Key Environmental Factors Controlling Planktonic Foraminiferal and Pteropod Community’s Response to Late Quaternary Hydroclimate Changes in the South Aegean Sea (Eastern Mediterranean)

 ,
,  , ,
, , 
Abstract
:1. Introduction
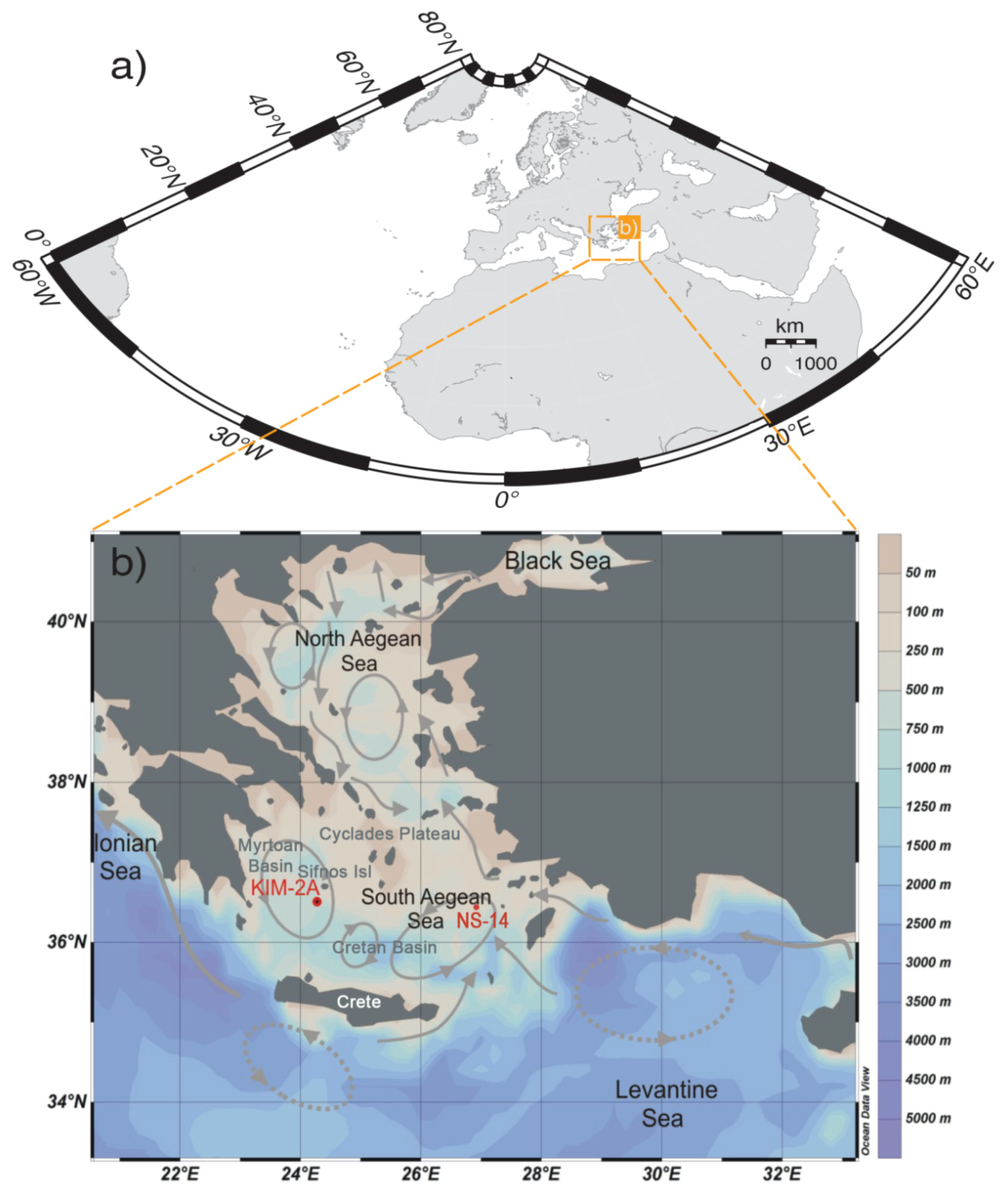
2. Regional and Climatic Setting
3. Materials and Methods
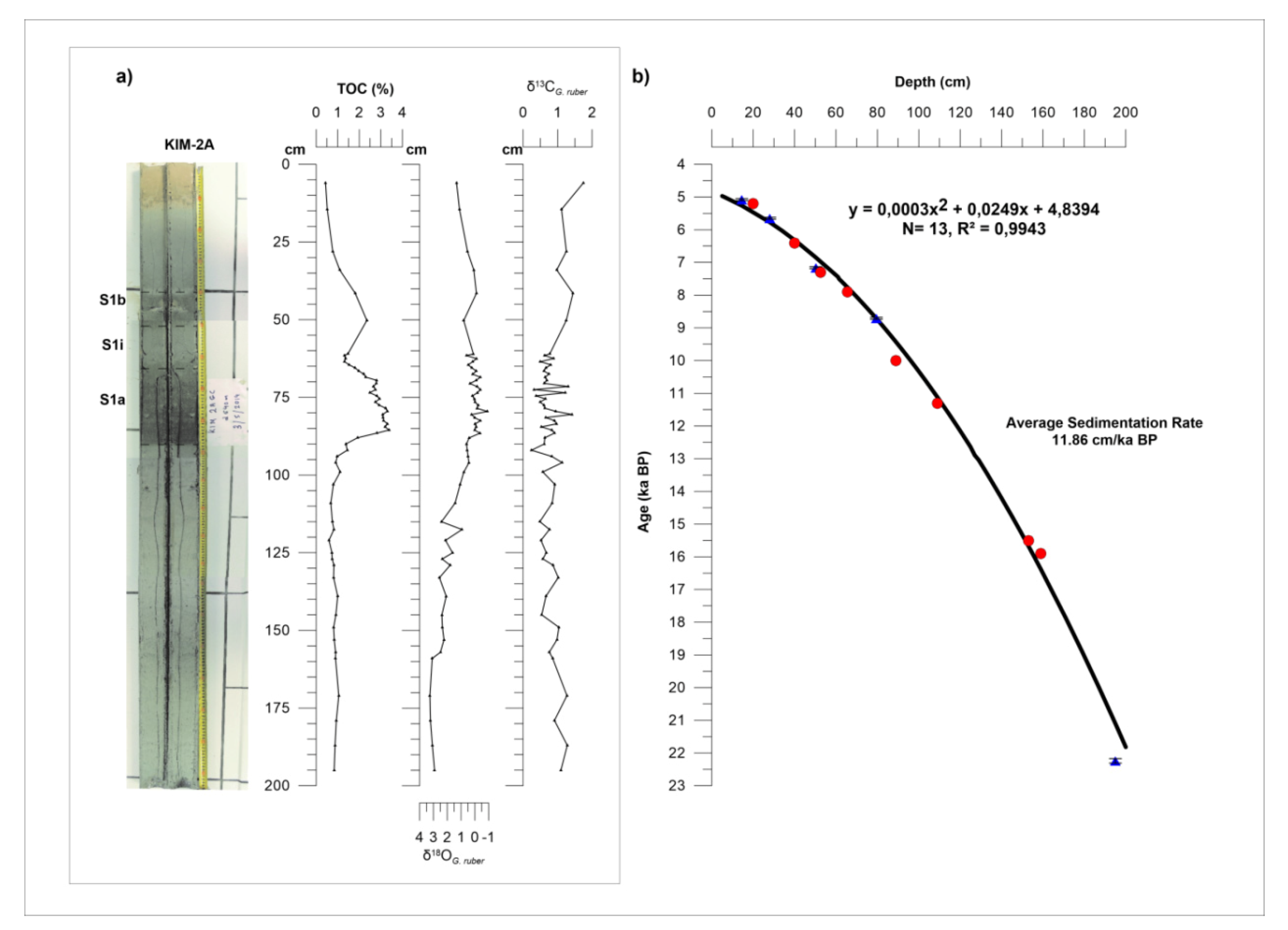
3.1. Location and Sampling Strategy of Core KIM-2A
3.2. Micropaleontological Analyses
3.3. Total Organic Carbon and Stable Isotopes
3.4. Chronology
3.5. Multivariate Statistical Analyses
3.6. Paleoceanographic Indices
4. Results
4.1. Lithological Description, Time Stratigraphic Framework, and Sedimentation Rates
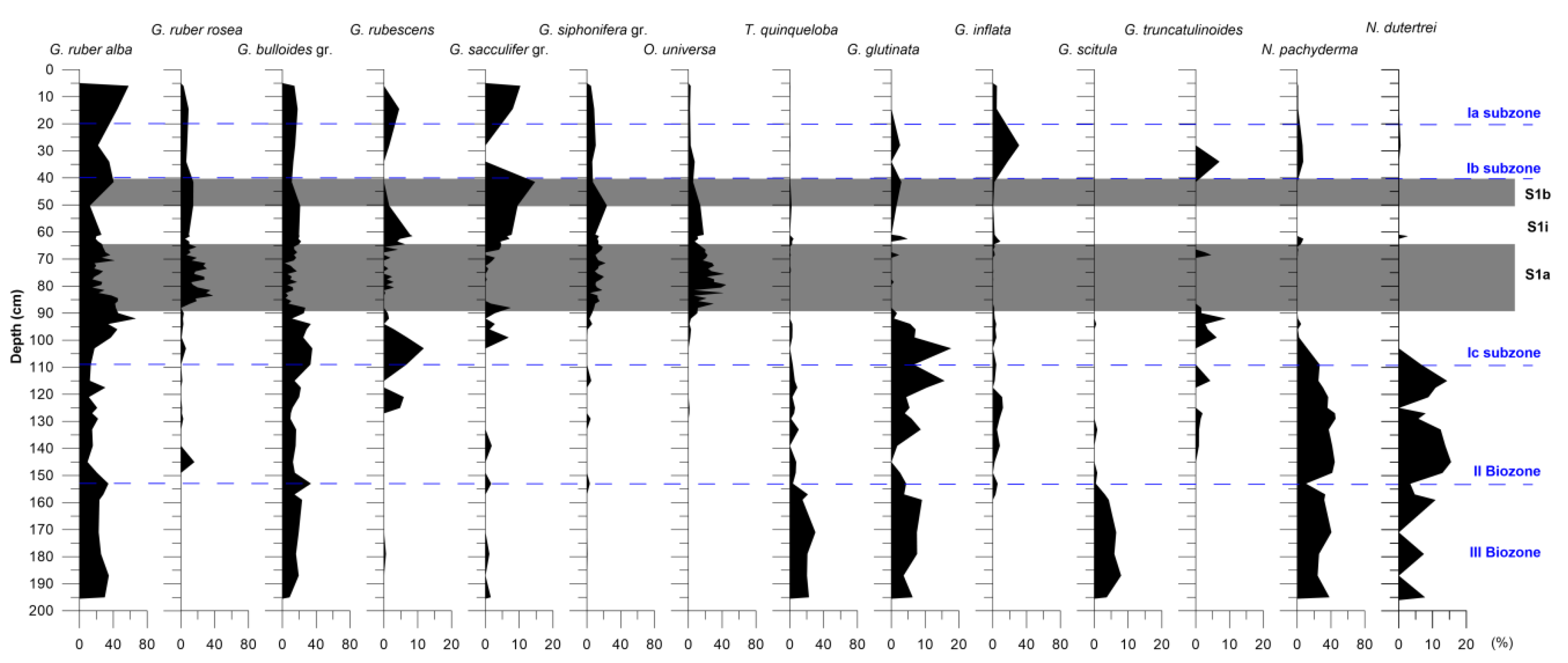
4.2. Planktonic Foraminifera Distribution Pattern
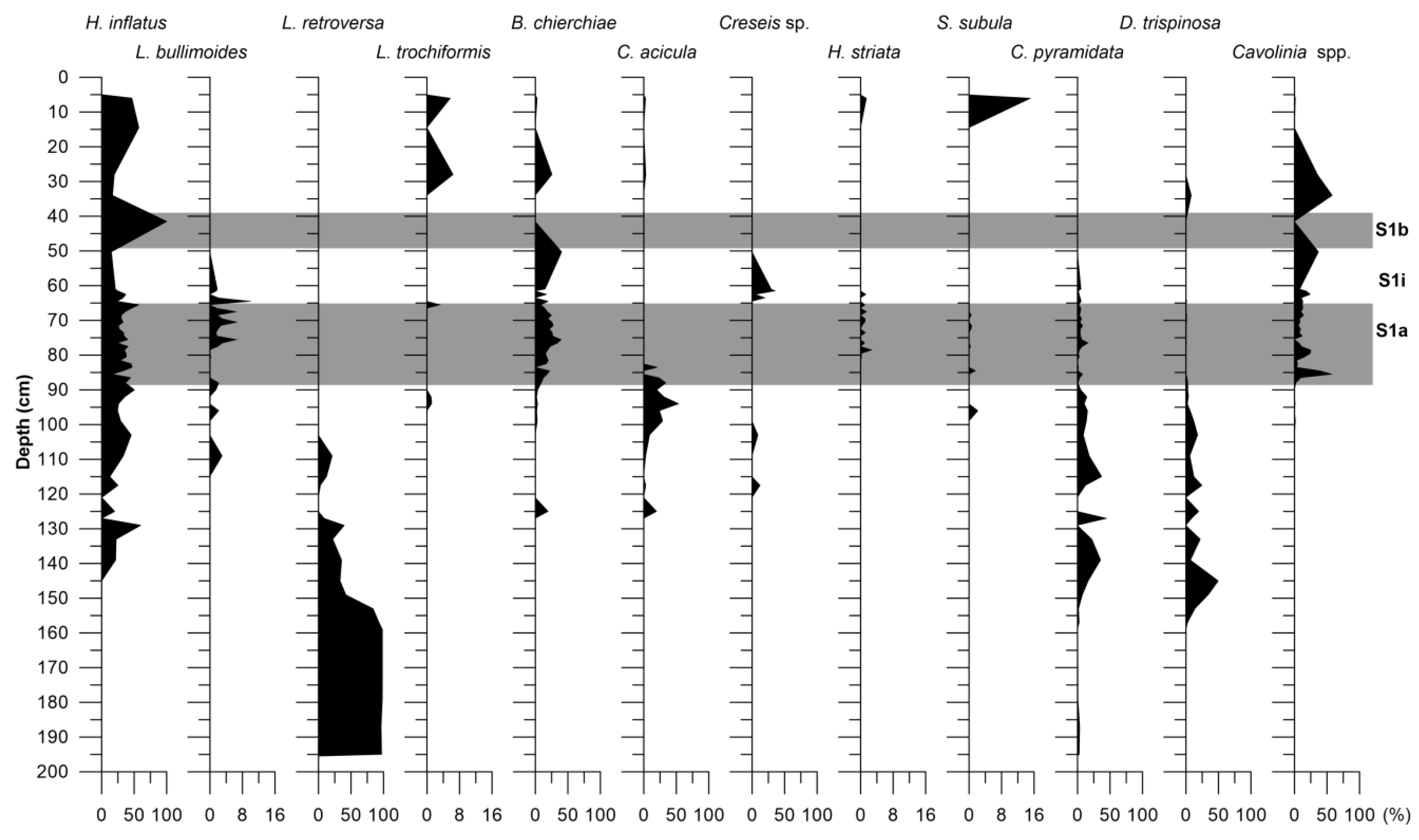
4.3. Pteropod Distribution Pattern
4.4. Total Organic Carbon and Stable Isotopes
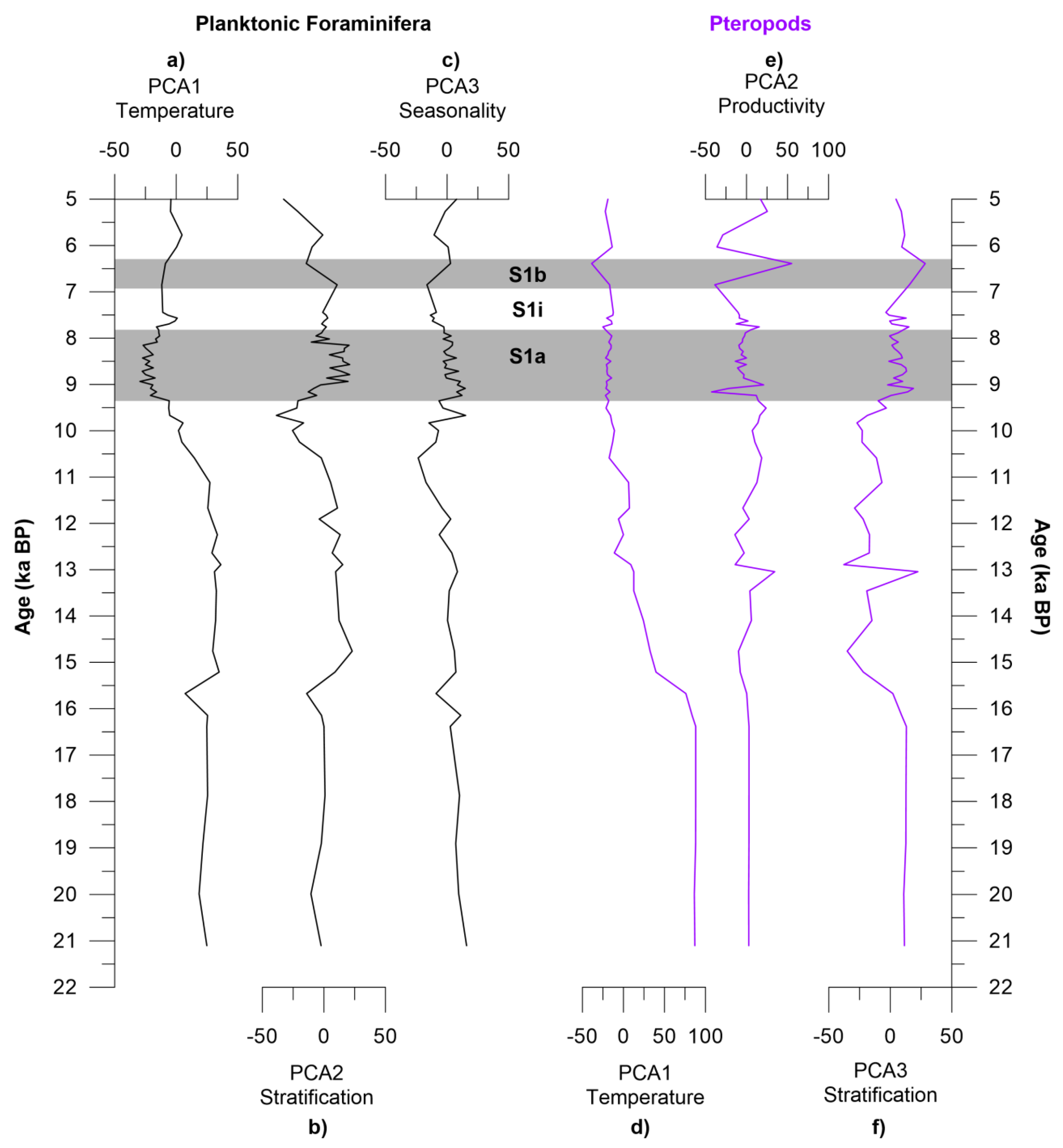
4.5. Principal Component Analysis
4.5.1. Planktonic Foraminifera
4.5.2. Pteropods
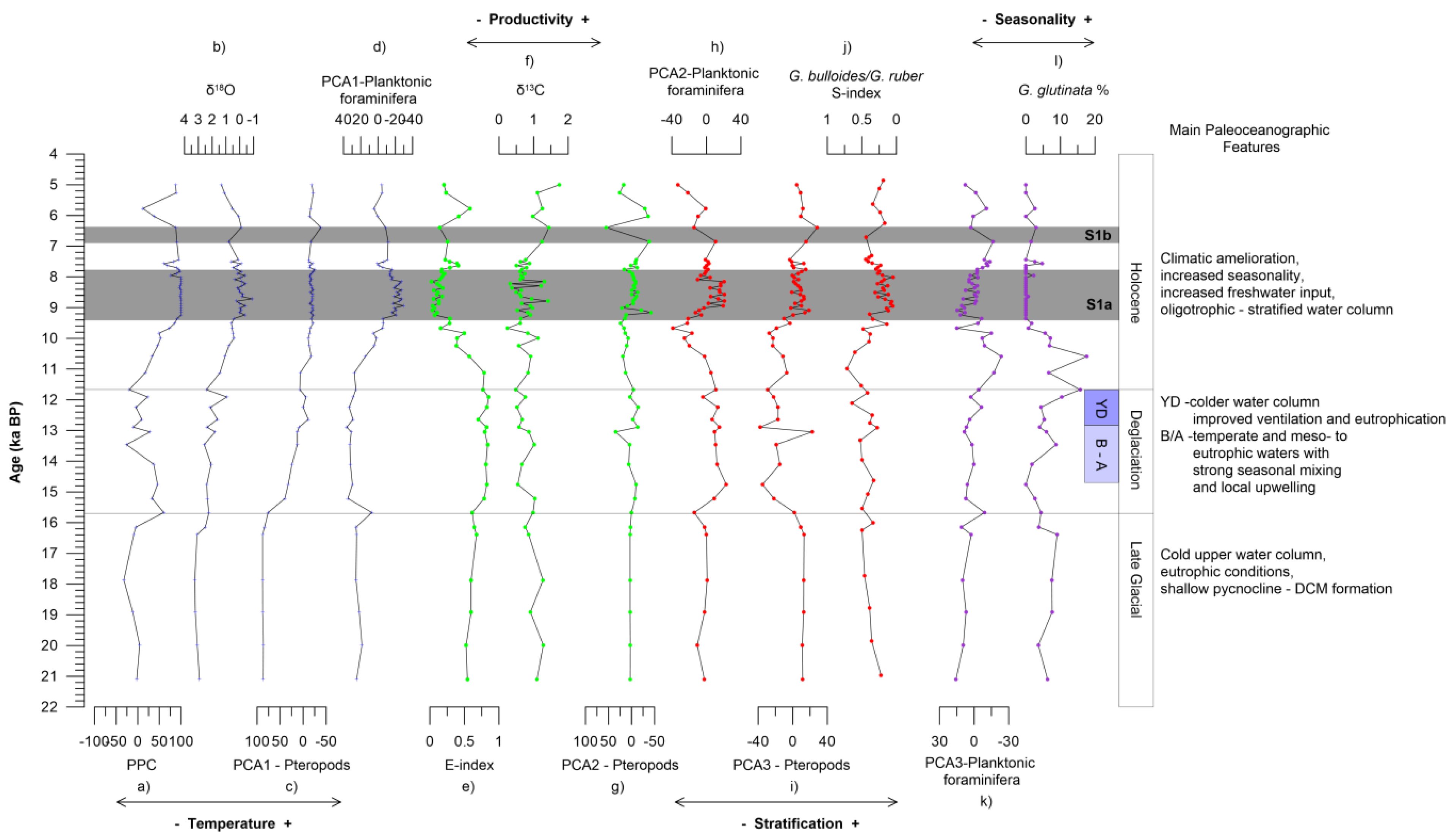
5. Discussion
5.1. Factors Controlling Planktonic Fauna Distribution in the Aegean Sea
5.2. Paleoceanographic Reconstruction
5.2.1. Late Glacial
5.2.2. Deglaciation
5.2.3. Holocene
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zervakis, G.I.; Moncalvo, J.-M.; Vilgalys, R. Molecular phylogeny, biogeography and speciation of the mushroom species Pleurotus cystidiosus and allied taxa. Microbiology 2004, 150, 715–726. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Rohling, E.J.; Grant, K.; Bolshaw, M.; Roberts, A.; Siddall, M.; Hemleben, C.; Kucera, M. Antarctic temperature and global sea level closely coupled over the past five glacial cycles. Nat. Geosci. 2009. [Google Scholar] [CrossRef]
- Aksu, A.E.; Yaşar, D.; Mudie, P.J. Paleoclimatic and paleoceanographic conditions leading to development of sapropel layer S1 in the Aegean Sea. Palaeogeogr. Palaeoclim. Palaeoecol. 1995, 116, 71–101. [Google Scholar] [CrossRef]
- Roussakis, G.; Karageorgis, A.P.; Conispoliatis, N.; Lykousis, V. Last glacial–Holocene sediment sequences in N. Aegean basins: Structure, accumulation rates and clay mineral distribution. Geo-Mar. Lett. 2004, 24, 97–111. [Google Scholar] [CrossRef]
- Geraga, M.; Ioakim, C.; Lykousis, V.; Tsaila-Monopolis, S.; Mylona, G. The high-resolution palaeoclimatic and palaeoceanographic history of the last 24,000 years in the central Aegean Sea, Greece. Palaeogeogr. Palaeoclim. Palaeoecol. 2010, 287, 101–115. [Google Scholar] [CrossRef]
- Kontakiotis, G. Late Quaternary paleoenvironmental reconstruction and paleoclimatic implications of the Aegean Sea (eastern Mediterranean) based on paleoceanographic indexes and stable isotopes. Quat. Int. 2016, 401, 28–42. [Google Scholar] [CrossRef] [Green Version]
- Lykousis, V.; Chronis, G.; Tselepides, A.; Price, N.B.; Theocharis, A.; Siokou-Frangou, I.; Van Wambeke, F.; Danovaro, R.; Stavrakakis, S.; Duineveld, G.; et al. Major outputs of the recent multidisciplinary biogeochemical researches undertaken in the Aegean Sea. J. Mar. Syst. 2002, 33–34, 313–334. [Google Scholar] [CrossRef]
- Poulos, S.E. The Mediterranean and Black Sea Marine System: An overview of its physico-geographic and oceanographic characteristics. Earth Sci. Rev. 2020, 200, 103004. [Google Scholar] [CrossRef]
- Kontakiotis, G. Palaeoceanographic and Palaeoclimatic Study of Eastern Mediterranean During Late Quaternary, Based on Planktonic Foraminiferal Assemblages (in Greek, with English extended abstract). Ph.D. Thesis, National and Kapodistrian University of Athens, Athens, Greece, 2012. [Google Scholar]
- Poulos, S.E.; Drakopoulos, P.G.; Collins, M.B. Seasonal variability in sea surface oceanographic conditions in the Aegean Sea (Eastern Mediterranean): An overview. J. Mar. Syst. 1997, 13, 225–244. [Google Scholar] [CrossRef]
- Geraga, M.; Tsaila-Monopolis, S.; Ioakim, C.; Papatheodorou, G.; Ferentinos, G. Short-term climate changes in the southern Aegean Sea over the last 48,000 years. Palaeogeogr. Palaeoclim. Palaeoecol. 2005, 220, 311–332. [Google Scholar] [CrossRef]
- Kuhnt, T.; Schmiedl, G.; Ehrmann, W.; Hamann, Y.; Hemleben, C. Deep-sea ecosystem variability of the Aegean Sea during the past 22 kyr as revealed by Benthic Foraminifera. Mar. Micropaleontol. 2007, 64, 141–162. [Google Scholar] [CrossRef]
- Kotthoff, U.; Müller, U.C.; Pross, J.; Schmiedl, G.; Lawson, I.T.; van de Schootbrugge, B.; Schulz, H. Lateglacial and Holocene vegetation dynamics in the Aegean region: An integrated view based on pollen data from marine and terrestrial archives. Holocene 2008, 18, 1019–1032. [Google Scholar] [CrossRef]
- Giamali, C.; Koskeridou, E.; Antonarakou, A.; Ioakim, C.; Kontakiotis, G.; Karageorgis, A.P.; Roussakis, G.; Karakitsios, V. Multiproxy ecosystem response of abrupt Holocene climatic changes in the northeastern Mediterranean sedimentary archive and hydrologic regime. Quat. Res. 2019, 92, 665–685. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Ziveri, P.; Gogou, A.; Marino, G.; Lykousis, V.; Bouloubassi, I.; Emeis, K.-C.; Kouli, K.; Dimiza, M.; Rosell-Melé, A.; et al. Late Glacial-Holocene climate variability at the south-eastern margin of the Aegean Sea. Mar. Geol. 2009, 266, 182–197. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Antonarakou, A.; Kouli, K.; Dimiza, M.; Kontakiotis, G.; Papanikolaou, M.D.; Ziveri, P.; Mortyn, P.G.; Lianou, V.; Lykousis, V.; et al. Late Glacial–Holocene ecostratigraphy of the south-eastern Aegean Sea, based on plankton and pollen assemblages. Geo Mar. Lett. 2009, 29, 249–267. [Google Scholar] [CrossRef]
- Drinia, H.; Antonarakou, A.; Tsourou, T.; Kontakiotis, G.; Psychogiou, M.; Anastasakis, G. Foraminifera eco-biostratigraphy of the southern Evoikos outer shelf, central Aegean Sea, during MIS 5 to present. Cont. Shelf Res. 2016, 126. [Google Scholar] [CrossRef]
- Koutrouli, A.; Anastasakis, G.; Kontakiotis, G.; Ballengee, S.; Kuehn, S.; Pe-Piper, G.; Piper, D.J.W. The early to mid-Holocene marine tephrostratigraphic record in the Nisyros-Yali-Kos volcanic center, SE Aegean Sea. J. Volcanol. Geotherm. Res. 2018, 366, 96–111. [Google Scholar] [CrossRef]
- Antonarakou, A.; Kontakiotis, G.; Zarkogiannis, S.; Mortyn, P.G.; Drinia, H.; Koskeridou, E.; Anastasakis, G. Planktonic foraminiferal abnormalities in coastal and open marine eastern Mediterranean environments: A natural stress monitoring approach in recent and early Holocene marine systems. J. Mar. Syst. 2018, 181, 63–78. [Google Scholar] [CrossRef]
- Louvari, M.A.; Drinia, H.; Kontakiotis, G.; Di Bella, L.; Antonarakou, A.; Anastasakis, G. Impact of latest-glacial to Holocene sea-level oscillations on central Aegean shelf ecosystems: A benthic foraminiferal palaeoenvironmental assessment of South Evoikos Gulf, Greece. J. Mar. Syst. 2019, 199, 103181. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Mortyn, P.G.; Antonarakou, A.; Martínez-Botí, M.A.; Triantaphyllou, M.V. Field-based validation of a diagenetic effect on G. ruber Mg/Ca paleothermometry: Core top results from the Aegean Sea (eastern Mediterranean). Geochem. Geophys. Geosyst. 2011, 12. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Karakitsios, V.; Mortyn, P.G.; Antonarakou, A.; Drinia, H.; Anastasakis, G.; Agiadi, K.; Kafousia, N.; De Rafelis, M. New insights into the early Pliocene hydrographic dynamics and their relationship to the climatic evolution of the Mediterranean Sea. Palaeogeogr. Palaeoclim. Palaeoecol. 2016, 459, 348–364. [Google Scholar] [CrossRef]
- Antonarakou, A.; Kontakiotis, G.; Mortyn, P.G.; Drinia, H.; Sprovieri, M.; Besiou, E.; Tripsanas, E. Biotic and geochemical (δ18O, δ13C, Mg/Ca, Ba/Ca) responses of Globigerinoides ruber morphotypes to upper water column variations during the last deglaciation, Gulf of Mexico. Geochim. Cosmochim. Acta 2015, 170, 69–93. [Google Scholar] [CrossRef]
- Le Houedec, S.; Mojtahid, M.; Bicchi, E.; de Lange, G.J.; Hennekam, R. Suborbital Hydrological Variability Inferred From Coupled Benthic and Planktic Foraminiferal-Based Proxies in the Southeastern Mediterranean During the Last 19 ka. Paleoceanogr. Paleoclimatol. 2020, 35, e2019PA003827. [Google Scholar] [CrossRef]
- Cacho, I.; Grimalt, J.O.; Canals, M.; Sbaffi, L.; Shackleton, N.J.; Schönfeld, J.; Zahn, R. Variability of the western Mediterranean Sea surface temperature during the last 25,000 years and its connection with the Northern Hemisphere climatic changes. Paleoceanography 2001, 16, 40–52. [Google Scholar] [CrossRef]
- Filippidi, A.; Triantaphyllou, M.; de Lange, G. Eastern-Mediterranean ventilation variability during sapropel S1 formation, evaluated at two sites influenced by deep-water formation from Adriatic and Aegean Seas. Quat. Sci. Rev. 2016, 144, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Kontakiotis, G.; Besiou, E.; Antonarakou, A.; Zarkogiannis, S.D.; Kostis, A.; Mortyn, P.G.; Moissette, P.; Cornée, J.J.; Schulbert, C.; Drinia, H.; et al. Decoding sea surface and paleoclimate conditions in the eastern Mediterranean over the Tortonian-Messinian Transition. Palaeogeogr. Palaeoclim. Palaeoecol. 2019, 534, 109312. [Google Scholar] [CrossRef]
- Vasiliev, I.; Karakitsios, V.; Bouloubassi, I.; Agiadi, K.; Kontakiotis, G.; Antonarakou, A.; Triantaphyllou, M.; Gogou, A.; Kafousia, N.; de Rafélis, M.; et al. Large Sea Surface Temperature, Salinity, and Productivity-Preservation Changes Preceding the Onset of the Messinian Salinity Crisis in the Eastern Mediterranean Sea. Paleoceanogr. Paleoclimatol. 2019, 34, 182–202. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Mortyn, G.P.; Antonarakou, A.; Drinia, H. Assessing the reliability of foraminiferal Mg/Ca thermometry by comparing field-samples and culture experiments: A review. Geol. Q. 2016, 60, 547–560. [Google Scholar] [CrossRef] [Green Version]
- Kontakiotis, G.; Karakitsios, V.; Cornée, J.-J.; Moissette, P.; Zarkogiannis, S.D.; Pasadakis, N.; Koskeridou, E.; Manoutsoglou, E.; Drinia, H.; Antonarakou, A. Preliminary results based on geochemical sedimentary constraints on the hydrocarbon potential and depositional environment of a Messinian sub-salt mixed siliciclastic-carbonate succession onshore Crete (Plouti section, eastern Mediterranean). Mediterr. Geosci. Rev. 2020. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Antonarakou, A.; Mortyn, P.G.; Drinia, H.; Anastasakis, G.; Zarkogiannis, S.; Möbius, J. Morphological recognition of Globigerinoides ruber morphotypes and their susceptibility to diagenetic alteration in the eastern Mediterranean Sea. J. Mar. Syst. 2017, 174, 12–24. [Google Scholar] [CrossRef]
- Wall-Palmer, D.; Smart, C.W.; Hart, M.B.; Leng, M.J.; Borghini, M.; Manini, E.; Aliani, S.; Conversi, A. Late Pleistocene pteropods, heteropods and planktonic foraminifera from the Caribbean Sea, Mediterranean Sea and Indian Ocean. Micropaleontology 2014, 60, 557–578. [Google Scholar]
- Buccheri, G.; Capretto, G.; Di Donato, V.; Esposito, P.; Ferruzza, G.; Pescatore, T.; Russo Ermolli, E.; Senatore, M.R.; Sprovieri, M.; Bertoldo, M.; et al. A high resolution record of the last deglaciation in the southern Tyrrhenian Sea: Environmental and climatic evolution. Mar. Geol. 2002, 186, 447–470. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Antonarakou, A.; Zachariasse, W.J. Late Quaternary palaeoenvironmental changes in the Aegean Sea: Interrelations and interactions between north and south Aegean Sea. Bull. Geol. Soc. Greece 2013, 47, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Zarkogiannis, S.; Kontakiotis, G.; Antonarakou, A. Recent planktonic foraminifera population and size response to Eastern Mediterranean hudrography. Rev. Micropaleontol. 2020, 69, 100450. [Google Scholar] [CrossRef]
- Casford, J.S.L.; Abu-Zied, R.; Rohling, E.J.; Cooke, S.; Fontanier, C.; Leng, M.; Millard, A.; Thomson, J. A stratigraphically controlled multiproxy chronostratigraphy for the eastern Mediterranean. Paleoceanography 2007, 22. [Google Scholar] [CrossRef]
- Comeau, S.; Jeffree, R.; Teyssié, J.-L.; Gattuso, J.-P. Response of the Arctic Pteropod Limacina helicina to Projected Future Environmental Conditions. PLoS ONE 2010, 5, e11362. [Google Scholar] [CrossRef]
- Lirer, F.; Sprovieri, M.; Vallefuoco, M.; Ferraro, L.; Pelosi, N.; Giordano, L.; Capotondi, L. Planktonic foraminifera as bio-indicators for monitoring the climatic changes that have occurred over the past 2000 years in the southeastern Tyrrhenian Sea. Integr. Zool. 2014, 9, 542–554. [Google Scholar] [CrossRef]
- Margaritelli, G.; Cisneros, M.; Cacho, I.; Capotondi, L.; Vallefuoco, M.; Rettori, R.; Lirer, F. Climatic variability over the last 3000 years in the Central—Western Mediterranean Sea (Menorca Basin) detected by planktonic foraminifera and stable isotope records. Glob. Planet. Chang. 2018, 169, 179–187. [Google Scholar] [CrossRef]
- Margaritelli, G.; Cacho, I.; Català, A.; Barra, M.; Bellucci, L.G.; Lubritto, C.; Rettori, R.; Lirer, F. Persistent warm Mediterranean surface waters during the Roman period. Sci. Rep. 2020, 10, 10431. [Google Scholar] [CrossRef]
- Tsiolakis, E.; Tsaila-Monopoli, S.; Kontakiotis, G.; Antonarakou, A.; Sprovieri, M.; Geraga, M.; Ferentinos, G.; Zissimos, A. Integrated paleohydrology reconstruction and Pliocene climate variability in Cyprus Island (eastern Mediterranean). IOP Conf. Ser. Earth Environ. Sci. 2019, 362, 012103. [Google Scholar] [CrossRef]
- Zarkogiannis, S.; Kontakiotis, G.; Antonarakou, A.; Mortyn, P.; Drinia, H. Latitudinal Variation of Planktonic Foraminifera Shell Masses During Termination I. IOP Conf. Ser. Earth Environ. Sci. 2019, 221, 012052. [Google Scholar] [CrossRef]
- Checa, H.; Margaritelli, G.; Pena, L.D.; Frigola, J.; Cacho, I.; Rettori, R.; Lirer, F. High resolution paleo-environmental changes during the Sapropel 1 in the North Ionian Sea, central Mediterranean. Holocene 2020. [Google Scholar] [CrossRef]
- Antonarakou, A.; Kontakiotis, G.; Karageorgis, A.P.; Besiou, E.; Zarkogiannis, S.; Drinia, H.; Mortyn, G.P.; Tripsanas, E. Eco-biostratigraphic advances on late Quaternary geochronology and palaeoclimate: The marginal Gulf of Mexico analogue. Geol. Q. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Pujol, C.; Grazzini, C. Distribution patterns of live planktic foraminifers as related to regional hydrology and productive systems of the Mediterranean Sea. Mar. Micropaleontol. 1995, 25, 187–217. [Google Scholar] [CrossRef]
- Zarkogiannis, S.D.; Antonarakou, A.; Tripati, A.; Kontakiotis, G.; Mortyn, P.G.; Drinia, H.; Greaves, M. Influence of surface ocean density on planktonic foraminifera calcification. Sci. Rep. 2019, 9, 533. [Google Scholar] [CrossRef]
- Bazzicalupo, P.; Maiorano, P.; Girone, A.; Marino, M.; Combourieu-Nebout, N.; Pelosi, N.; Salgueiro, E.; Incarbona, A. Holocene climate variability of the Western Mediterranean: Surface water dynamics inferred from calcareous plankton assemblages. Holocene 2020, 30, 691–708. [Google Scholar] [CrossRef]
- Koskeridou, E.; Giamali, C.; Antonarakou, A.; Kontakiotis, G.; Karakitsios, V. Early Pliocene gastropod assemblages from the eastern Mediterranean (SW Peloponnese, Greece) and their palaeobiogeographic implications. Geobios 2017, 50, 267–277. [Google Scholar] [CrossRef]
- Moissette, P.; Cornée, J.-J.; Antonarakou, A.; Kontakiotis, G.; Drinia, H.; Koskeridou, E.; Tsourou, T.; Agiadi, K.; Karakitsios, V. Palaeoenvironmental changes at the Tortonian/Messinian boundary: A deep-sea sedimentary record of the eastern Mediterranean Sea. Palaeogeogr. Palaeoclim. Palaeoecol. 2018, 505, 217–233. [Google Scholar] [CrossRef]
- Karakitsios, V.; Roveri, M.; Lugli, S.; Manzi, V.; Gennari, R.; Antonarakou, A.; Triantaphyllou, M.; Agiadi, K.; Kontakiotis, G.; Kafousia, N.; et al. A record of the Messinian salinity crisis in the eastern Ionian tectonically active domain (Greece, eastern Mediterranean). Basin Res. 2017, 29, 203–233. [Google Scholar] [CrossRef]
- Casford, J.S.L.; Rohling, E.J.; Abu-Zied, R.; Cooke, S.; Fontanier, C.; Leng, M.; Lykousis, V. Circulation changes and nutrient concentrations in the late Quaternary Aegean Sea: A nonsteady state concept for sapropel formation. Paleoceanography 2002, 17, 14-1–14-11. [Google Scholar] [CrossRef] [Green Version]
- Casford, J.S.L.; Rohling, E.J.; Abu-Zied, R.H.; Fontanier, C.; Jorissen, F.J.; Leng, M.J.; Schmiedl, G.; Thomson, J. A dynamic concept for eastern Mediterranean circulation and oxygenation during sapropel formation. Palaeogeogr. Palaeoclim. Palaeoecol. 2003, 190, 103–119. [Google Scholar] [CrossRef]
- Aksu, A.E.; Hiscott, R.; Isler, E. Late Quaternary chronostratigraphy of the Aegean Sea sediments: Special reference to the ages of sapropels S1–S5. Turk. J. Earth Sci. 2016, 25, 1–18. [Google Scholar] [CrossRef]
- Howes, E.L.; Eagle, R.A.; Gattuso, J.-P.; Bijma, J. Comparison of Mediterranean Pteropod Shell Biometrics and Ultrastructure from Historical (1910 and 1921) and Present Day (2012) Samples Provides Baseline for Monitoring Effects of Global Change. PLoS ONE 2017, 12, e0167891. [Google Scholar] [CrossRef] [PubMed]
- Lalli, C.M.; Gilmer, R.W. Pelagic Snails: The Biology of Holoplanktonic Gastropod Mollusks; Stanford University Press: Stanford, CA, USA, 1989. [Google Scholar]
- Hunt, B.; Pakhomov, E.; Hosie, G.W.; Siegel, V.; Ward, P.; Bernard, K. Pteropods in Southern Ocean ecosystems. Prog. Oceanogr. 2008, 78, 193–221. [Google Scholar] [CrossRef]
- Lochte, K.; Pfannkuche, O. Processes driven by the small sized organisms at the water-sediment interface. In Ocean Margin Systems; Wefer, G., Billet, D., Hebbeln, D., Jørgensen, B.B., Schluter, M., Weering, T.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Vinogradov, M. Food sources for the deep water fauna. Speed of decomposition of dead Pteropoda. Dokl. Akad. Nauk SSSR 1961, 138, 1439–1442. [Google Scholar]
- Herman, Y. Vertical and horizontal distribution of pteropods in Quaternary sequences. In The Micropalaeontology of Oceans; Funnell, B.M., Reidel, W.R., Eds.; Cambridge University Press: Cambridge, UK, 1971; pp. 463–486. [Google Scholar]
- Buccheri, G. Pteropods as climatic indicators in Quaternary sequences: A Lower-Middle Pleistocene sequence outcropping in Cava Puleo (Ficarazzi, Palermo, Sicilia). Palaeogeogr. Palaeoclim. Palaeoecol. 1984, 45, 75–86. [Google Scholar] [CrossRef]
- Almogi-Labin, A.; Hemleben, C.; Meischner, D.; Erlenkeuser, H. Paleoenvironmental events during the last 13,000 years in the central Red Sea as recorded by pteropoda. Paleoceanography 1991, 6, 83–98. [Google Scholar] [CrossRef]
- Almogi-Labin, A.; Hemleben, C.; Meischner, D. Carbonate preservation and climatic changes in the central Red Sea during the last 380 kyr as recorded by pteropods. Mar. Micropaleontol. 1998, 33, 87–107. [Google Scholar] [CrossRef]
- Almogi-Labin, A.; Bar-Matthews, M.; Shriki, D.; Kolosovsky, E.; Paterne, M.; Schilman, B.; Ayalon, A.; Aizenshtat, Z.; Matthews, A. Climatic variability during the last ∼90 ka of the southern and northern Levantine Basin as evident from marine records and speleothems. Quat. Sci. Rev. 2009, 28, 2882–2896. [Google Scholar] [CrossRef]
- Singh, A.D.; Nisha, N.R.; Joydas, T.V. Distribution patterns of Recent pteropods in surface sediments of the western continental shelf of India. J. Micropalaeontol. 2005, 24, 39. [Google Scholar] [CrossRef] [Green Version]
- Almogi-Labin, A.; Edelman-Furstenberg, Y.; Hemleben, C. Variations in the biodiversity of thecosomatous pteropods during the Late Quaternary as a response to environmental changes in the Gulf of Aden—Red Sea—Gulf of Aqaba ecosystem. In Aqaba–Eliat, the Improbable Gulf—Environment, Biodiversity and Preservation; Por, F.D., Ed.; The Hebrew University Magnes Press: Jerusalem, Israel, 2008; pp. 31–48. [Google Scholar]
- Johnson, R.; Manno, C.; Ziveri, P. Spring distribution of shelled pteropods across the Mediterranean Sea. Biogeosci. Discuss. 2020, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Howes, E.L.; Stemmann, L.; Assailly, C.; Irisson, J.O.; Dima, M.; Bijma, J.; Gattuso, J.P. Pteropod time series from the North Western Mediterranean (1967–2003): Impacts of pH and climate variability. Mar. Ecol. Prog. Ser. 2015, 531, 193–206. [Google Scholar] [CrossRef] [Green Version]
- Lykousis, V. Subaqueous bedforms on the Cyclades Plateau (NE Mediterranean)—Evidence of Cretan Deep Water Formation? Cont. Shelf Res. 2001, 21, 495–507. [Google Scholar] [CrossRef]
- Karageorgis, A.P.; Ioakim, C.; Rousakis, G.; Sakellariou, D.; Vougioukalakis, G.; Panagiotopoulos, I.P.; Zimianitis, E.; Koutsopoulou, E.; Kanellopoulos, T.; Papatrechas, C. Geomorphology, sedimentology and geochemistry in the marine area between Sifnos and Kimolos Islands, Greece. Bull. Geol. Soc. Greece 2016, 50, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Piper, D.; Perissoratis, C. Quaternary neotectonics of the South Aegean arc. Mar. Geol. 2003, 198, 259–288. [Google Scholar] [CrossRef]
- Antonarakou, A.; Kontakiotis, G.; Vasilatos, C.; Besiou, E.; Zarkogiannis, S.; Drinia, H.; Mortyn, P.; Tsaparas, N.; Makri, P.; Karakitsios, V. Evaluating the Effect of Marine Diagenesis on Late Miocene Pre-Evaporitic Sedimentary Successions of Eastern Mediterranean Sea. IOP Conf. Ser. Earth Environ. Sci. 2019, 221, 012051. [Google Scholar] [CrossRef]
- Hemleben, C.; Spindler, M.; Anderson, O. Modern Planktic Foraminifera; Springer-Verlag: New York, NY, USA, 1989; Volume 22. [Google Scholar]
- Rohling, E.J.; Jorissen, F.; Grazzini, C.V.; Zachariasse, W.J. Northern Levantine and Adriatic Quaternary planktic foraminifera; Reconstruction of paleoenvironmental gradients. Mar. Micropaleontol. 1993, 21, 191–218. [Google Scholar] [CrossRef]
- Aurahs, R.; Grimm, G.; Hemleben, V.; Hemleben, C.; Kucera, M. Geographical distribution of cryptic genetic types in the planktonic foraminifer Globigerinoides ruber. Mol. Ecol. 2009, 18, 1692–1706. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff Method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Angelova, V.; Akova, V.; Ivanov, K.; Licheva, P.A. Comparative study of titrimetric methods for determination of organic carbon in soils, compost and sludge. J. Int. Sci. Publ. Ecol. Saf. 2014, 8, 430–440. [Google Scholar] [CrossRef]
- Wang, B.; Wu, R.; Fu, X. Pacific–East Asian Teleconnection: How Does ENSO Affect East Asian Climate? J. Clim. 2000, 13, 1517–1536. [Google Scholar] [CrossRef]
- Löwemark, L.; Hong, W.-L.; Yui, T.-F.; Hugn, G.-W. A test of different factors influencing the isotopic signal of planktonic foraminifera in surface sediments from the northern South China Sea. Mar. Micropaleontol. 2005, 55, 49–62. [Google Scholar] [CrossRef]
- Spero, H.J.; Mielke, K.M.; Kalve, E.M.; Lea, D.W.; Pak, D.K. Multispecies approach to reconstructing eastern equatorial Pacific thermocline hydrography during the past 360 kyr. Paleoceanography 2003, 18. [Google Scholar] [CrossRef]
- Stuiver, M.; Reimer, P.J. Extended 14C data base and revised CALIB 3.0 14C age calibration program. Radiocarbon 1993, 35, 215–320. [Google Scholar] [CrossRef] [Green Version]
- Facorellis, Y.; Maniatis, Y. Apparent 14C ages of marine mollusk shells from a Greek Island: Calculation of the marine reservoir effect in the Aegean Sea. Radiocarbon 1998, 40, 963–973. [Google Scholar] [CrossRef] [Green Version]
- Reimer, P.; Bard, E.; Bayliss, A.; Beck, J.; Blackwell, P.; Ramsey, C.; Buck, C.; Cheng, H.; Edwards, R.; Friedrich, M.; et al. IntCal13 and MARINE13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 2013, 55, 1869–1887. [Google Scholar] [CrossRef] [Green Version]
- Jorissen, F.J.; Asioli, A.; Borsetti, A.M.; Capotondi, L.; de Visser, J.P.; Hilgen, F.J.; Rohling, E.J.; van der Borg, K.; Vergnaud Grazzini, C.; Zachariasse, W.J. Late Quaternary central Mediterranean biochronology. Mar. Micropaleontol. 1993, 21, 169–189. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 21 June 2001).
- Sbaffi, L.; Wezel, F.C.; Curzi, G.; Zoppi, U. Millennial- to centennial-scale palaeoclimatic variations during Termination I and the Holocene in the central Mediterranean Sea. Global Planet. Change 2004, 40, 201. [Google Scholar] [CrossRef]
- Anastasakis, G.C.; Stanley, D.J. Sapropels and organic-rich variants in the Mediterranean: Sequence development and classification. Geol. Soc. Spec. Publ. 1984, 15, 497. [Google Scholar] [CrossRef]
- Mercone, D.; Thomson, J.; Croudace, I.W.; Siani, G.; Paterne, M.; Troelstra, S. Duration of S1, the most recent sapropel in the eastern Mediterranean Sea, as indicated by accelerator mass spectrometry radiocarbon and geochemical evidence. Paleoceanography 2000, 15, 336–347. [Google Scholar] [CrossRef]
- Zachariasse, W.; Jorissen, F.; Perissoratis, C.; Rohling, E.; Tsapralis, V. Late Quaternary foraminiferal changes and the nature of sapropel S1 in Skopelos Basin. In Proceedings of the 5th Hellenic Symposium of Oceanography and Fisheries, Kavala, Greece, 15–18 April 1997; pp. 391–394. [Google Scholar]
- Capotondi, L.; Maria Borsetti, A.; Morigi, C. Foraminiferal ecozones, a high resolution proxy for the late Quaternary biochronology in the central Mediterranean Sea. Mar. Geol. 1999, 153, 253–274. [Google Scholar] [CrossRef]
- Hayes, A.; Rohling, E.J.; De Rijk, S.; Kroon, D.; Zachariasse, W.J. Mediterranean planktonic foraminiferal faunas during the last glacial cycle. Mar. Geol. 1999, 153, 239–252. [Google Scholar] [CrossRef]
- Casford, J.; Rohling, E.J.; Abu-Zied, R.; Cooke, S.; Boessenkool, K.P.; Brinkhuis, H.; Vries, C.; Wefer, G.; Geraga, M.; Papatheodorou, G.; et al. Mediterranean climate variability during the Holocene. Mediterr. Mar. Sci. 2001, 2, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Schneider, A.; Wallace, W.R.D.; Kortzinger, A. Alkalinity of the Mediterranean Sea. Geophys. Res. Lett. 2007, 34. [Google Scholar] [CrossRef] [Green Version]
- Wilke, I.; Meggers, H.; Bickert, T. Depth habitats and seasonal distributions of recent planktic foraminifers in the Canary Islands region (29 °N) based on oxygen isotopes. Deep Sea Res. Part I Oceanogr. Res. Pap. 2009, 56, 89. [Google Scholar] [CrossRef]
- Wit, J.C.; Reichart, G.-J.; Jung, S.J.A.; Kroon, D. Approaches to unravel seasonality in sea surface temperatures using paired single-specimen foraminiferal δ18O and Mg/Ca analyses. Paleoceanography 2010, 25. [Google Scholar] [CrossRef] [Green Version]
- Goudeau, M.L.S. Seasonality variations in the Central Mediterranean during climate change events in the Late Holocene. Palaeogeogr. Palaeoclim. Palaeoecol. 2015, 418, 304–318. [Google Scholar] [CrossRef]
- Kucera, M.; Weinelt, M.; Kiefer, T.; Pflaumann, U.; Hayes, A.; Weinelt, M.; Chen, M.-T.; Mix, A.C.; Barrows, T.T.; Cortijo, E.; et al. Reconstruction of sea-surface temperatures from assemblages of planktonic foraminifera: Multi-technique approach based on geographically constrained calibration data sets and its application to glacial Atlantic and Pacific Oceans. Quat. Sci. Rev. 2005, 24, 951–998. [Google Scholar] [CrossRef]
- Žarić, S.; Donner, B.; Fischer, G.; Mulitza, S.; Wefer, G. Sensitivity of planktic foraminifera to sea surface temperature and export production as derived from sediment trap data. Mar. Micropaleontol. 2005, 55, 75–105. [Google Scholar] [CrossRef]
- Fraile, I.; Schulz, M.; Mulitza, S.; Kucera, M. Predicting the global distribution of planktonic foraminifera using a dynamic ecosystem model. Biogeosciences 2008, 5, 891–911. [Google Scholar] [CrossRef] [Green Version]
- Fraile, I.; Mulitza, S.; Schulz, M. Modeling planktonic foraminiferal seasonality: Implications for sea-surface temperature reconstructions. Mar. Micropaleontol. 2009, 72, 1–9. [Google Scholar] [CrossRef]
- Sprovieri, R.; Stefano, E.; Incarbona, A.; Gargano, M. A high-resolution record of the last deglaciation in the Sicily Channel based on foraminifera and calcareous nannofossil quantitative distribution. Palaeogeogr. Palaeoclim. Palaeoecol. 2003, 202, 119–142. [Google Scholar] [CrossRef]
- Di Donato, V.; Esposito, P.; Russo-Ermolli, E.; Scarano, A.; Cheddadi, R. Coupled atmospheric and marine palaeoclimatic reconstruction for the last 35 ka in the Sele Plain—Gulf of Salerno area (southern Italy). Quat. Int. 2008, 190, 146–157. [Google Scholar] [CrossRef]
- Biekart, W.J. Euthecosomatous pteropods as paleohydrological and paleoecological indicators in a Tyrrhenian deep-sea core. Palaeogeogr. Palaeoclim. Palaeoecol. 1989, 71, 205–224. [Google Scholar] [CrossRef]
- Kutzbach, J.E.; Guetter, P.J. The Influence of Changing Orbital Parameters and Surface Boundary Conditions on Climate Simulations for the Past 18,000 Years. J. Atmos. Sci. 1986, 43, 1726–1759. [Google Scholar] [CrossRef] [Green Version]
- Kotthoff, U.; Koutsodendris, A.; Pross, J.; Schmiedl, G.; Bornemann, A.; Kaul, C.; Marino, G.; Peyron, O.; Schiebel, R. Impact of Lateglacial cold events on the northern Aegean region reconstructed from marine and terrestrial proxy data. J. Quat. Sci. 2011, 26, 86–96. [Google Scholar] [CrossRef]
- Dormoy, I.; Peyron, O.; Combourieu Nebout, N.; Goring, S.; Kotthoff, U.; Magny, M.; Pross, J. Terrestrial climate variability and seasonality changes in the Mediterranean region between 15,000 and 4000 years BP deduced from marine pollen records. Clim. Past 2009, 5, 615–632. [Google Scholar] [CrossRef] [Green Version]
- Björck, S.; Walker, M.; Cwynar, L.; Johnsen, S.; Knudsen, K.; Lowe, J.; Wohlfarth, B. An event stratigraphy for the Last Termination in the North Atlantic region based on the Greenland ice-core record: A proposal by the INTIMATE group. J. Quat. Sci. 1998, 13, 283–292. [Google Scholar] [CrossRef]
- Rasmussen, S.O.; Vinther, B.M.; Clausen, H.B.; Andersen, K.K. Early Holocene climate oscillations recorded in three Greenland ice cores. Quat. Sci. Rev. 2007, 26, 1907–1914. [Google Scholar] [CrossRef] [Green Version]
- Peyron, O.; Bégeot, C.; Brewer, S.; Heiri, O.; Magny, M.; Millet, L.; Ruffaldi, P.; Campo, E.; Yu, G. Lateglacial climate in the Jura mountains (France) based on different quantitative reconstruction approaches from pollen, lake-levels, and chironomids. Quat. Res. 2005, 64, 197–211. [Google Scholar] [CrossRef]
- Bordon, A.; Peyron, O.; Lezine, A.-M.; Brewer, S.; Fouache, E. Pollen-inferred Late-Glacial and Holocene climate in southern Balkans (Lake Maliq). Quat. Int. 2009, 200, 19–30. [Google Scholar] [CrossRef]
- Larocque, I.; Finsinger, W. Late-glacial chironomid-based temperature reconstructions for Lago Piccolo di Avigliana in the southwestern Alps (Italy). Palaeogeogr. Palaeoclim. Palaeoecol. 2008, 257, 207–223. [Google Scholar] [CrossRef]
- Sijinkumar, A.V.; Bejugam, N.; Guptha, M.V.S. Late Quaternary record of pteropod preservation from the Andaman Sea. Mar. Geol. 2010, 275. [Google Scholar] [CrossRef] [Green Version]
- Rottman, M.L. Net tow and surface sediment distributions of pteropods in the South China Sea region: Comparison and oceanographic implications. Mar. Micropaleontol. 1980, 5, 71–110. [Google Scholar] [CrossRef]
- Rossignol-Strick, M. Sea-land correlation of pollen records in the Eastern Mediterranean for the glacial-interglacial transition: Biostratigraphy versus radiometric time-scale. Quat. Sci. Rev. 1995, 14, 893. [Google Scholar] [CrossRef]
- Laskar, J.; Robutel, P.; Joutel, F.; Gastineau, M.; Correia, A.C.M.; Levrard, B. A long-term numerical solution for the insolation quantities of the Earth. A&A 2004, 428, 261–285. [Google Scholar]
- Rossignol-Strick, M. African monsoons, an immediate climate response to orbital insolation. Nature 1983, 304, 46–49. [Google Scholar] [CrossRef]
- Rohling, E.J. Glacial conditions in the Red Sea. Paleoceanography 1994, 9, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Weikert, H.G.; Cederbaum, L.S. Particle-number-dependent theory of few-and many-body systems. Few Body Syst. 1987, 2, 33–51. [Google Scholar] [CrossRef]
- Weikert, H. The vertical distribution of zooplankton in relation to habitat zones in the area of the Atlantis II Deep, central Red Sea. Mar. Ecol. Prog. Ser. 1982, 8, 129–143. [Google Scholar] [CrossRef]
- Wormuth, J. Vertical distributions and diel migrations of Euthecosomata in the northwest Sargasso Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 1981, 28, 1493–1515. [Google Scholar] [CrossRef]
- Bé, A.W.H.; Gilmer, R.W. A zoogeographic and taxonomic review of euthecosomatous Pteropoda. Ocean. Micropaleontol. 1977, 1, 733–808. [Google Scholar]
- Sakthivel, M. A preliminary report on the distribution and relative abundance of Euthecosomata with a note on the seasonal variation of Limacina species in the Indian Ocean. Bull. Nat. Inst. Sci. India 1969, 38, 700–717. [Google Scholar]
- Almogi-Labin, A.; Hemleben, C.; Deuser, W.G. Seasonal variation in the flux of euthecosomatous pteropods collected in a deep sediment trap in the Sargasso Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 1988, 35, 441–464. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMS & Chronostratigraphic Control Points | Depth (cm) | Conventional Radiocarbon Age (BP) | Two Sigma Calibrated Age Range (BP) | Mean Calibrated Age (ka BP) | References |
|---|---|---|---|---|---|
| Beta—425634 | 14.5 | 4890+/−30 | 4845–5325 | 5.08 | |
| Ia/Ib boundary | 20 | 5.2 | [84] | ||
| Beta—425635 | 28 | 5320+/−30 | 5444–5855 | 5.65 | |
| S1b top | 40 | 6.4 | [7,16] | ||
| Beta—425636 | 50.25 | 6790+/−30 | 7036–7292 | 7.16 | |
| S1b base | 52.5 | 7.3 | [7,16] | ||
| S1a top | 65.5 | 7.9 | [7,16] | ||
| Beta—425637 | 79.5 | 8320+/−30 | 8532–8883 | 8.71 | |
| S1a base | 89 | 10 | [7,16] | ||
| Ic/II boundary | 109 | 11.3 | [84] | ||
| II/III boundary | 153 | 15.5 | [84] | ||
| δ18OG. ruber depletion | 159 | 15.9 | [52] | ||
| Beta—425638 | 195 | 18890+/−70 | 21962–22508 | 22.24 |
| PCA Factors | Eigenvalue | % Variance | Cumulative % of the Total Variance |
|---|---|---|---|
| 1 | 432.137 | 50.26 | 50.26 |
| 2 | 193.038 | 22.45 | 72.70 |
| 3 | 76.2131 | 8.86 | 81.57 |
| 4 | 53.3399 | 6.20 | 87.77 |
| 5 | 38.4841 | 4.47 | 92.25 |
| 6 | 24.4952 | 2.85 | 95.09 |
| 7 | 15.5611 | 1.81 | 96.90 |
| 8 | 7.97016 | 0.93 | 97.83 |
| 9 | 7.63218 | 0.89 | 98.72 |
| 10 | 5.03693 | 0.58 | 99.30 |
| 11 | 2.88765 | 0.33 | 99.64 |
| 12 | 2.25794 | 0.26 | 99.90 |
| 13 | 0.666282 | 0.08 | 99.98 |
| 14 | 0.159802 | 0.02 | 100.00 |
| PCA | Eigenvalue | % Variance | Cumulative % of the Total Variance |
|---|---|---|---|
| 1 | 1209.14 | 58.66 | 58.66 |
| 2 | 273.91 | 13.29 | 71.95 |
| 3 | 223.817 | 10.86 | 82.81 |
| 4 | 104.846 | 5.09 | 87.89 |
| 5 | 100.727 | 4.89 | 92.78 |
| 6 | 63.5077 | 3.08 | 95.87 |
| 7 | 53.2518 | 2.58 | 98.45 |
| 8 | 23.7844 | 1.15 | 99.60 |
| 9 | 4.32791 | 0.21 | 99.81 |
| 10 | 2.86233 | 0.14 | 99.95 |
| 11 | 0.755953 | 0.04 | 99.99 |
| 12 | 0.236728 | 0.01 | 100.00 |
| Variables | Factor 1 | Factor 2 | Factor 3 |
|---|---|---|---|
| O. universa | −0.511 | 0.363 | 0.194 |
| G. ruber f. alba | −0.158 | −0.776 | 0.443 |
| G. ruber f. rosea | −0.375 | 0.302 | 0.101 |
| G. sacculifer gr. | −0.028 | −0.091 | −0.122 |
| G. siphonifera gr. | −0.251 | 0.099 | −0.152 |
| G. inflata | 0.079 | −0.020 | −0.136 |
| G. bulloides gr. | 0.094 | −0.199 | −0.691 |
| G. rubescens | 0.008 | −0.012 | −0.209 |
| N. pachyderma | 0.643 | 0.320 | 0.305 |
| N. dutertrei | 0.155 | 0.100 | 0.051 |
| T. quinqueloba | 0.197 | 0.001 | 0.258 |
| G. truncatulinoides | 0.007 | −0.060 | 0.004 |
| G. glutinata | 0.129 | −0.014 | −0.110 |
| G. scitula | 0.032 | −0.009 | 0.064 |
| Species | Factor 1 | Factor 2 | Factor 3 |
|---|---|---|---|
| H. inflatus | −0.387 | 0.693 | 0.453 |
| L. bulimoides | −0.015 | −0.012 | 0.010 |
| L. retroversa | 0.895 | 0.175 | 0.304 |
| L. trochiformis | −0.005 | 0.000 | 0.008 |
| B. chierchiae | −0.154 | −0.334 | 0.286 |
| C. acicula | −0.064 | 0.227 | −0.328 |
| Creseis sp. | −0.027 | −0.031 | −0.038 |
| H. striata | −0.004 | −0.002 | 0.008 |
| S. subula | −0.006 | 0.015 | 0.004 |
| C. pyramidata | 0.009 | −0.024 | −0.437 |
| D. trispinosa | 0.051 | −0.020 | −0.420 |
| Cavolinia sp. | −0.131 | −0.570 | 0.380 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giamali, C.; Kontakiotis, G.; Koskeridou, E.; Ioakim, C.; Antonarakou, A. Key Environmental Factors Controlling Planktonic Foraminiferal and Pteropod Community’s Response to Late Quaternary Hydroclimate Changes in the South Aegean Sea (Eastern Mediterranean). J. Mar. Sci. Eng. 2020, 8, 709. https://doi.org/10.3390/jmse8090709
Giamali C, Kontakiotis G, Koskeridou E, Ioakim C, Antonarakou A. Key Environmental Factors Controlling Planktonic Foraminiferal and Pteropod Community’s Response to Late Quaternary Hydroclimate Changes in the South Aegean Sea (Eastern Mediterranean). Journal of Marine Science and Engineering. 2020; 8(9):709. https://doi.org/10.3390/jmse8090709
Chicago/Turabian StyleGiamali, Christina, George Kontakiotis, Efterpi Koskeridou, Chryssanthi Ioakim, and Assimina Antonarakou. 2020. "Key Environmental Factors Controlling Planktonic Foraminiferal and Pteropod Community’s Response to Late Quaternary Hydroclimate Changes in the South Aegean Sea (Eastern Mediterranean)" Journal of Marine Science and Engineering 8, no. 9: 709. https://doi.org/10.3390/jmse8090709
APA StyleGiamali, C., Kontakiotis, G., Koskeridou, E., Ioakim, C., & Antonarakou, A. (2020). Key Environmental Factors Controlling Planktonic Foraminiferal and Pteropod Community’s Response to Late Quaternary Hydroclimate Changes in the South Aegean Sea (Eastern Mediterranean). Journal of Marine Science and Engineering, 8(9), 709. https://doi.org/10.3390/jmse8090709