||||||||||
| 2. DO (mg L−1) | 5.19 ± 0.13 | 0.48 | 1 | ||||||||||||||
| 3. Sal. (PSU) | 23.85 ± 0.40 | 0.14 | −0.00 | 1 | |||||||||||||
| 4. pH | 7.41 ± 0.07 | −0.41 | 0.02 | 0.34 | 1 | ||||||||||||
| 5. TDS (g L−1) | 24.61 ± 0.22 | −0.18 | −0.09 | 0.70 | 0.13 | 1 | |||||||||||
| 6. Turb. (NTU) | 48.60 ± 12.12 | 0.39 | −0.27 | 0.10 | −0.30 | 0.04 | 1 | ||||||||||
| 7. TN (μg L−1) | 144.73 ± 40.19 | 0.24 | −0.42 | 0.17 | −0.37 | 0.02 | 0.59 | 1 | |||||||||
| 8. TP (μg L−1) | 159.68 ± 23.61 | 0.23 | −0.39 | 0.07 | −0.24 | −0.00 | 0.89 | 0.58 | 1 | ||||||||
| 9. TAN (μg L−1) | 18.45 ± 1.82 | 0.31 | 0.41 | −0.34 | −0.13 | −0.38 | −0.22 | −0.59 | −0.32 | 1 | |||||||
| 10. NO-N (μg L−1) | 52.69 ± 14.03 | −0.41 | −0.12 | −0.80 | −0.38 | −0.18 | −0.19 | −0.16 | −0.16 | 0.01 | 1 | ||||||
| 11. SRP (μg L−1) | 42.46 ± 4.71 | −0.40 | −0.48 | −0.29 | −0.43 | −0.02 | −0.05 | 0.46 | 0.06 | −0.51 | 0.54 | 1 | |||||
| 12. Phyto. (cell L−1) | 288,813.89 ± 104,249.51 | −0.03 | −0.41 | 0.28 | −0.22 | 0.06 | 0.07 | 0.69 | 0.15 | −0.64 | −0.15 | 0.69 | 1 | ||||
| 13. Diatom (cell L−1) | 270,986.11 ± 104,492.24 | −0.06 | −0.42 | 0.29 | −0.21 | 0.09 | 0.07 | 0.69 | 0.14 | −0.66 | −0.15 | 0.69 | 0.99 | 1 | |||
| 14. Dino. (cell L−1) | 16,679.86 ± 3664.83 | 0.69 | 0.42 | −0.29 | −0.28 | −0.59 | 0.22 | 0.10 | 0.15 | 0.35 | −0.16 | −0.25 | −0.06 | −0.10 | 1 | ||
| 15. Rainfall (mm) | 347.70 ± 23.32 | 0.40 | 0.63 | −0.37 | −0.02 | −0.43 | −0.31 | −0.58 | −0.40 | 0.80 | 0.06 | −0.62 | −0.64 | −0.65 | 0.44 | 1 | |
| 16. Tidal level (m) | 2.28 ± 0.05 | −0.46 | −0.54 | 0.20 | 0.11 | −0.10 | 0.37 | 0.07 | 0.50 | 0.16 | −0.77 | 0.83 | 0.72 | 0.74 | −0.44 | −0.93 | 1 |
| Phytoplankton | Southwest | Inter-Monsoon 1 | Northeast | Inter-Monsoon 2 |
|---|---|---|---|---|
| TN:TP = 3 | TN:TP = 37 | TN:TP = 10 | TN:TP = 8 | |
| Diatom (Bacillariophyta) | 92.08% | 98.90% | 66.04% | 43.68% |
| Dinoflagellates (Dinophyta) | 7.21% | 0.98% | 33.10% | 52.23% |
| Others group | 0.70% | 0.12% | 0.86% | 4.09% |
| No. of species | 64 | 34 | 54 | 53 |
| F1 | F2 | F3 | |
|---|---|---|---|
| Eigenvalue | 0.27 | 0.14 | 0.02 |
| % of constrained inertia | 60.20 | 32.42 | 4.85 |
| % of cumulative | 60.20 | 92.62 | 97.47 |
| Total inertia | 59.19 | 31.88 | 4.77 |
| % of inertia cumulative | 59.19 | 91.07 | 93.83 |
| Cyclotella sp. | 0.59 | 0.41 | 0.002 |
| Coscinodiscus sp. | 0.83 | 0.001 | 0.04 |
| Skeletonema costatum | 0.99 | 0.002 | 0.01 |
| Pleurosigma sp. | 0.89 | 0.003 | 0.06 |
| Nitzschia longissima | 0.34 | 0.32 | 0.33 |
| Thalassionema nitzschioides | 0.40 | 0.16 | 0.00 |
| Total phytoplankton | 0.64 | 0.34 | 0.004 |
| Total diatom | 0.34 | 0.47 | 0.17 |
| Total dinoflagellates | 0.51 | 0.45 | 0.04 |
| Temperature | −0.07 | 0.85 | −0.02 |
| Dissolved oxygen | −0.39 | 0.14 | −0.16 |
| Salinity | 0.33 | −0.24 | 0.33 |
| Conductivity | 0.29 | -0.19 | 0.37 |
| pH | −0.65 | −0.61 | 0.009 |
| Total dissolved solids | 0.21 | -0.09 | 0.34 |
| Turbidity | 0.17 | 0.43 | 0.50 |
| Total nitrogen | 0.53 | 0.11 | 0.07 |
| Total phosphorus | 0.12 | 0.11 | 0.35 |
| Total ammonium nitrogen | −0.83 | 0.17 | −0.07 |
| Nitrate + nitrite | 0.18 | 0.34 | −0.24 |
| Soluble reactive phosphorus | 0.89 | 0.007 | −0.14 |
| Rainfall | −0.94 | 0.17 | −0.10 |
| Tidal level (m) | 0.98 | −0.11 | −0.02 |
| Source | Phytoplankton | Diatoms | Dinoflagellates | |
|---|---|---|---|---|
| Statistics | df | 3.20 | 3.20 | 3.20 |
| MS | 3487.7 | 2550.7 | 5205.7 | |
| F | 6.28 | 6.31 | 6.36 | |
| P | <0.001 | <0.001 | <0.001 | |
| Pairwise test, P | Southwest, IM1 | 0.0022 | 0.0029 | 0.002 |
| Southwest, Northeast | 0.0053 | 0.0035 | 0.0003 | |
| Southwest, IM2 | 0.0053 | 0.0033 | 0.0029 | |
| IM1, Northeast | 0.0004 | 0.0006 | 0.0527 | |
| IM1, IM2 | 0.0289 | 0.026 | 0.0263 | |
| Northeast, IM2 | 0.0069 | 0.0772 | 0.028 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hilaluddin, F.; Yusoff, F.M.; Toda, T. Shifts in Diatom Dominance Associated with Seasonal Changes in an Estuarine-Mangrove Phytoplankton Community. J. Mar. Sci. Eng. 2020, 8, 528. https://doi.org/10.3390/jmse8070528
Hilaluddin F, Yusoff FM, Toda T. Shifts in Diatom Dominance Associated with Seasonal Changes in an Estuarine-Mangrove Phytoplankton Community. Journal of Marine Science and Engineering. 2020; 8(7):528. https://doi.org/10.3390/jmse8070528
Chicago/Turabian StyleHilaluddin, Fareha, Fatimah Md. Yusoff, and Tatsuki Toda. 2020. "Shifts in Diatom Dominance Associated with Seasonal Changes in an Estuarine-Mangrove Phytoplankton Community" Journal of Marine Science and Engineering 8, no. 7: 528. https://doi.org/10.3390/jmse8070528
APA StyleHilaluddin, F., Yusoff, F. M., & Toda, T. (2020). Shifts in Diatom Dominance Associated with Seasonal Changes in an Estuarine-Mangrove Phytoplankton Community. Journal of Marine Science and Engineering, 8(7), 528. https://doi.org/10.3390/jmse8070528