
Invasive Species Control: Predation on the Alien Crab Percnon gibbesi (H. Milne Edwards, 1853) (Malacostraca: Percnidae) by the Rock Goby, Gobius paganellus Linnaeus, 1758 (Actinopterygii: Gobiidae)




Abstract
:1. Introduction
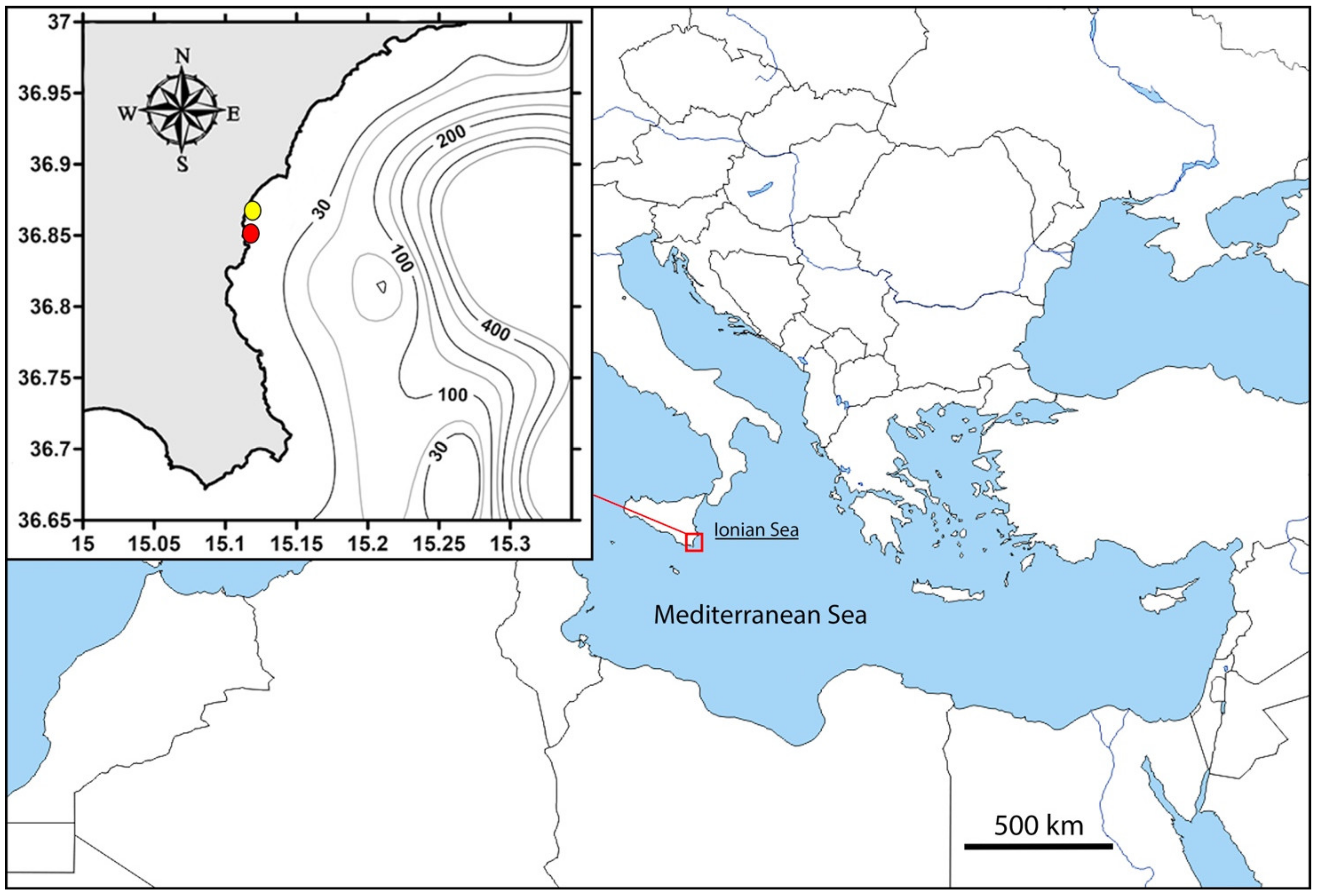
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosholz, E. Ecological and evolutionary consequences of coastal invasions. Trends Ecol. Evol. 2002, 17, 22–27. [Google Scholar] [CrossRef]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef]
- Ricciardi, A.; Palmer, M.E.; Yan, N.D. Should biological invasions be managed as natural disasters? BioScience 2011, 61, 312–317. [Google Scholar] [CrossRef] [Green Version]
- Bailey, S.; Brown, L.; Campbell, M.; Canning-Clode, J.; Carlton, J.; Castro, N.; Chainho, P.; Chan, F.T.; Creed, J.C.; Curd, A.; et al. Trends in the detection of aquatic non-indigenous species across global marine, estuarine and freshwater ecosystems: A 50-year perspective. Divers. Distrib. 2020, 26, 1780–1797. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Noè, S.; Gianguzza, P.; di Trapani, F.; Badalamenti, F.; Vizzini, S.; Fernández, T.V.; Bonaviri, C. Native predators control the population of an invasive crab in no-take marine protected areas. Aquatic Conserv. Mar. Freshw. Ecosyst. 2018, 28, 1229–1237. [Google Scholar] [CrossRef]
- Madenjian, C.P.; Stapanian, M.A.; Witzel, L.D.; Einhouse, D.W.; Pothoven, S.A.; Whitford, H.L. Evidence for predatory control of the invasive round goby. Biol. Invasions 2011, 13, 987–1002. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer’s Catalog of Fishes: Species by Family/Subfamily. California Academy of Sciences, San Francisco, USA. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 17 November 2020).
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley and Sons: Hoboken, NJ, USA, 2016; p. 752. [Google Scholar]
- Kovačić, M. Checklist of gobies (Teleostei: Gobiidae) of the Mediterranean Sea and a key for species identification. Zootaxa 2020, 4877, 75–101. [Google Scholar] [CrossRef]
- Hajji, F.; Ouannes-Ghorbel, A.; Ghorbel, M.; Jarboui, O. Diet shifts of Gobius paganellus (Teleostei, Gobiidae) from the Gulf of Gabes (Central Mediterranean). Vie Milieu 2012, 62, 69–76. [Google Scholar]
- Hajji, F.; Ouannes-Ghorbel, A.; Ghorbel, M.; Jarboui, O. Reproductive biology of the rock goby, Gobius paganellus (Actinopterygii: Perciformes: Gobiidae), on the southern Tunisian coast (Gulf of Gabes). Cienc. Mar. 2012, 38, 505–515. [Google Scholar] [CrossRef] [Green Version]
- Kovačić, M.; la Mesa, A. Feeding ecology of the Buen’s goby, Buenia affinis, in the Kvarner area (Adriatic Sea). Vie Milieu 2008, 58, 277–281. [Google Scholar]
- Tiralongo, F.; Messina, G.; Lombardo, B.M. First data on habitat preference, diet and length-weight relationship of Gobius incognitus Kovačić & Šanda, 2016 (Pisces: Gobiidae). Acta Adriat. 2020, 61, 67–78. [Google Scholar] [CrossRef]
- Iglésias, S.P.; Vukić, J.; Sellos, D.Y.; Soukupová, T.; Šanda, R. Gobius xoriguer, a new offshore Mediterranean goby (Gobiidae), and phylogenetic relationships within the genus Gobius. Ichthyol. Res. 2021, 1–15. [Google Scholar] [CrossRef]
- Kovačić, M.; Šanda, R. A new species of Gobius (Perciformes: Gobiidae) from the Mediterranean Sea and the redescription of Gobius bucchichi. J. Fish Biol. 2016, 88, 1104–1124. [Google Scholar] [CrossRef]
- Kovačić, M.; Patzner, R.; Schliewen, U. A first quantitative assessment of the ecology of cryptobenthic fishes in the Mediterranean Sea. Mar. Biol. 2012, 159, 2731–2742. [Google Scholar] [CrossRef]
- Miller, P.J. Gobiidae. In Fishes of the North-Eastern Atlantic and Mediterranean; Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; pp. 1019–1085. [Google Scholar]
- Tortonese, E. Mediterranean fishes present in the Red Sea: Pan-oceanic and anti-lessepsian species. Cybium 1984, 8, 99–102. [Google Scholar]
- Engin, S.; Seyhan, K. Biological Characteristics of rock goby, Gobius paganellus (Actinopterygii: Perciformes: Gobiidae), in the South-Eastern Black Sea. Acta Ichthyol. Pisc. 2009, 39, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.J. Age, growth, and reproduction of the rock goby, Gobius paganellus L., in the Isle of Man. J. Mar. Biol. Assoc. UK 1961, 41, 737–769. [Google Scholar] [CrossRef]
- Wilkins, H.K.A.; Myers, A.A. Microhabitat utilisation by an assemblage of temperate Gobiidae (Pisces: Teleostei). Mar. Ecol. Prog. Ser. 1992, 90, 103–112. [Google Scholar] [CrossRef]
- Azevedo, J.M.N.; Simas, A.M.V. Age and growth, reproduction and diet of a sublittoral population of the rock goby Gobius paganellus (Teleostei, Gobiidae). Hydrobiologia 2000, 440, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Louiz, I.; Attia, M.B.; Hassine, O.K.B. Some aspects of reproductive biology of Gobius paganellus (Gobiidae) on the north-eastern coasts of Tunisia (Bizerta lagoon). J. Mar. Biol. Assoc. UK 2013, 93, 2235–2246. [Google Scholar] [CrossRef]
- Mazé, R.A. Seasonal and ontogenetic diet shift in an intertidal population of Gobius paganellus (Teleostei, Gobiidae) from the Cantabrian Coast. Vie Milieu 2004, 54, 1–6. [Google Scholar]
- Katsanevakis, S.; Poursanidis, D.; Yokes, M.B.; Mačić, V.; Beqiraj, S.; Kashta, L.; Sghaier, Y.R.; Zakhama-Sraieb, R.; Benamer, I.; Bitar, G.; et al. Twelve years after the first report of the crab Percnon gibbesi (H. Milne Edwards, 1853) in the Mediterranean: Current distribution and invasion rates. J. Biol. Res. 2011, 16, 224–236. [Google Scholar]
- Relini, M.; Orsi, L.; Puccio, V.; Azzurro, E. The exotic crab Percnon gibbesi (H. Milne Edwards, 1853) (Decapoda, Grapsidae) in the Central Mediterranean. Sci. Mar. 2000, 64, 337–340. [Google Scholar] [CrossRef] [Green Version]
- Suaria, G.; Pierucci, A.; Zanello, P.; Fanelli, E.; Chiesa, S.; Azzurro, E. Percnon gibbesi (H. Milne Edwards, 1853) and Callinectes sapidus (Rathbun, 1896) in the Ligurian Sea: Two additional invasive species detection made in collaboration with local fishermen. BioInvasions Rec. 2017, 6, 147–151. [Google Scholar] [CrossRef]
- Pipitone, C.; Badalamenti, F.; Sparrow, A. Contribution to the knowledge of Percnon gibbesi (Decapoda, Grapsidae), an exotic species spreading rapidly in Sicilian waters. Crustaceana 2001, 74, 1009–1017. [Google Scholar]
- Sciberras, M.; Schembri, P.J. Biology and interspecific interactions of the alien crab Percnon gibbesi in the Maltese Islands. Mar. Biol. Res. 2008, 4, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Crocetta, F.; Shokouros-Oskarsson, M.; Doumpas, N.; Giovos, I.; Kalogirou, S.; Langeneck, J.; Tanduo, V.; Tiralongo, F.; Virgili, R.; Kleitou, P. Protect the natives to combat the aliens: Could Octopus vulgaris Cuvier, 1797 be a natural agent for the control of the lionfish invasion in the Mediterranean Sea? J. Mar. Sci. Eng. 2021, 9, 308. [Google Scholar] [CrossRef]
- Kaldonski, N.; Lagrue, C.; Motreuil, S.; Rigaud, T.; Bollache, L. Habitat segregation mediates predation by the benthic fish Cottus gobio on the exotic amphipod species Gammarus roeseli. Naturwissenschaften 2008, 95, 839–844. [Google Scholar] [CrossRef]
- Maljković, A.; van Leeuwen, T.E.; Cove, S.N. Predation on the invasive red lionfish, Pterois volitans (Pisces: Scorpaenidae), by native groupers in the Bahamas. Coral Reefs 2008, 27, 501. [Google Scholar] [CrossRef]
- Louette, G. Use of a native predator for the control of an invasive amphibian. Wildl. Res. 2012, 39, 271–278. [Google Scholar] [CrossRef]
- Giakoumi, S.; Pey, A.; Thiriet, P.; Francour, P.; Guidetti, P. Patterns of predation on native and invasive alien fish in Mediterranean protected areas. Mar. Environ. Res. 2019, 150, 104792. [Google Scholar] [CrossRef]
- Yokes, B.; Galil, B.S. Touchdown—First record of Percnon gibbesi (H. Milne Edwards, 1853) (Crustacea: Decapoda: Grapsidae) from the Levantine coast. Aquat. Invasions 2006, 1, 130–132. [Google Scholar] [CrossRef]
- Galil, B.; Froglia, C.; Noel, P.Y. Atlas of Exotic Species in the Mediterranean. 2008. Available online: www.ciesm.org/atlas/appendix2.html (accessed on 10 December 2020).
- Deudero, S.; Frau, A.; Cerda, M.; Hampel, H. Distribution and densities of the decapod crab Percnon gibbesi, an invasive Grapsidae, in western Mediterranean waters. Mar. Ecol. Prog. Ser. 2005, 285, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Cannicci, S.; Badalamenti, F.; Milazzo, M.; Gomei, M.; Baccarella, A.; Vannini, M. Unveiling the secrets of a successful invader: Preliminary data on the biology and the ecology of the crab Percnon gibbesi (H. Milne Edwards, 1853). Rapp. Com. Int. Mer Médit. 2004, 37, 326. [Google Scholar]
- Jones, R.E.; Petrell, R.J.; Pauly, D. Using modified length-weight relationships to assess the condition of fish. Aquac. Eng. 1999, 20, 261–276. [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor and weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Riedl, R. Fauna e Flora del Mediterraneo; Franco Muzzio Editore: Padua, Italy, 2010; p. 777. [Google Scholar]
- Hayward, P.J.; Ryland, J.S. Handbook of the Marine Fauna of North-West Europe; Oxford University Press: Oxford, UK, 1990; p. 816. [Google Scholar]
- Hyslop, E.J. Stomach content analysis, a review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef] [Green Version]
- Carrassòn, M.; Matallanas, J.; Casadevall, M. Feeding strategies of deep-water morids on the western Mediterranean slope. Deep Sea Res. 1997, 44, 1685–1699. [Google Scholar] [CrossRef]
- N’da, K. Regime alimentaire du rouget de roche Mullus surmuletus (Mullidae) dans le nord du golfe de Gascogne. Cybium 1992, 16, 159–168. [Google Scholar]
- Krebs, J.C. Ecological Methodology; Harper & Row: New York, NY, USA, 1989; p. 620. [Google Scholar]
- Novakowski, G.C.; Hahn, N.S.; Fugi, R. Diet seasonality and food overlap of the fish assemblage in a pantanal pond. Neotrop. Ichthyol. 2008, 6, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.C.; Bizzarro, J.J.; Cailliet, G.M.; Ebert, D.A. Breaking with tradition: Redefining measures for diet description with a case study of the Aleutian skate Bathyraja aleutica (Gilbert 1869). Environ. Biol. Fish. 2012, 95, 3–20. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2018. Available online: http://www.R-project.org (accessed on 10 December 2020).
- Zander, C.D.; Berg, J. Feeding ecology of littoral gobiid and blennioid fishes of the Banyuls area (Mediterranean Sea) II. Prey selection and size preference. Vie Milieu 1984, 34, 149–157. [Google Scholar]
- Cubedo-Verdiell, D.; Oliva-Paterna, F.J.; Torralva, M. Length-weight relationships for 22 fish species of the Mar Menor coastal lagoon (western Mediterranean Sea). J. Appl. Ichthyol. 2006, 22, 293–294. [Google Scholar] [CrossRef]
- Giakoumi, S.; Katsanevakis, S.; Albano, P.G.; Azzurro, E.; Cardoso, A.C.; Cebrian, E.; Deidun, A.; Edelist, D.; Francour, P.; Jimenez, C.; et al. Management priorities for marine invasive species. Sci. Total Environ. 2019, 688, 976–982. [Google Scholar] [CrossRef]
- Byun, C.; Lee, E.J. Ecological application of biotic resistance to control the invasion of an invasive plant, Ageratina altissima. Ecol. Evol. 2017, 7, 2181–2192. [Google Scholar] [CrossRef] [PubMed]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Funk, J.L.; Cleland, E.E.; Suding, K.N.; Zavaleta, E.S. Restoration trough reassembly: Plant traits and invasion resistance. Trends Ecol. Evol. 2008, 23, 695–703. [Google Scholar] [CrossRef]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; The University of Chicago Press: Chicago, IL, USA, 1958. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | N | N Avola | N Noto | N Prey | TL (cm) | SL (cm) | W (g) |
|---|---|---|---|---|---|---|---|
| 4 September 2020 | 15 | 5 | 10 | 11 | 9.52 | 7.89 | 11.45 |
| 14 September 2020 | 11 | 2 | 9 | 7 | 9.24 | 7.68 | 10.52 |
| 21 September 2020 | 12 | 4 | 8 | 9 | 9.54 | 8 | 11.26 |
| 28 September 2020 | 16 | 8 | 8 | 12 | 9.50 | 7.99 | 11.78 |
| 2 October 2020 | 12 | 5 | 7 | 10 | 9.82 | 8.20 | 12.91 |
| 9 October 2020 | 15 | 10 | 5 | 13 | 9.83 | 8.25 | 12.09 |
| 16 October 2020 | 13 | 3 | 10 | 9 | 9.09 | 7.57 | 10.05 |
| 27 October 2020 | 14 | 6 | 8 | 6 | 9.56 | 7.96 | 11.80 |
| 3 November 2020 | 12 | 8 | 4 | 8 | 9.59 | 8.06 | 11.48 |
| 10 November 2020 | 15 | 8 | 7 | 10 | 9.19 | 7.71 | 10.69 |
| 22 November 2020 | 14 | 9 | 5 | 11 | 9.8 | 8.19 | 12.85 |
| 5 December 2020 | 13 | 4 | 9 | 9 | 9.49 | 7.95 | 11.26 |
| Prey types | %F | %N | %W | IRI | %IRI |
|---|---|---|---|---|---|
| Mollusca | 11.31 | 9.36 | 10.24 | 49.916 | 1.62 |
| Aplysia sp. | 0.87 | 0.72 | 1.63 | 2.0435 | 0.07 |
| Bittium lacteum | 0.87 | 0.72 | 0.54 | 1.0956 | 0.04 |
| Patella caerulea | 1.74 | 1.44 | 1.01 | 4.2610 | 0.14 |
| Tritia cuvierii | 1.74 | 1.44 | 1.90 | 5.8116 | 0.19 |
| Gastropoda n.i. | 4.35 | 3.60 | 3.67 | 31.6087 | 1.02 |
| Trochidae n.i. | 1.74 | 1.44 | 1.49 | 5.0956 | 0.16 |
| Polychaeta | 2.61 | 2.16 | 0.95 | 8.113 | 0.26 |
| Polychaetes n.i. | 2.61 | 2.16 | 0.95 | 8.113 | 0.26 |
| Crustacea | 89.59 | 77.73 | 79.65 | 2925.5660 | 94.42 |
| Clibanarius erythropus | 6.96 | 6.47 | 4.27 | 74.7130 | 2.41 |
| Eriphia verrucosa | 4.35 | 3.60 | 3.49 | 30.8415 | 1.00 |
| Pachygrapsus marmoratus | 5.22 | 5.04 | 5.03 | 52.5391 | 1.70 |
| Percnon gibbesi | 33.91 | 30.22 | 40.15 | 2386.4608 | 77.01 |
| Pisidia sp. | 6.09 | 5.04 | 12.42 | 106.2782 | 3.43 |
| Xantho sp. | 6.96 | 5.76 | 4.17 | 69.0783 | 2.23 |
| Amphipoda n.i. | 6.09 | 5.04 | 1.22 | 38.1234 | 1.23 |
| Brachyura n.i. | 6.96 | 5.76 | 4.61 | 69.3753 | 2.24 |
| Caridea n.i. | 6.96 | 5.76 | 1.49 | 50.4347 | 1.63 |
| Isopoda n.i. | 6.09 | 5.04 | 2.80 | 47.7217 | 1.54 |
| Algae | 6.09 | 5.04 | 3.53 | 52.1652 | 1.68 |
| Seagrass (Posidonia oceanica) | 0.87 | 0.72 | 0.43 | 1.0005 | 0.03 |
| Digested | 6.09 | 5.04 | 5.19 | 62.2695 | 2.01 |
| Total Length (TL)-cm | Standard Length (SL)-cm | Weight (W)-g | ||||||
|---|---|---|---|---|---|---|---|---|
| Range | Mean | SD | Range | Mean | SD | Range | Mean | SD |
| 7.1–11.6 | 9.5 | 1.11 | 5.9–9.6 | 7.9 | 0.97 | 4.98–20.14 | 11.53 | 3.80 |
| a | C.I. a | b | C.I. b | r2 | p-value | |||
| 0.014 | 0.011–0.018 | 2.959 | 2.849–3.068 | 0.95 | >0.05 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiralongo, F.; Messina, G.; Lombardo, B.M. Invasive Species Control: Predation on the Alien Crab Percnon gibbesi (H. Milne Edwards, 1853) (Malacostraca: Percnidae) by the Rock Goby, Gobius paganellus Linnaeus, 1758 (Actinopterygii: Gobiidae). J. Mar. Sci. Eng. 2021, 9, 393. https://doi.org/10.3390/jmse9040393
Tiralongo F, Messina G, Lombardo BM. Invasive Species Control: Predation on the Alien Crab Percnon gibbesi (H. Milne Edwards, 1853) (Malacostraca: Percnidae) by the Rock Goby, Gobius paganellus Linnaeus, 1758 (Actinopterygii: Gobiidae). Journal of Marine Science and Engineering. 2021; 9(4):393. https://doi.org/10.3390/jmse9040393
Chicago/Turabian StyleTiralongo, Francesco, Giuseppina Messina, and Bianca Maria Lombardo. 2021. "Invasive Species Control: Predation on the Alien Crab Percnon gibbesi (H. Milne Edwards, 1853) (Malacostraca: Percnidae) by the Rock Goby, Gobius paganellus Linnaeus, 1758 (Actinopterygii: Gobiidae)" Journal of Marine Science and Engineering 9, no. 4: 393. https://doi.org/10.3390/jmse9040393
APA StyleTiralongo, F., Messina, G., & Lombardo, B. M. (2021). Invasive Species Control: Predation on the Alien Crab Percnon gibbesi (H. Milne Edwards, 1853) (Malacostraca: Percnidae) by the Rock Goby, Gobius paganellus Linnaeus, 1758 (Actinopterygii: Gobiidae). Journal of Marine Science and Engineering, 9(4), 393. https://doi.org/10.3390/jmse9040393