Spatial and Temporal Variability of Ice Algal Trophic Markers—With Recommendations about Their Application

 , , , , and
, , , , and
Abstract
:1. Introduction
1.1. Fatty Acid Trophic Markers (FATM)
1.2. Highly Branched Isoprenoids (HBIs)
1.3. Carbon Stable Isotope Ratios (δ13C)
2. Materials and Methods
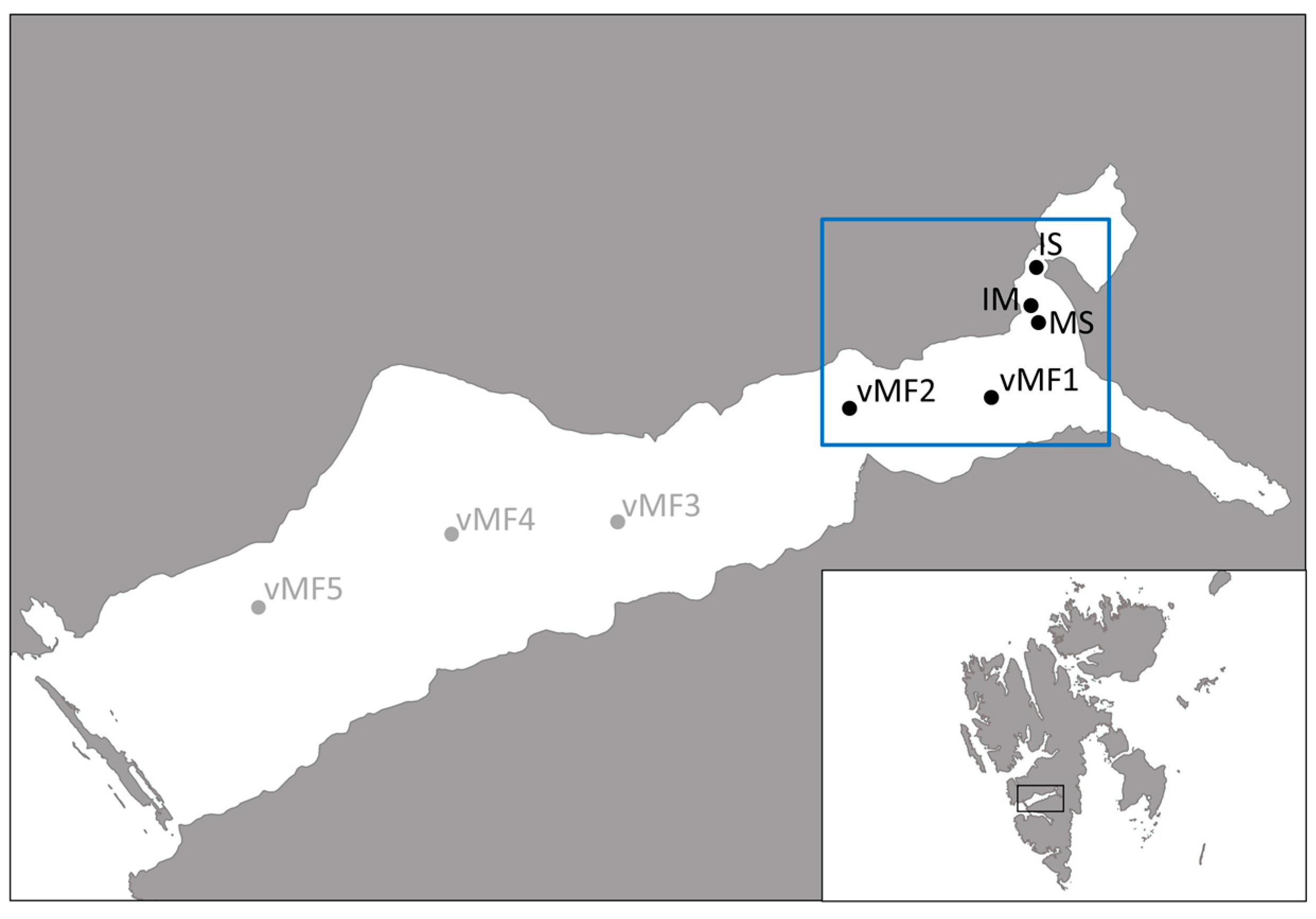
2.1. Site Description
2.2. Sampling
2.3. Organic Carbon Analysis
2.4. Lipid Analysis
2.5. Bulk Stable Isotope Analysis
2.6. Compound-Specific Isotope Analysis of Fatty Acids
2.7. Sea Ice Algae Taxonomy
2.8. Determination of Inorganic Nutrient Concentrations
2.9. Determination of Total Dissolved Inorganic Carbon
2.10. Determination of Chlorophyll a (Chl a)
2.11. Statistical Analysis
3. Results
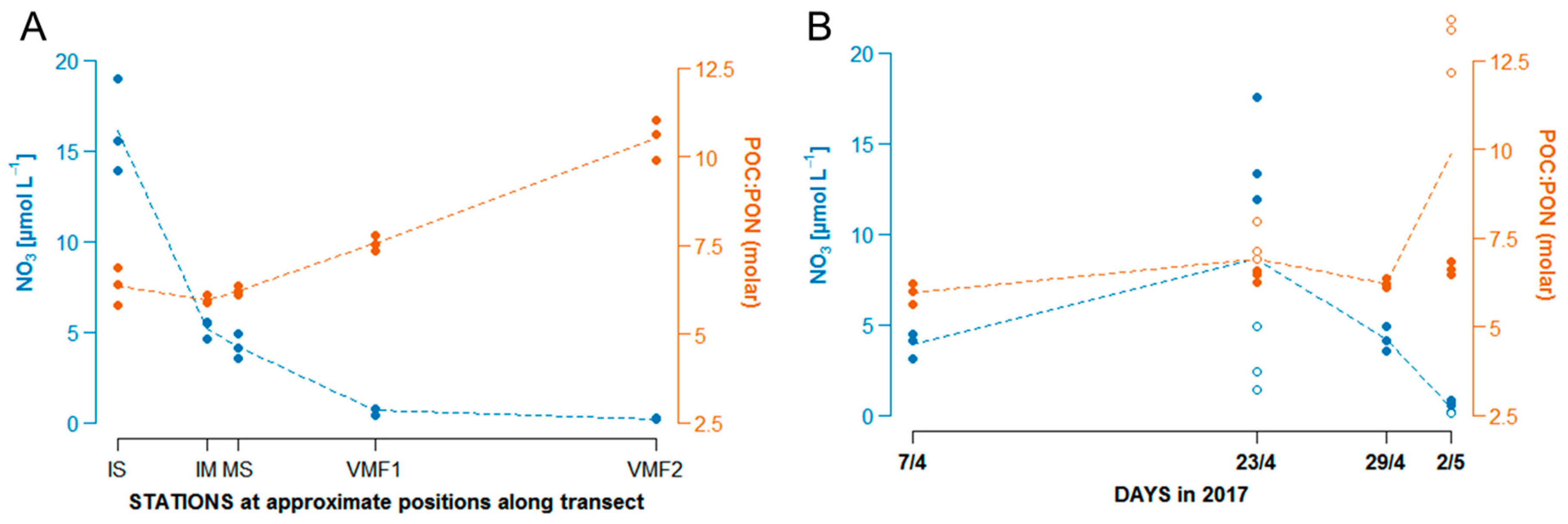
3.1. General Physical Situation, Development of Chl a, Nutrients and C:N Ratios along the Transect and over Time
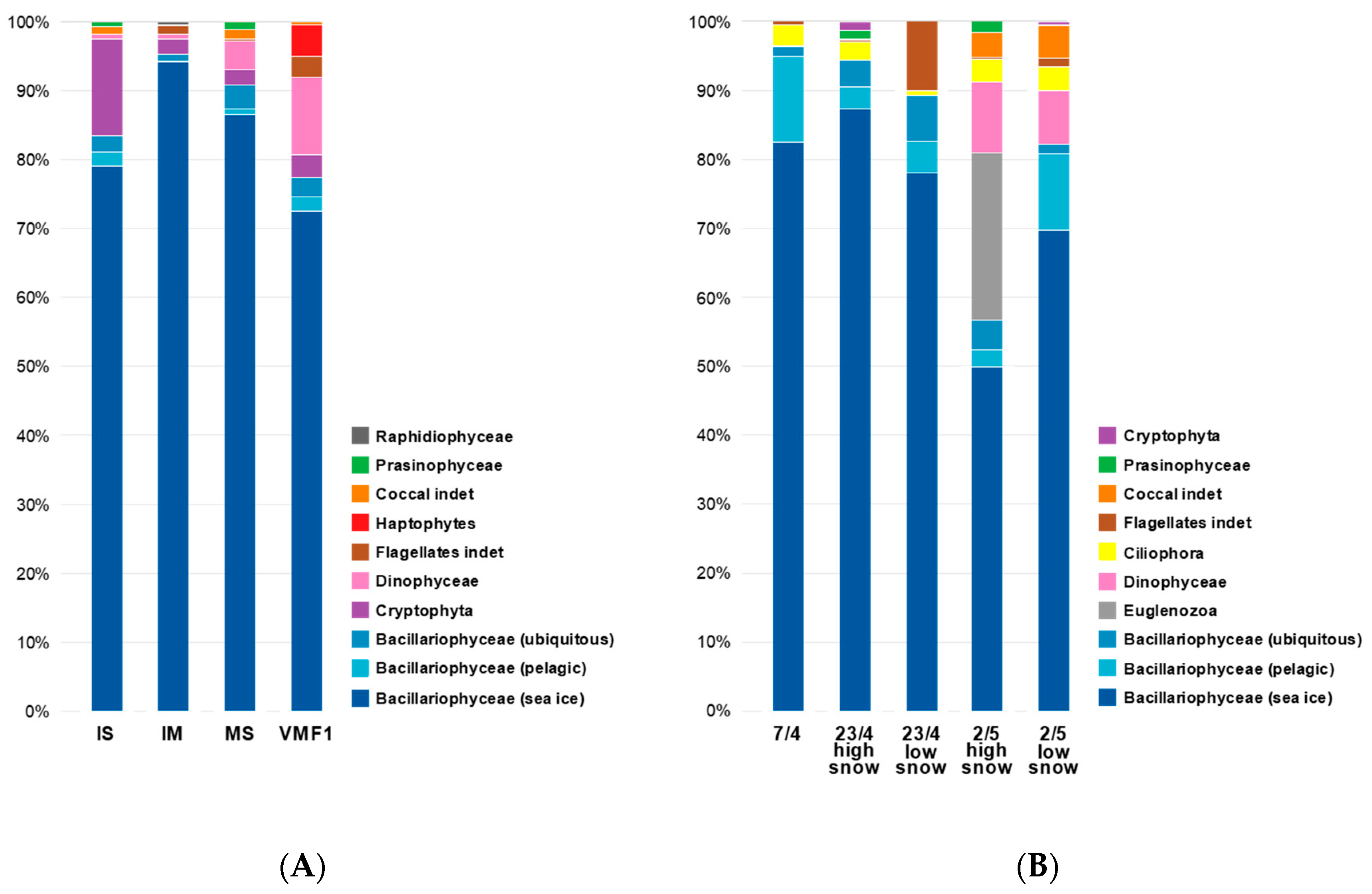
3.2. Species Composition
3.3. Variability of Ice Algal Biomarkers on Short Spatial and Temporal Scales
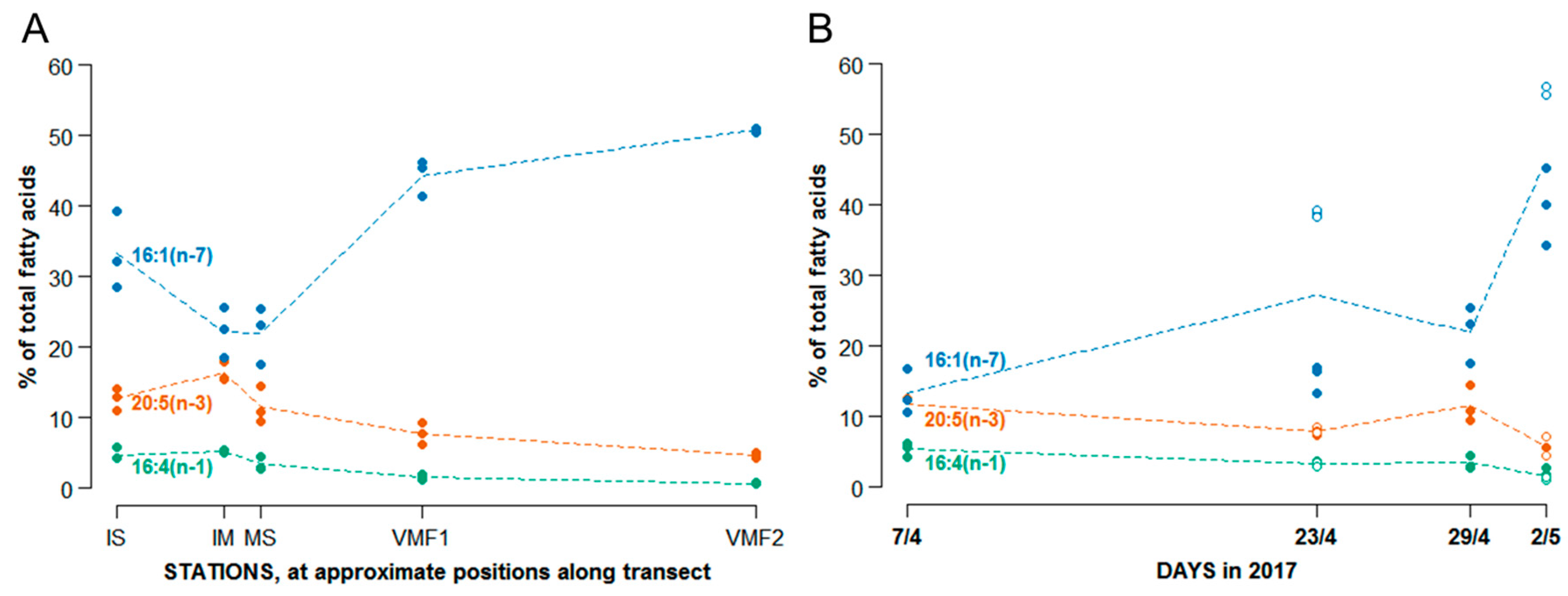
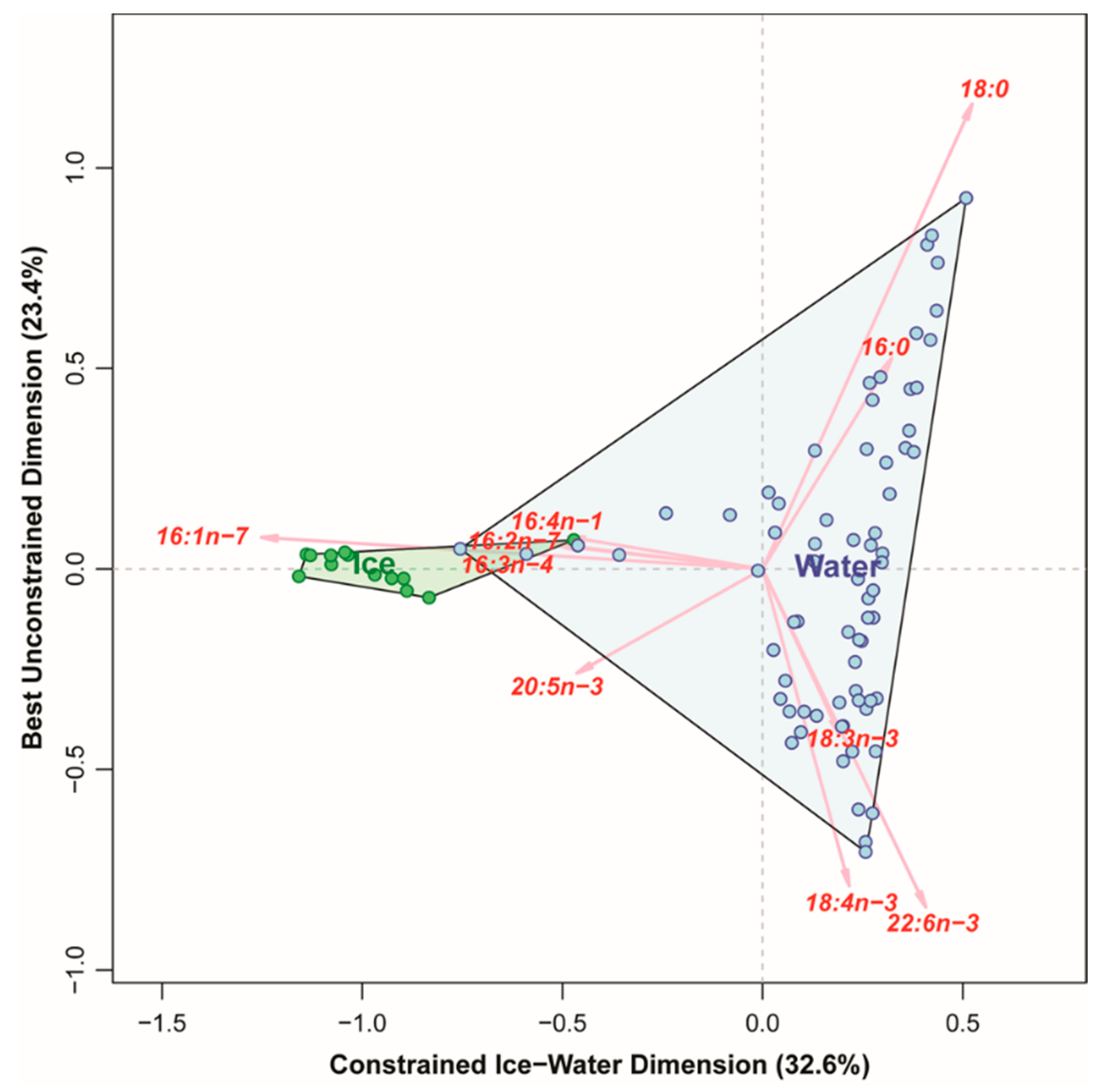
3.3.1. FATM
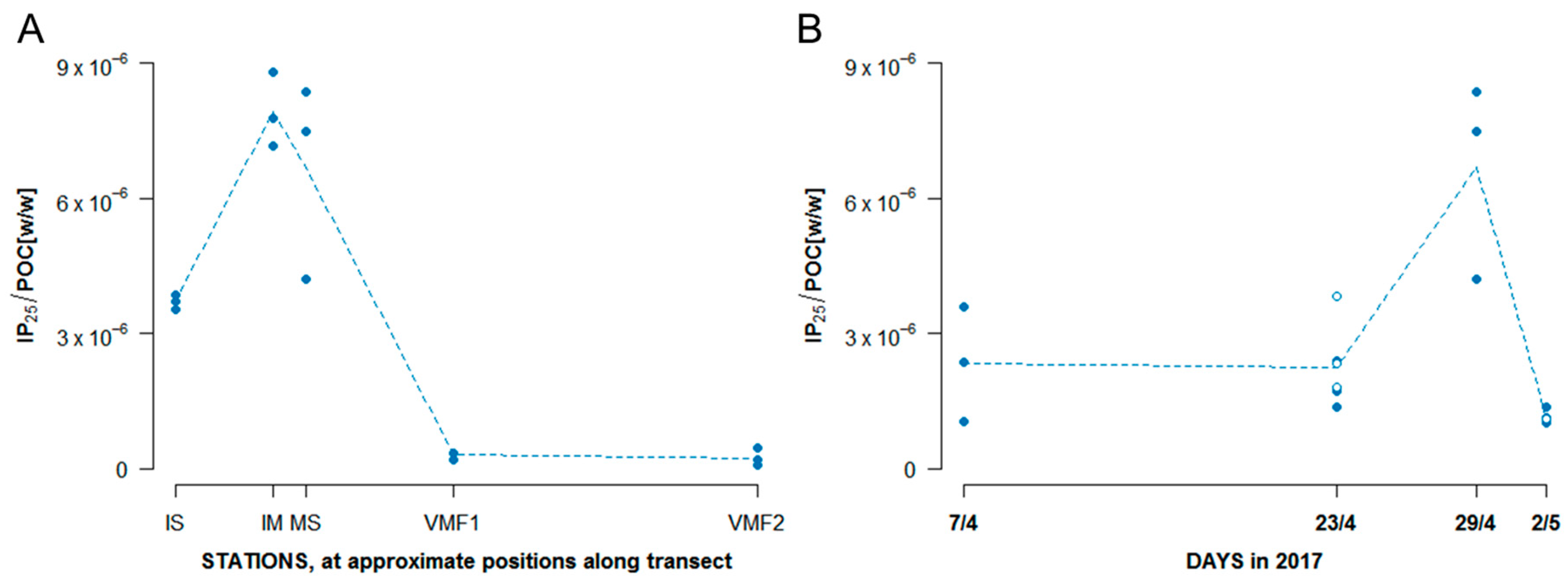
3.3.2. HBIs
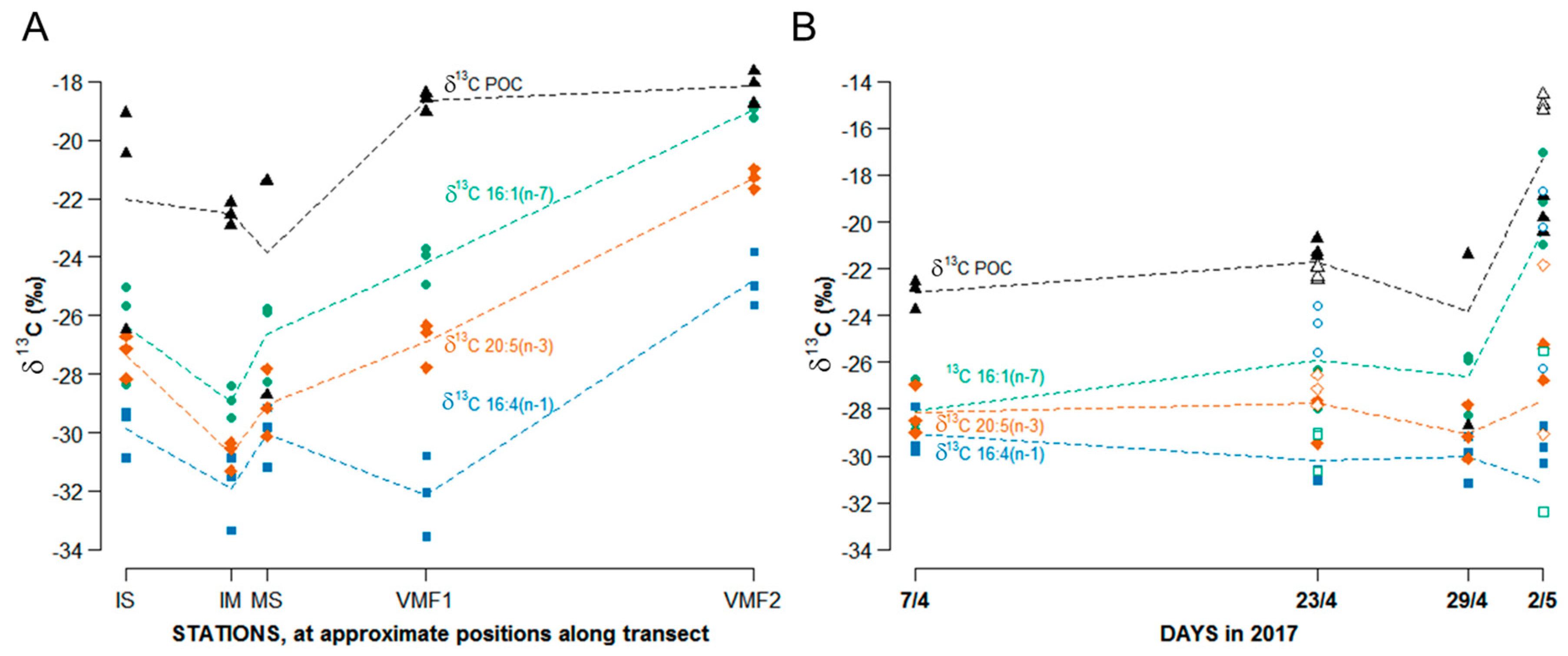
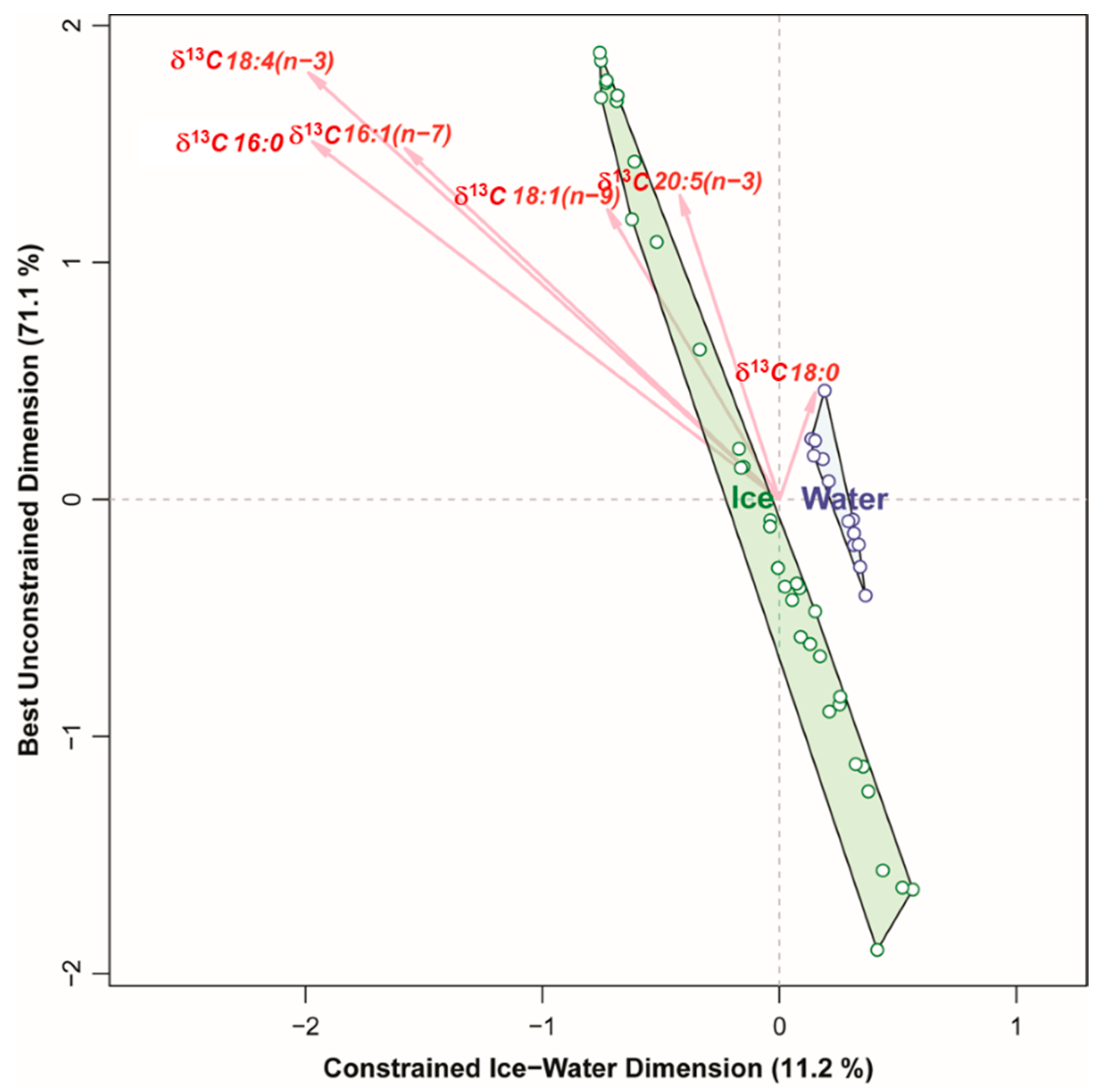
3.3.3. Stable Isotope Ratios
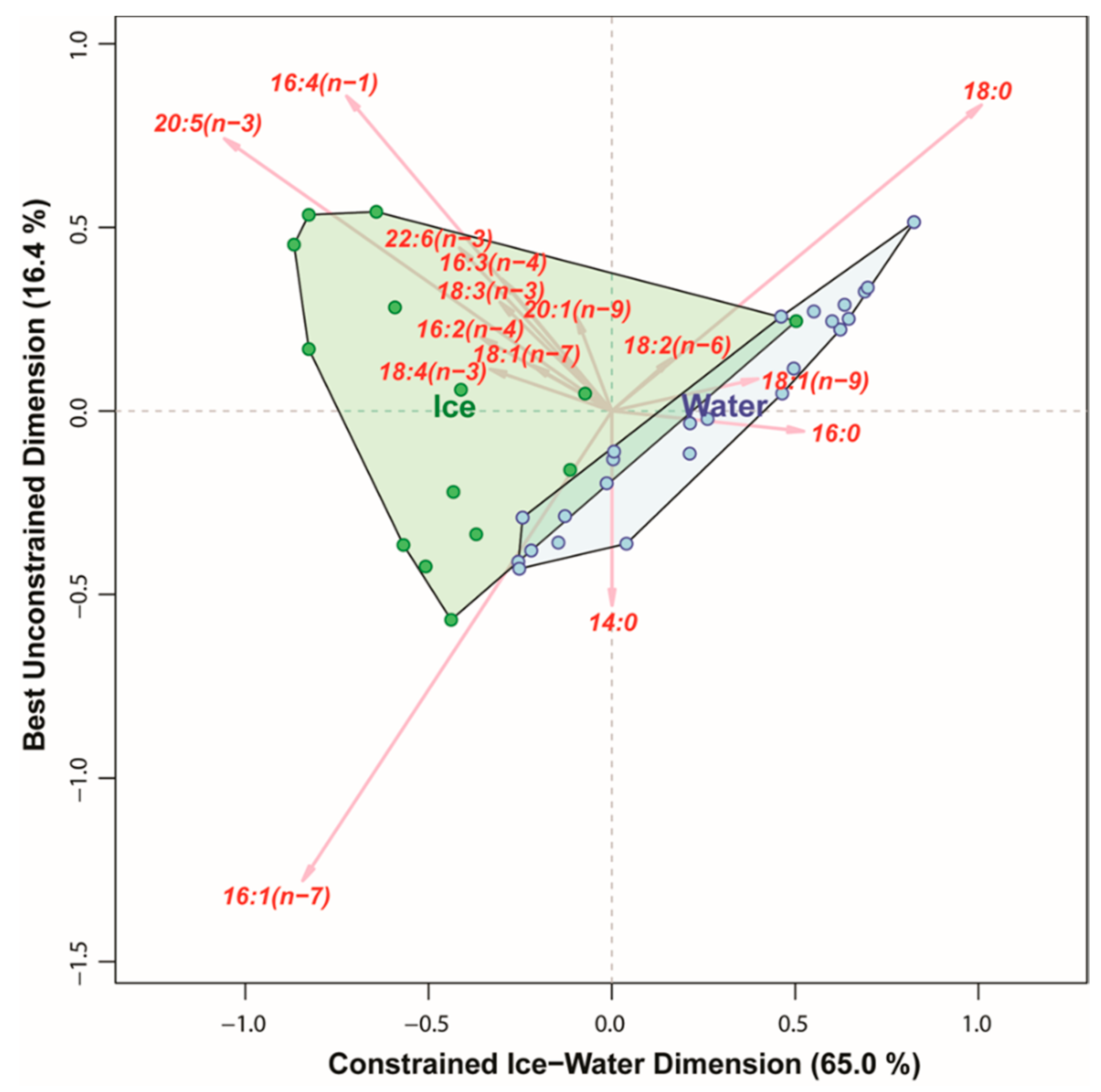
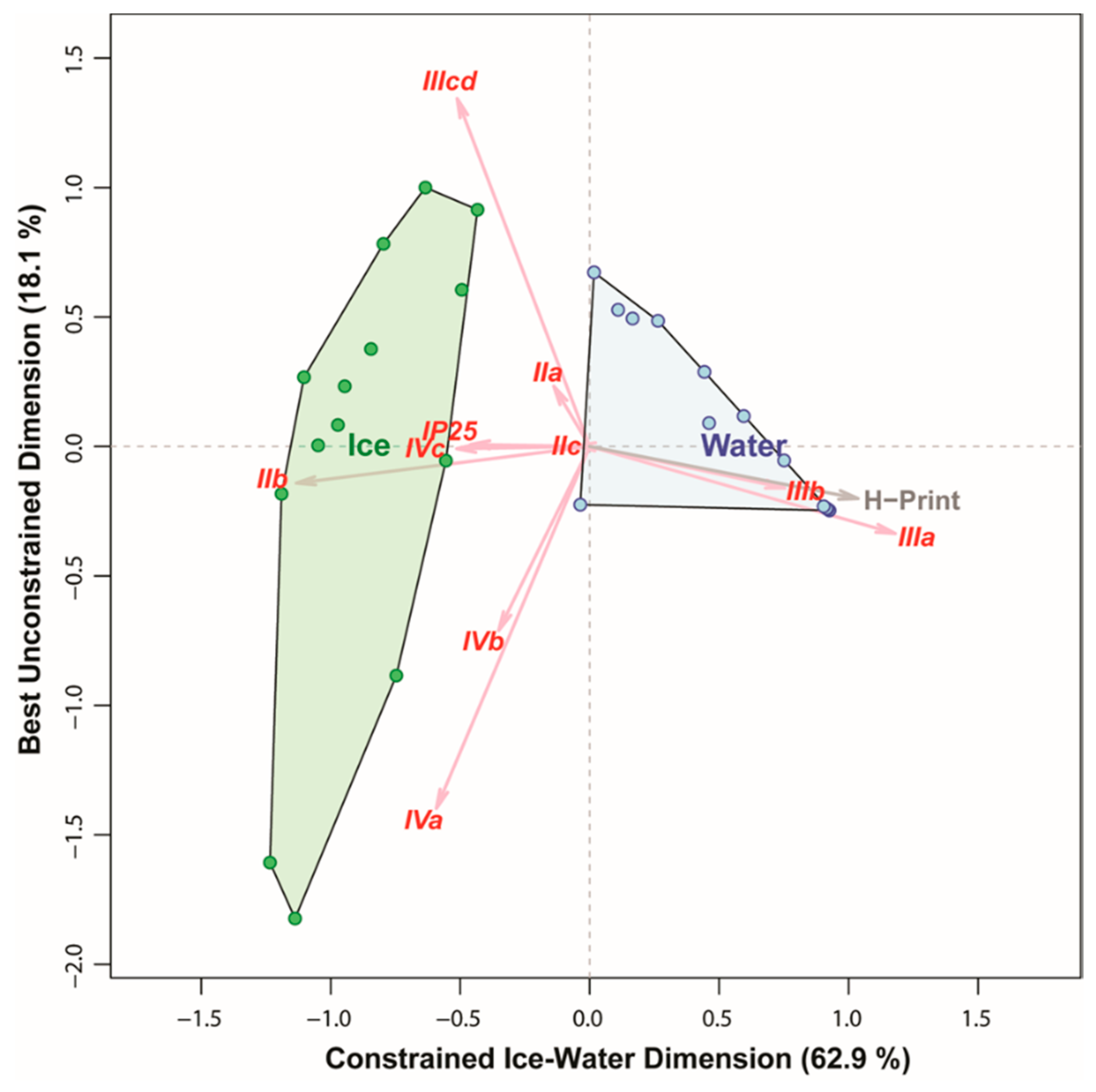
3.4. Comparing the Ability of Different Markers to Distinguish between Algae Collected in Sea Ice vs. Water Column
4. Discussion
4.1. Variability of Trophic Markers—And How This Relates to Environmental Conditions
4.2. Fatty Acid Trophic Markers (FATM)
4.3. Highly Branched Isoprenoids (HBIs)
4.4. Stable Isotope Ratios in Bulk POM and Specific Fatty Acids
4.5. Implications for Inferring Ice Algal Bloom Characteristics from Different Trophic Markers
- FATM: It is difficult to arrive at consistent conclusions based solely on single fatty acid trophic markers as the relative contributions of the three most widely used diatom marker FAs changed strongly, with partly contrasting patterns. Highest diatom contributions were indicated by 16:1(n-7) at VMF1 and VMF2, whereas typical diatom PUFAs (16:4(n-1) and 20:5(n-3)) were highest at the shallower stations. Summarizing total diatom FAs normalized to POC yielded similar values for IS, IM, and VMF1 (appx. 0.012), with MS being lower (0.005) and VMF2 much higher (0.039). Inferences based on individual %fatty acids only are inconclusive, total diatom FAs normalized to POC indicate highest diatom contributions at VMF2, and lowest at MS.
- δ13C POM: at stations VMF1 and VMF2 depleted ratios (−19 to −18‰) were reported that are typically considered to be indicative for sea ice algae, whereas MS and IM exhibited δ13C values that are normally considered to be of pelagic origin (−23 to −22‰). Based on these results, we would assume highest relative contributions of sea ice algae at VMF1 and VMF2, and more “pelagic” influenced species composition in bottom sea ice at the shallower stations.
- δ13C of single FAs: Isotope ratios of single fatty acids varied similar to those of bulk POM, with the exception of VMF1 that in terms of δ13C 16:4(n-1) was not significantly different from MS, IM and IS, while in all other FAs, the isotopic signal at this station was between that of VMF2 and the remaining shallower stations. Based on these results, we would assume highest relative contributions of sea ice algae at VMF1 and VMF2, and more “pelagic” influenced species at the shallower stations.
- IP25.: the highest concentration of this ice algal biomarker (both in absolute terms, and normalized to POC) was clearly found at the shallowest stations, and—despite some variability—at MS. Almost no IP25 could be detected at VMF1 and VMF2, which indicates the absence (or extremely low abundances) of IP25 producing sea ice algae. Based on these findings, one would conclude that no sea ice diatoms (or, more specifically, IP25 producers) were present at VMF1 and VMF2, whereas IS and MS clearly had sea ice diatom assemblages.
4.6. Comparing Trophic Markers’ Potential to Distinguish Reliably between Sea Ice Algae and Phytoplankton
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arrigo, K.R.; van Dijken, G.; Pabi, S. Impact of a shrinking Arctic ice cover on marine primary production. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef]
- Huntington, H.P.; Danielson, S.L.; Wiese, F.K.; Baker, M.; Boveng, P.; Citta, J.J.; Robertis, A.D.; Dickson, D.M.S.; Farley, E.; George, J.C.; et al. Evidence suggests potential transformation of the Pacific Arctic ecosystem is underway. Nat. Clim. Chang. 2020, 10, 342–348. [Google Scholar] [CrossRef]
- Grebmeier, J.M.; Overland, J.E.; Moore, S.E.; Farley, E.V.; Carmack, E.C.; Cooper, L.W.; Frey, K.E.; Helle, J.H.; McLaughlin, F.A.; McNutt, S.L. A major ecosystem shift in the northern Bering Sea. Science 2006, 311, 1461–1464. [Google Scholar] [CrossRef] [PubMed]
- Soreide, J.E.; Falk-Petersen, S.; Hegseth, E.N.; Hop, H.; Carroll, M.L.; Hobson, K.A.; Blachowiak-Samolyk, K. Seasonal feeding strategies of Calanus in the high-Arctic Svalbard region. Deep Sea Res. Part II-Top. Stud. Oceanogr. 2008, 55, 2225–2244. [Google Scholar] [CrossRef]
- Brown, T.A.; Belt, S.T.; Piepenburg, D. Evidence for a pan-Arctic sea-ice diatom diet in Strongylocentrotus spp. Polar Biol. 2012, 35, 1281–1287. [Google Scholar] [CrossRef]
- Brown, T.A.; Galicia, M.P.; Thiemann, G.W.; Belt, S.T.; Yurkowski, D.J.; Dyck, M.G. High contributions of sea ice derived carbon in polar bear (Ursus maritimus) tissue. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [Green Version]
- Budge, S.M.; Wooller, M.J.; Springer, A.M.; Iverson, S.J.; McRoy, C.P.; Divoky, G.J. Tracing carbon flow in an arctic marine food web using fatty acid-stable isotope analysis. Oecologia 2008, 157, 117–129. [Google Scholar] [CrossRef]
- Søreide, J.E.; Hop, H.; Carroll, M.; Falk-Petersen, S.; Hegseth, E.N. Seasonal food web structures and sympagic-pelagic coupling in the European Arctic revealed by stable isotopes and a two-source food web model. Prog. Oceanogr. 2006, 71, 59–87. [Google Scholar] [CrossRef]
- Kohlbach, D.; Ferguson, S.H.; Brown, T.A.; Michel, C. Landfast sea ice-benthic coupling during spring and potential impacts of system changes on food web dynamics in Eclipse Sound, Canadian Arctic. Mar. Ecol. Prog. Ser. 2019, 627, 33–48. [Google Scholar] [CrossRef] [Green Version]
- Tamelander, T.; Renaud, P.E.; Hop, H.; Carroll, M.L.; Ambrose, W.G.; Hobson, K.A. Trophic relationships and pelagic-benthic coupling during summer in the Barents Sea Marginal Ice Zone, revealed by stable carbon and nitrogen isotope measurements. Mar. Ecol. Prog. Ser. 2006, 310, 33–46. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H. Fatty acid profiles and production in marine phytoplankton. Mar. Drugs 2019, 17, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalsgaard, J.; St John, M.; Kattner, G.; Muller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [PubMed]
- Reitan, K.I.; Rainuzzo, J.R.; Olsen, Y. Effect of nutrient limitation on fatty acid and lipid content of marine microalgae. J. Phycol. 1994, 30, 972–979. [Google Scholar] [CrossRef]
- Thompson, P.A.; Guo, M.; Harrison, P.J. The influence of irradiance on the biochemical composition of 3 phytoplankton species and their nutritional value for larvae of the pacific oyster (Crassostrea gigas). Mar. Biol. 1993, 117, 259–268. [Google Scholar] [CrossRef]
- Thompson, P.A.; Guo, M.X.; Harrison, P.J.; Whyte, J.N.C. Effects of variation in temperature on the fatty acid composition of 8 species of marine phytoplankton. J. Phycol. 1992, 28, 488–497. [Google Scholar] [CrossRef]
- Leu, E.; Wiktor, J.; Soreide, J.; Berge, J.; Falk-Petersen, S. Increased irradiance reduces food quality of sea ice algae. Mar. Ecol. Prog. Ser. 2010, 411, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Leu, E.; Falk-Petersen, S.; Kwaśniewski, S.; Wulff, A.; Edvardsen, K.; Hessen, D.O. Fatty acid dynamics during the spring bloom in a High Arctic fjord: Importance of abiotic factors versus community changes. Can. J. Fish. Aquat. Sci. 2006, 63, 2760–2779. [Google Scholar] [CrossRef]
- Leu, E.; Wängberg, S.-Å.; Wulff, A.; Falk-Petersen, S.; Borre Orbaek, J.; Hessen, D.O. Effects of changes in ambient PAR and UV radiation on the nutritional quality of an Arctic diatom (Thalassiosira antarctica var. borealis). J. Exp. Mar. Biol. Ecol. 2006, 337, 65–81. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.A.; Belt, S.T.; Tatarek, A.; Mundy, C.J. Source identification of the Arctic sea ice proxy IP25. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef]
- Belt, S.T.; Masse, G.; Rowland, S.J.; Poulin, M.; Michel, C.; LeBlanc, B. A novel chemical fossil of palaeo sea ice: IP25. Organ. Geochem. 2007, 38, 16–27. [Google Scholar] [CrossRef]
- Volkman, J.K.; Barrett, S.M.; Dunstan, G.A. C25 and C30 highly branched isoprenoid alkenes in laboratory cultures of two marine diatoms. Organ. Geochem. 1994, 21, 407–413. [Google Scholar] [CrossRef]
- Limoges, A.; Massé, G.; Weckström, K.; Poulin, M.; Ellegaard, M.; Heikkilä, M.; Geilfus, N.-X.; Sejr, M.K.; Rysgaard, S.; Ribeiro, S. Spring succession and vertical export of diatoms and IP25 in a seasonally ice-covered high Arctic Fjord. Front. Earth Sci. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Amiraux, R.; Smik, L.; Köseoğlu, D.; Rontani, J.-F.; Galindo, V.; Grondin, P.-L.; Babin, M.; Belt, S.T. Temporal evolution of IP25 and other highly branched isoprenoid lipids in sea ice and the underlying water column during an Arctic melting season. Elem. Sci. Anth. 2019, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.A.; Belt, S.T.; Philippe, B.; Mundy, C.J.; Masse, G.; Poulin, M.; Gosselin, M. Temporal and vertical variations of lipid biomarkers during a bottom ice diatom bloom in the Canadian Beaufort Sea: Further evidence for the use of the IP25 biomarker as a proxy for spring Arctic sea ice. Polar Biol. 2011, 34, 1857–1868. [Google Scholar] [CrossRef]
- Brown, T.A.; Belt, S.T. Biomarker-based H-Print quantifies the composition of mixed sympagic and pelagic algae consumed by Artemia sp. J. Exp. Mar. Biol. Ecol. 2017, 488, 32–37. [Google Scholar] [CrossRef]
- Brown, T.A.; Yurkowski, D.J.; Ferguson, S.H.; Alexander, C.; Belt, S.T. H-Print: A new chemical fingerprinting approach for distinguishing primary production sources in Arctic ecosystems. Environ. Chem. Lett. 2014, 12, 387–392. [Google Scholar] [CrossRef]
- Brown, T.A.; Rad-Menéndez, C.; Ray, J.L.; Skaar, K.S.; Thomas, N.; Ruiz-Gonzalez, C.; Leu, E. Influence of nutrient availability on Arctic sea ice diatom HBI lipid synthesis. Organ. Geochem. 2020, 141, 103977. [Google Scholar] [CrossRef]
- Pineault, S.; Tremblay, J.-E.; Gosselin, M.; Thomas, H.; Shadwick, E. The isotopic signature of particulate organic C and N in bottom ice: Key influencing factors and applications for tracing the fate of ice-algae in the Arctic Ocean. J. Geophys. Res. Oceans 2013, 118, 287–300. [Google Scholar] [CrossRef] [Green Version]
- de la Vega, C.; Jeffreys, R.M.; Tuerena, R.; Ganeshram, R.; Mahaffey, C. Temporal and spatial trends in marine carbon isotopes in the Arctic Ocean and implications for food web studies. Glob. Chang. Biol. 2019, 25, 4116–4130. [Google Scholar] [CrossRef] [Green Version]
- Tamelander, T.; Kivimäe, C.; Bellerby, R.G.J.; Renaud, P.E.; Kristiansen, S. Base-line variations in stable isotope values in an Arctic marine ecosystem: Effects of carbon and nitrogen uptake by phytoplankton. Hydrobiologia 2009, 630, 63–73. [Google Scholar] [CrossRef]
- Wang, S.W.; Budge, S.M.; Gradinger, R.R.; Iken, K.; Wooller, M.J. Fatty acid and stable isotope characteristics of sea ice and pelagic particulate organic matter in the Bering Sea: Tools for estimating sea ice algal contribution to Arctic food web production. Oecologia 2014, 174, 699–712. [Google Scholar] [CrossRef]
- Hoyland, K.V. Ice thickness, growth and salinity in Van Mijenfjorden, Svalbard, Norway. Polar Res. 2009, 28, 339–352. [Google Scholar] [CrossRef]
- Osuch, M.; Wawrzyniak, T. Inter- and intra-annual changes in air temperature and precipitation in western Spitsbergen. Int. J. Climatol. 2017, 37, 3082–3097. [Google Scholar] [CrossRef]
- Gosselin, M.; Legendre, L.; Therriault, J.C.; Demers, S.; Rochet, M. Physical control of the horizontal patchiness of sea-ice microalgae. Mar. Ecol. Prog. Ser. 1986, 29, 289–298. [Google Scholar] [CrossRef]
- Garrison, D.L.; Buck, K.R. Organism losses during ice melting: A serious bias in sea ice community studies. Polar Biol. 1986, 6, 237–239. [Google Scholar] [CrossRef]
- Brown, T.A. Stability of the lipid biomarker H-Print within preserved animals. Polar Biol. 2018, 41, 1901–1905. [Google Scholar] [CrossRef]
- Boissonnot, L.; Niehoff, B.; Hagen, W.; Søreide, J.E.; Graeve, M. Lipid turnover reflects life-cycle strategies of small-sized Arctic copepods. J. Plankton Res. 2016. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton Methodik. Mitt. Int. Ver. Limnol. 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Olenina, I. Biovolumes and Size-Classes of Phytoplankton in the Baltic Sea. Available online: https://epic.awi.de/id/eprint/30141/ (accessed on 21 June 2020).
- Dickson, A.G.; Sabine, C.L.; Christian, J.R.; Bargeron, C.P. North Pacific Marine Science Organization. In Guide to Best Practices for Ocean CO2 Measurements; Dickson, A.G., Sabine, C.L., Christian, J.R., Bargeron, C.P., North Pacific Marine Science Organization, Eds.; North Pacific Marine Science Organization: Sidney, BC, Australia, 2007; ISBN 978-1-897176-07-8. [Google Scholar]
- Higgins, J.J. Introduction to Modern Nonparametric Statistics; Duxbury Advanced Series: Pacific Grove, CA, USA, 2004; ISBN 0-534-38775-6. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Method 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Greenacre, M. Data reporting and visualization in ecology. Polar Biol. 2016, 39, 2189–2205. [Google Scholar] [CrossRef] [Green Version]
- Kvernvik, A.C.; Hoppe, C.J.M.; Greenacre, M.; Verbiest, S.; Wiktor, J.; Gabrielsen, T.M.; Reigstad, M.; Leu, E. Arctic sea ice algae differ markedly from phytoplankton in their ecophysiological characteristics. Mar. Ecol. Progress Ser. in press.
- Leu, E.; Graeve, M.; Wulff, A. A (too) bright future? Arctic diatoms under radiation stress. Polar Biol. 2016, 39, 1711–1724. [Google Scholar] [CrossRef] [Green Version]
- Kvernvik, A.C.; Rokitta, S.D.; Leu, E.; Harms, L.; Gabrielsen, T.M.; Rost, B.; Hoppe, C.J.M. Higher sensitivity towards light stress and ocean acidification in an Arctic sea-ice-associated diatom compared to a pelagic diatom. New Phytol. 2020, 226, 1708–1724. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, J.P.; Church, D.F.; Pryor, W.A. The kinetics of the autoxidation of polyunsaturated fatty acids. Lipids 1987, 22, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Girotti, A.W. Photosensitized oxidation of membrane lipids: Reaction pathways, cytotoxic effects, and cytoprotective mechanisms. J. Photochem. Photobiol. B Biol. 2001, 63, 103–113. [Google Scholar] [CrossRef]
- Mayzaud, P.; Claustre, H.; Augier, P. Effect of variable nutrient supply on fatty acid composition of phytoplankton grown in an enclosed experimental ecosystem. Mar. Ecol. Prog. Ser. 1990, 60, 123–140. [Google Scholar] [CrossRef]
- Hayakawa, K.; Handa, N.; Kawanobe, K.; Wong, C.S. Factors controlling the temporal variation of fatty acids in particulate matter during a phytoplankton bloom in a marine mesocosm. Mar. Chem. 1996, 52, 233–244. [Google Scholar] [CrossRef]
- Skerratt, J.H.; Nichols, P.D.; McMeekin, T.A.; Burton, H. Seasonal and inter-annual changes in planktonic biomass and community structure in eastern Antarctica using signature lipids. Mar. Chem. 1995, 51, 93–113. [Google Scholar] [CrossRef]
- Mayzaud, P.; Chanut, J.P.; Ackman, R.G. Seasonal changes of the biochemical composition of marine particulate matter with special reference to fatty acids and sterols. Mar. Ecol. Prog. Ser. 1989, 56, 189–204. [Google Scholar] [CrossRef]
- Wraige, J.E.; Belt, T.S.; Lewis, A.C.; Cooke, A.D.; Robert, J.-M.; Massé, G.; Rowland, S.J. Variations in structures and distributions of C25 highly branched isoprenoid (HBI) alkenes in cultures of the diatom, Haslea ostrearia (Simonsen). Organ. Geochem. 1997, 27, 497–505. [Google Scholar] [CrossRef]
- Wraige, E.J.; Belt, S.T.; Massé, G.; Robert, J.-M.; Rowland, S.J. Variations in distributions of C25 highly branched isoprenoid (HBI) alkenes in the diatom, Haslea ostrearia: Influence of salinity. Organ. Geochem. 1998, 28, 855–859. [Google Scholar] [CrossRef]
- Wraige, E.J.; Johns, L.; Belt, S.T.; Massé, G.; Robert, J.-M.; Rowland, S. Highly branched C25 isoprenoids in axenic cultures ofHaslea ostrearia. Phytochemistry 1999, 51, 69–73. [Google Scholar] [CrossRef]
- Dieckmann, G.; Hellmer, H. The Importance of Sea Ice: An Overview. Available online: https://epic.awi.de/id/eprint/21259/ (accessed on 17 June 2020).
- Gradinger, R.R.; Kaufman, M.R.; Bluhm, B.A. Pivotal role of sea ice sediments in the seasonal development of near-shore Arctic fast ice biota. Mar. Ecol. Progress Ser. 2009, 394, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Rau, G.H.; Takahashi, T.; Des Marais, D.J.; Repeta, D.J.; Martin, J.H. The relationship between δ13C of organic matter and (CO2 (aq)) in ocean surface water: Data from a JGOFS site in the northeast Atlantic Ocean and a model. Geochim. Cosmochim. Acta 1992, 56, 1413–1419. [Google Scholar] [CrossRef]
- Søreide, J.E.; Carroll, M.L.; Hop, H.; Ambrose, W.G., Jr.; Hegseth, E.N.; Falk-Petersen, S. Sympagic-pelagic-benthic coupling in Arctic and Atlantic waters around Svalbard revealed by stable isotopic and fatty acid tracers. Mar. Biol. Res. 2013, 9, 831–850. [Google Scholar] [CrossRef]
- Tremblay, J.E.; Michel, C.; Hobson, K.A.; Gosselin, M.; Price, N.M. Bloom dynamics in early opening waters of the Arctic Ocean. Limnol. Oceanogr. 2006, 51, 900–912. [Google Scholar] [CrossRef] [Green Version]
- Gradinger, R. Sea-ice algae: Major contributors to primary production and algal biomass in the Chukchi and Beaufort Seas during May/June 2002. Deep Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 1201–1212. [Google Scholar] [CrossRef]
- Fransson, A.; Chierici, M.; Miller, L.A.; Carnat, G.; Shadwick, E.; Thomas, H.; Pineault, S.; Papakyriakou, T.N. Impact of sea-ice processes on the carbonate system and ocean acidification at the ice-water interface of the Amundsen Gulf, Arctic Ocean. J. Geophys. Res. Oceans 2013, 118, 7001–7023. [Google Scholar] [CrossRef]
- Wang, S.W.; Budge, S.M.; Iken, K.; Gradinger, R.R.; Springer, A.M.; Wooller, M.J. Importance of sympagic production to Bering Sea zooplankton as revealed from fatty acid-carbon stable isotope analyses. Mar. Ecol. Progress Ser. 2015, 518, 31–50. [Google Scholar] [CrossRef]
- Burian, A.; Nielsen, J.M.; Hansen, T.; Bermudez, R.; Winder, M. The potential of fatty acid isotopes to trace trophic transfer in aquatic food-webs. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190652. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station. | Date | Snow | IP25 | Σ-Diatom FA | Σ-Diatom FA: POC | C (POC) | Chl a | Salinity | T ice | PO4 | SiO2 | NO3 | DIC | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (pg mL−1) | (ng mL−1) | (ng/µg mL−1) | (µg L−1) | (µg L−1) | (°C) | µmol/L | µmol/L | µmol/L | (µmol/kg) | ||||||||||||||||||||||||
| Transect | Mean | ± | SD | Mean | ± | SD | Mean | ± | SD | Mean | ± | SD | Mean | ± | SD | Mean | ± | SD | Mean | ± | SD | Mean | ± | SD | Mean | ± | SD | Mean | ± | SD | |||
| IS | 28.4.17 | NA | 50.5 | ± | 11.0 | 178.0 | ± | 82.0 | 12.7 | ± | 4.9 | 13,671.4 | ± | 2781.8 | 190.4 | ± | 19.8 | 8.7 | ± | 2.5 | −1.9 | ± | 0.0 | 1.1 | 1.0 | 1.5 | ± | 0.9 | 16.2 | ± | 2.6 | ||
| IM | 28.4.17 | NA | 40.5 | ± | 7.1 | 60.1 | ± | 20.0 | 11.5 | ± | 1.2 | 5216.7 | ± | 1411.4 | 119.5 | ± | 12.8 | 5.3 | ± | 0.8 | −1.6 | ± | 0.0 | 1.1 | 0.2 | 1.6 | ± | 0.4 | 5.2 | ± | 0.5 | ||
| MS | 29.4.17 | NA | 31.3 | ± | 10.5 | 25.1 | ± | 1.3 | 5.4 | ± | 0.3 | 4674.0 | ± | 41.7 | 167.9 | ± | 17.6 | 7.2 | ± | 1.3 | −1.7 | ± | 0.0 | 1.3 | 0.6 | 0.9 | ± | 0.3 | 4.2 | ± | 0.7 | 344 | |
| VMF1 | 30.4.17 | NA | 2.9 | ± | 0.6 | 116.1 | ± | 13.0 | 12.0 | ± | 0.6 | 9742.7 | ± | 1314.5 | 181.5 | ± | 7.9 | 12.7 | ± | 0.2 | −1.7 | ± | 0.1 | 1.5 | 0.8 | 0.3 | ± | 0.3 | 0.6 | ± | 0.2 | 331 | |
| VMF2 | 30.4.17 | NA | 0.5 | ± | 0.3 | 97.4 | ± | 41.7 | 38.8 | ± | 2.1 | 2542.8 | ± | 1174.2 | 44.5 | ± | 17.8 | 7.9 | ± | 0.0 | −1.5 | ± | 0.0 | 0.3 | 0.0 | 0.2 | ± | 0.1 | 0.2 | ± | 0.0 | 319 | |
| Time series | |||||||||||||||||||||||||||||||||
| MS | 7.4.17 | NA | 6.2 | ± | 5.7 | 13.1 | ± | 8.4 | 5.3 | ± | 1.0 | 2330.6 | ± | 1039.3 | 68.8 | ± | 9.8 | 12.2 | ± | 0.7 | −1.6 | ± | 0.0 | 0.4 | 0.1 | 0.3 | ± | 0.1 | 3.9 | ± | 0.7 | 387 | |
| MS | 23.4.17 | High snow | 10.9 | ± | 3.7 | 32.1 | ± | 2.4 | 5.4 | ± | 0.6 | 5923.3 | ± | 333.2 | 259.2 | ± | 20.1 | 14.0 | ± | 0.8 | −2.0 | ± | 0.1 | 0.8 | 0.1 | 0.5 | ± | 0.2 | 14.2 | ± | 2.8 | ||
| MS | 23.4.17 | Low snow | 22.1 | ± | 2.7 | 141.1 | ± | 21.1 | 16.2 | ± | 2.6 | 8938.2 | ± | 2377.1 | 252.3 | ± | 63.5 | 14.9 | ± | 0.7 | −1.9 | ± | 0.0 | 1.4 | 0.3 | 0.7 | ± | 0.2 | 3.1 | ± | 1.8 | 369 | |
| MS | 29.4.17 | NA | 31.3 | ± | 10.5 | 25.1 | ± | 1.3 | 5.4 | ± | 0.3 | 4674.0 | ± | 41.7 | 167.9 | ± | 17.6 | 7.2 | ± | 1.3 | −1.7 | ± | 0.0 | 1.3 | 0.6 | 0.9 | ± | 0.3 | 4.2 | ± | 0.7 | 344 | |
| MS | 2.5.17 | No snow | 10.0 | ± | 1.0 | 477.2 | ± | 24.7 | 49.4 | ± | 4.1 | 9666.8 | ± | 298.3 | 105.8 | ± | 8.3 | 12.2 | ± | 0.3 | −1.8 | ± | 0.1 | 1.5 | 0.2 | 0.5 | ± | 0.2 | 0.2 | ± | 0.0 | 337 | |
| MS | 2.5.17 | High snow | 9.0 | ± | 2.7 | 99.6 | ± | 42.8 | 12.7 | ± | 2.3 | 7632.1 | ± | 1906.9 | 161.4 | ± | 6.8 | 12.3 | ± | 0.1 | −1.7 | ± | 0.1 | 1.3 | ± | 0.3 | 0.3 | ± | 0.1 | 0.7 | ± | 0.2 | |
| Biomarkers | NO3 (μmol L−1) | SiO2 (μmol L−1) | C: N (molar) |
|---|---|---|---|
| IP25/POC | 0.770 * | 0.802 ** | −0.782 * |
| 16:1(n-7) | −0.794 ** | −0.261 | 0.903 ** |
| 16:4(n-1) | 0.758 * | 0.462 | −0.939 ** |
| 20:5(n-3) | 0.709 | 0.669 | −0.782 * |
| δ13C POC | −0.673 | −0.438 | 0.891 ** |
| δ13C 16:1(n-7) | −0.733 * | −0.498 | 0.818 ** |
| δ13C 16:4(n-1) | 0.006 | −0.486 | −0.188 |
| δ13C 20:5(n-3) | −0.370 | −0.608 | 0.418 |
| Unambiguously Specific for Sea Ice Algae vs. Phytoplankton | Produced in Dominating Species | Reflects Species Composition | Changes with Growth Conditions | Distinction between Sympagic and Pelagic POM Based on | |
|---|---|---|---|---|---|
| HBIs | Yes | No | +/− HBI producer | ? (probably yes) | Specific compounds, exclusively produced by some species belonging to only one of the groups |
| FATM | No | Yes | yes (group level) | Relative composition changes | Not possible—unless major taxonomic differences between both groups |
| δ13C POC | Sometimes | Yes | No | Yes | Higher δ13C (average from bulk POC) in sympagic communities due to high biomass densities and slow replenishment of DIC pool in spatially restricted environments only found under limited conditions |
| δ13C FATM | Sometimes | Yes | No | Yes | Higher δ13C (in a single compound) in sympagic communities due to high biomass densities and slow replenishment of DIC pool in spatially restricted environment—only found under limited conditions |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leu, E.; Brown, T.A.; Graeve, M.; Wiktor, J.; Hoppe, C.J.M.; Chierici, M.; Fransson, A.; Verbiest, S.; Kvernvik, A.C.; Greenacre, M.J. Spatial and Temporal Variability of Ice Algal Trophic Markers—With Recommendations about Their Application. J. Mar. Sci. Eng. 2020, 8, 676. https://doi.org/10.3390/jmse8090676
Leu E, Brown TA, Graeve M, Wiktor J, Hoppe CJM, Chierici M, Fransson A, Verbiest S, Kvernvik AC, Greenacre MJ. Spatial and Temporal Variability of Ice Algal Trophic Markers—With Recommendations about Their Application. Journal of Marine Science and Engineering. 2020; 8(9):676. https://doi.org/10.3390/jmse8090676
Chicago/Turabian StyleLeu, Eva, Thomas A. Brown, Martin Graeve, Jozef Wiktor, Clara J. M. Hoppe, Melissa Chierici, Agneta Fransson, Sander Verbiest, Ane C. Kvernvik, and Michael J. Greenacre. 2020. "Spatial and Temporal Variability of Ice Algal Trophic Markers—With Recommendations about Their Application" Journal of Marine Science and Engineering 8, no. 9: 676. https://doi.org/10.3390/jmse8090676
APA StyleLeu, E., Brown, T. A., Graeve, M., Wiktor, J., Hoppe, C. J. M., Chierici, M., Fransson, A., Verbiest, S., Kvernvik, A. C., & Greenacre, M. J. (2020). Spatial and Temporal Variability of Ice Algal Trophic Markers—With Recommendations about Their Application. Journal of Marine Science and Engineering, 8(9), 676. https://doi.org/10.3390/jmse8090676