
Statistical Optimisation and Kinetic Studies of Molybdenum Reduction Using a Psychrotolerant Marine Bacteria Isolated from Antarctica

 ,
,  ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culture, Maintenance and Media Preparation
2.2. Conventional One-Factor-At-a-Time (OFAT) Optimisation
2.3. Statistical Approach Response Surface Methodology (RSM)
2.4. Kinetic Modelling
3. Results
3.1. Optimisation of Mo Reduction Using OFAT
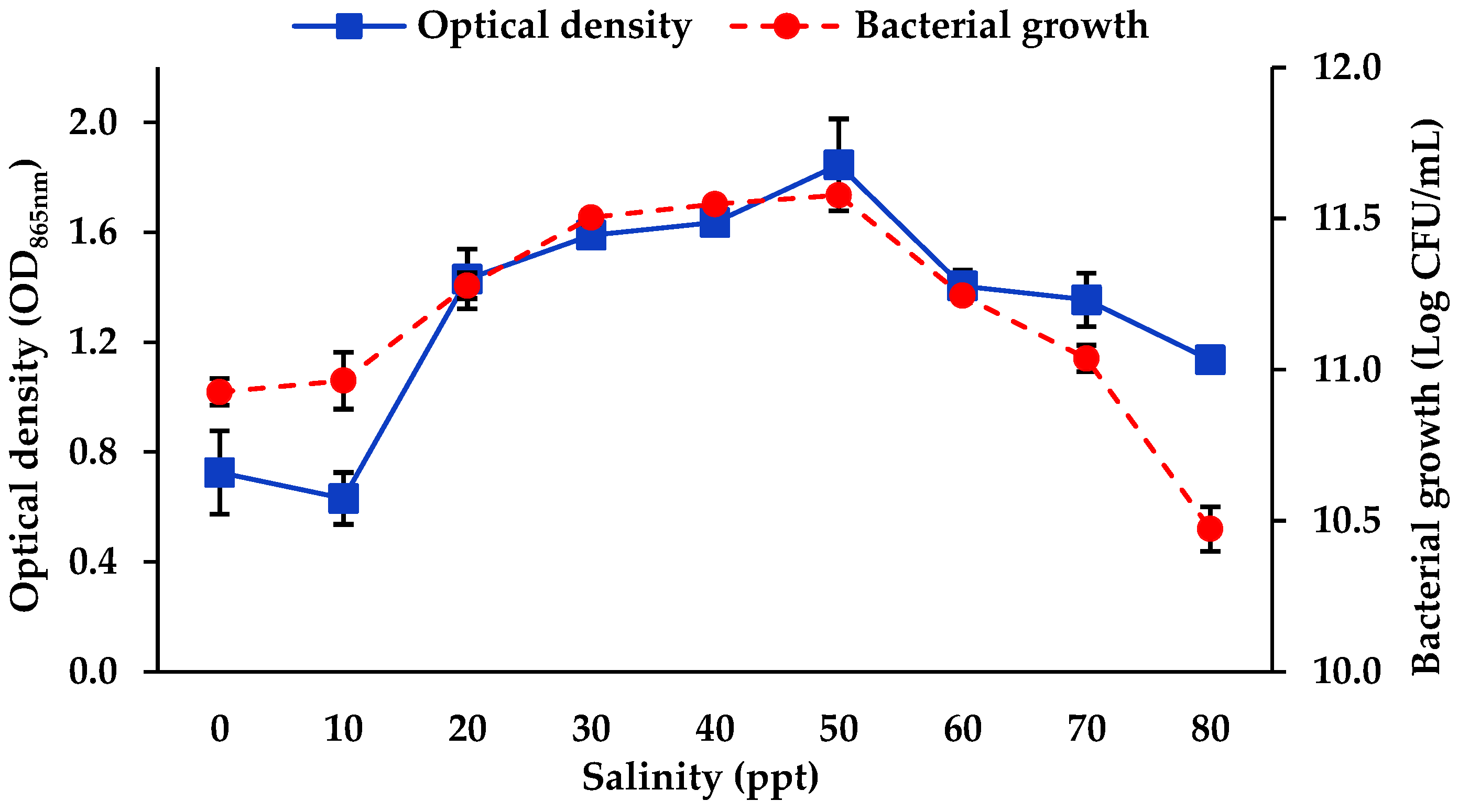
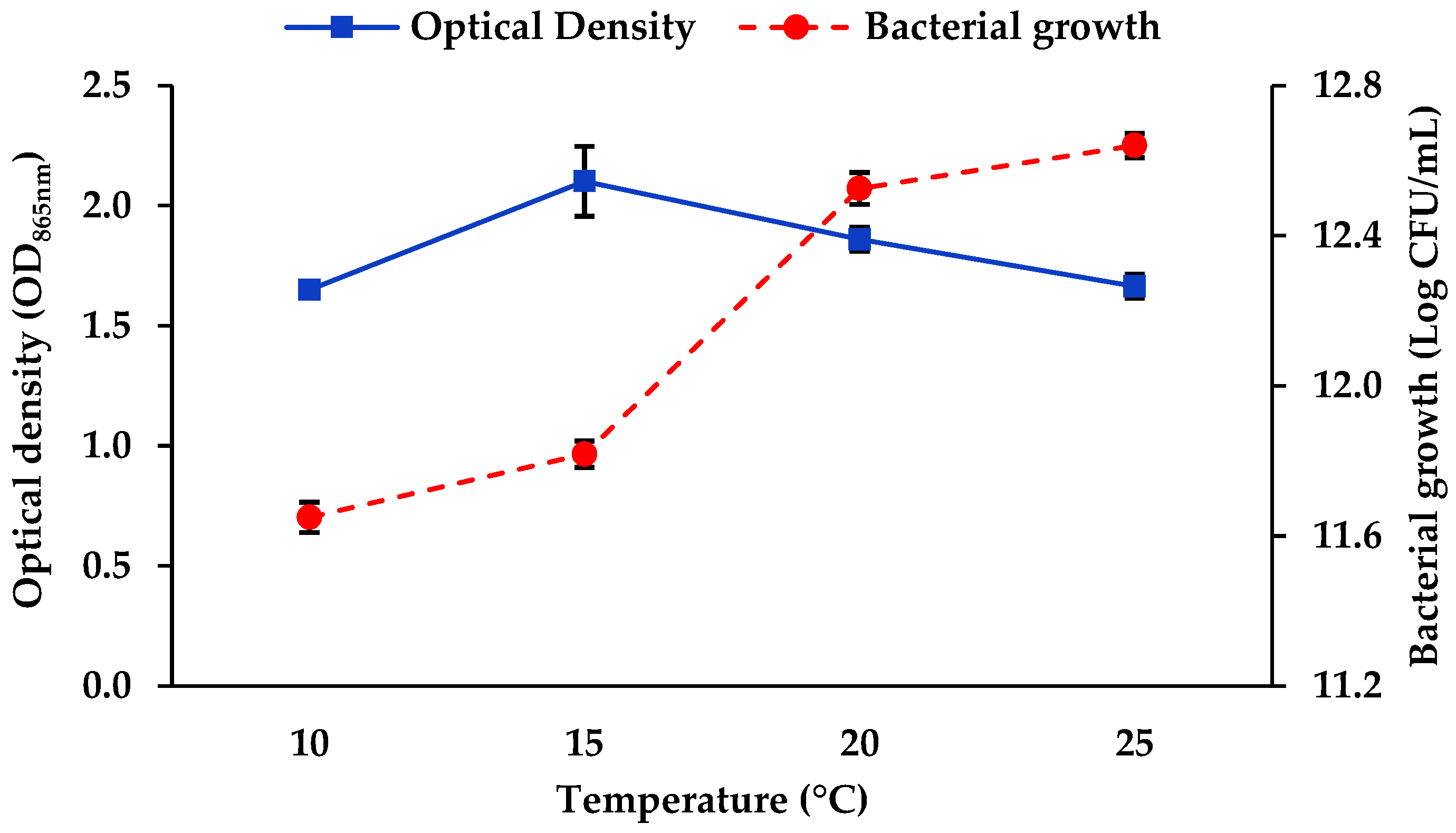
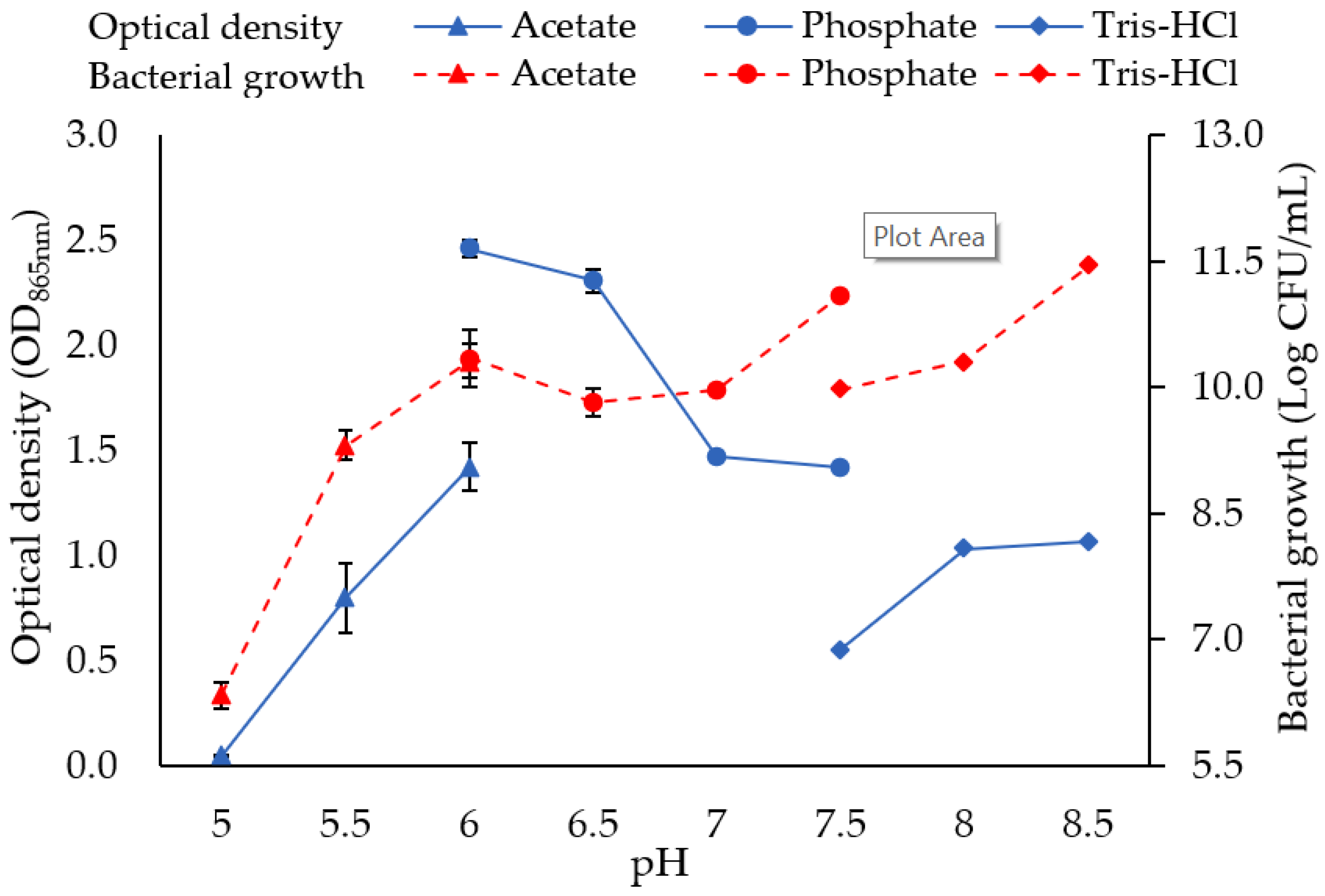
3.1.1. Salinity and Temperature
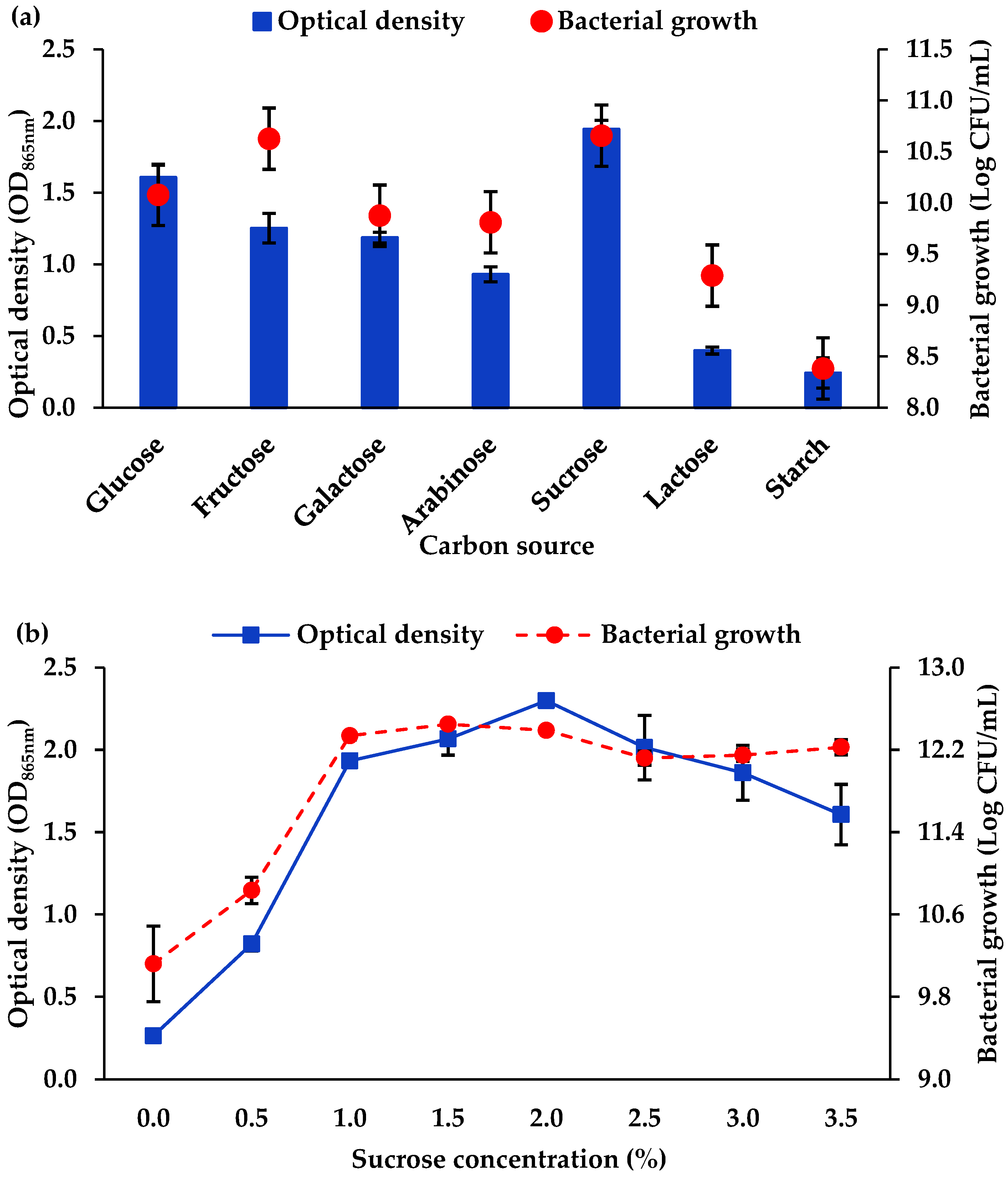
3.1.2. Carbon Source and Concentration
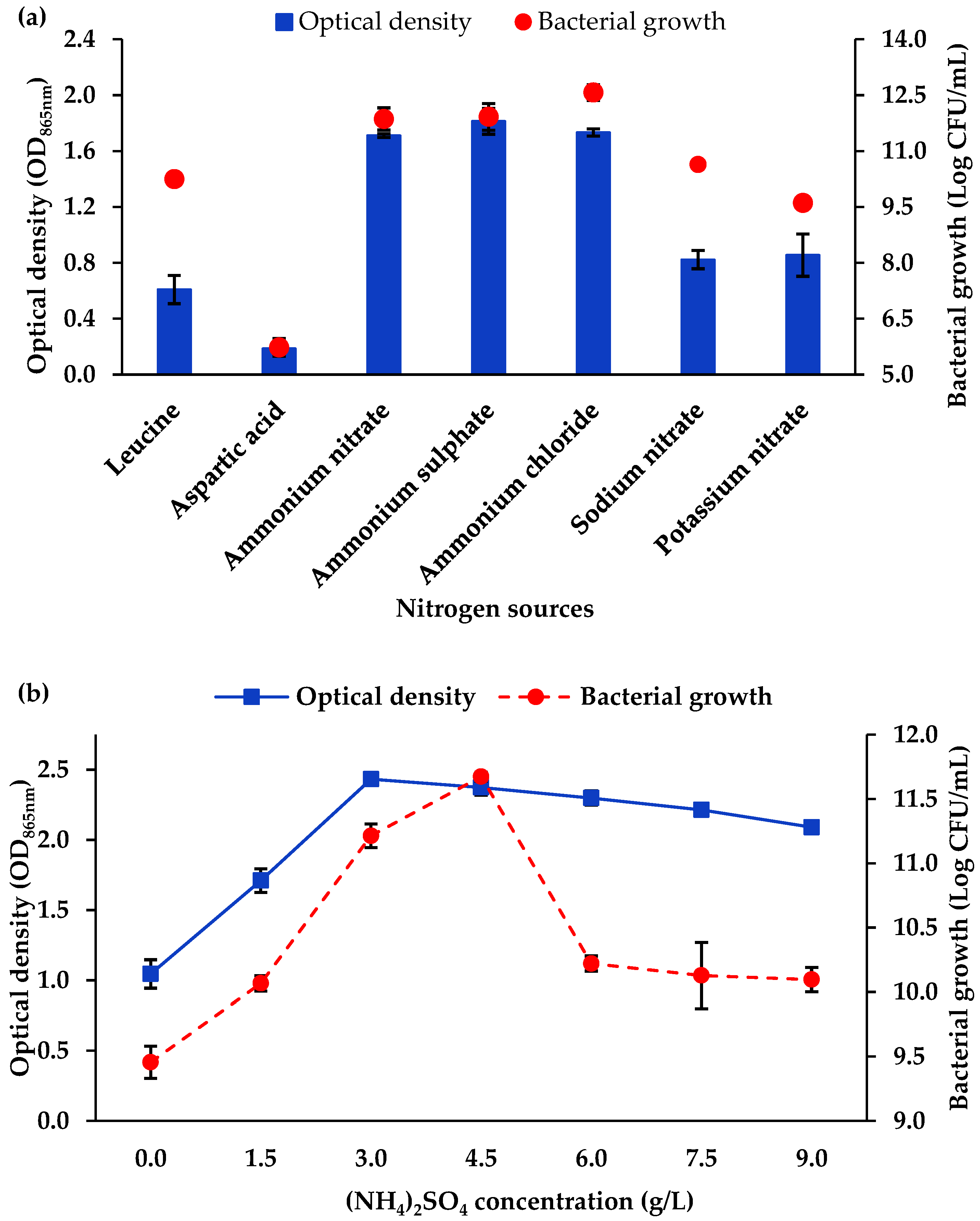
3.1.3. Nitrogen Source and Concentration
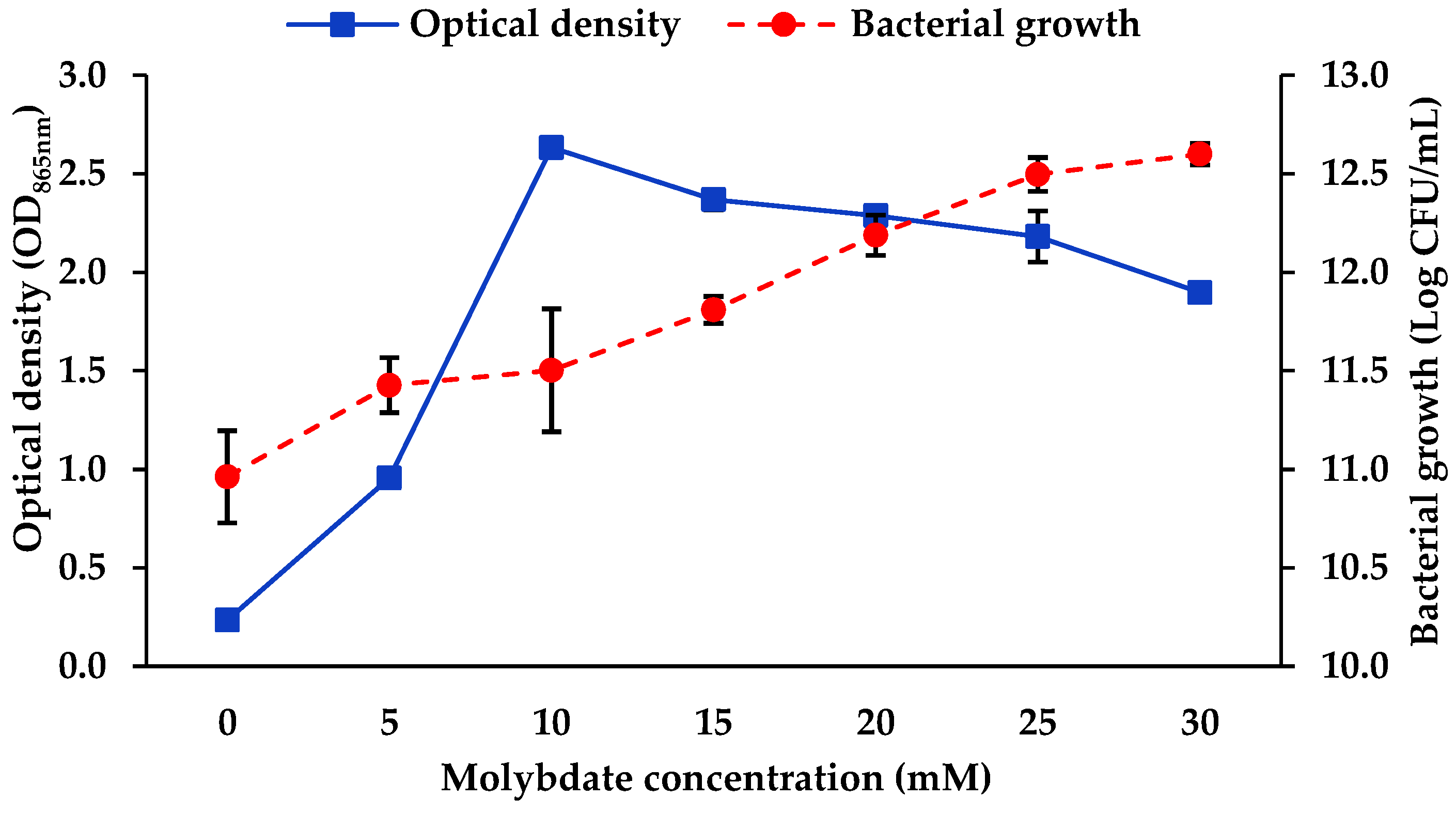
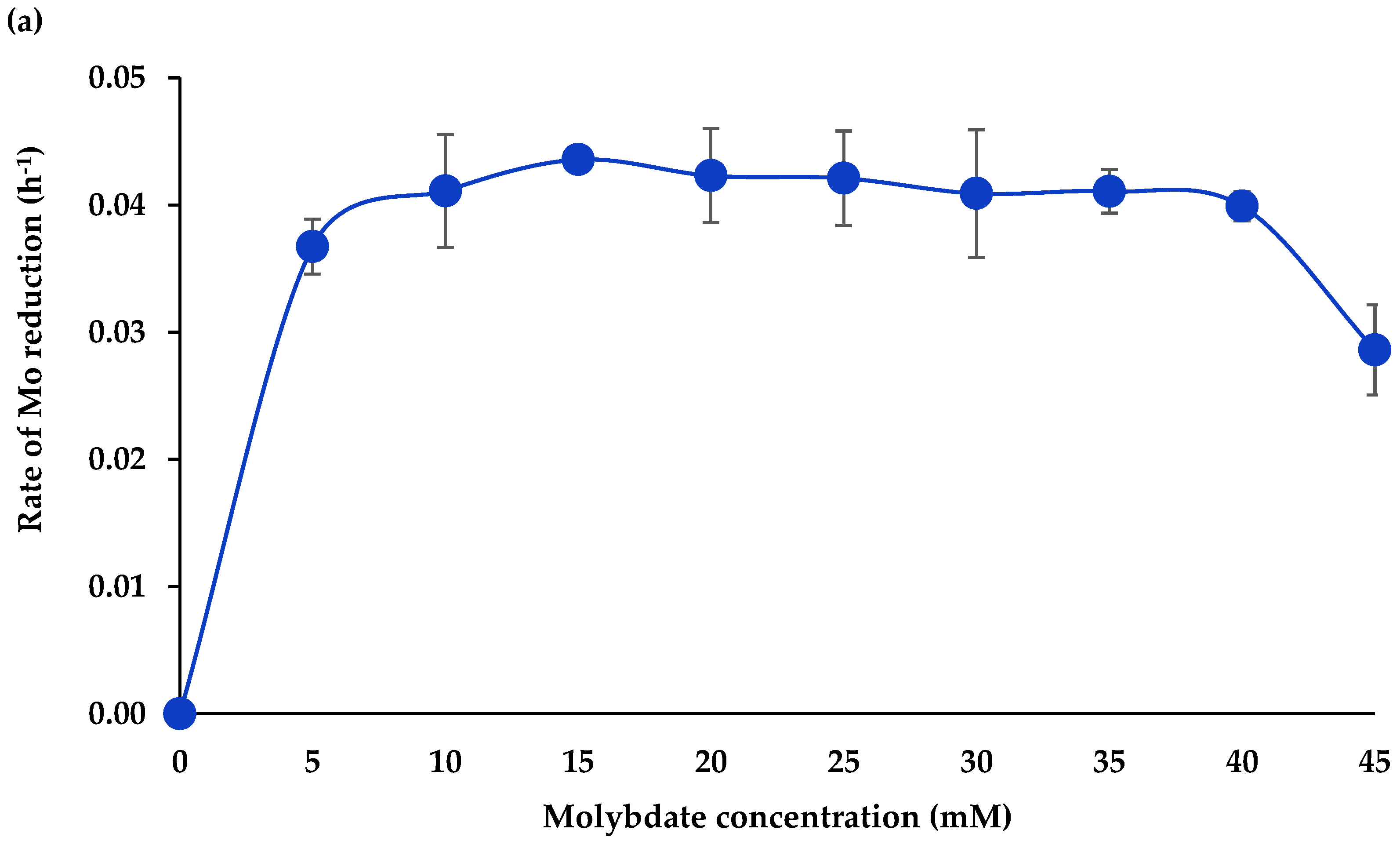
3.1.4. Molybdate Concentration
3.1.5. pH
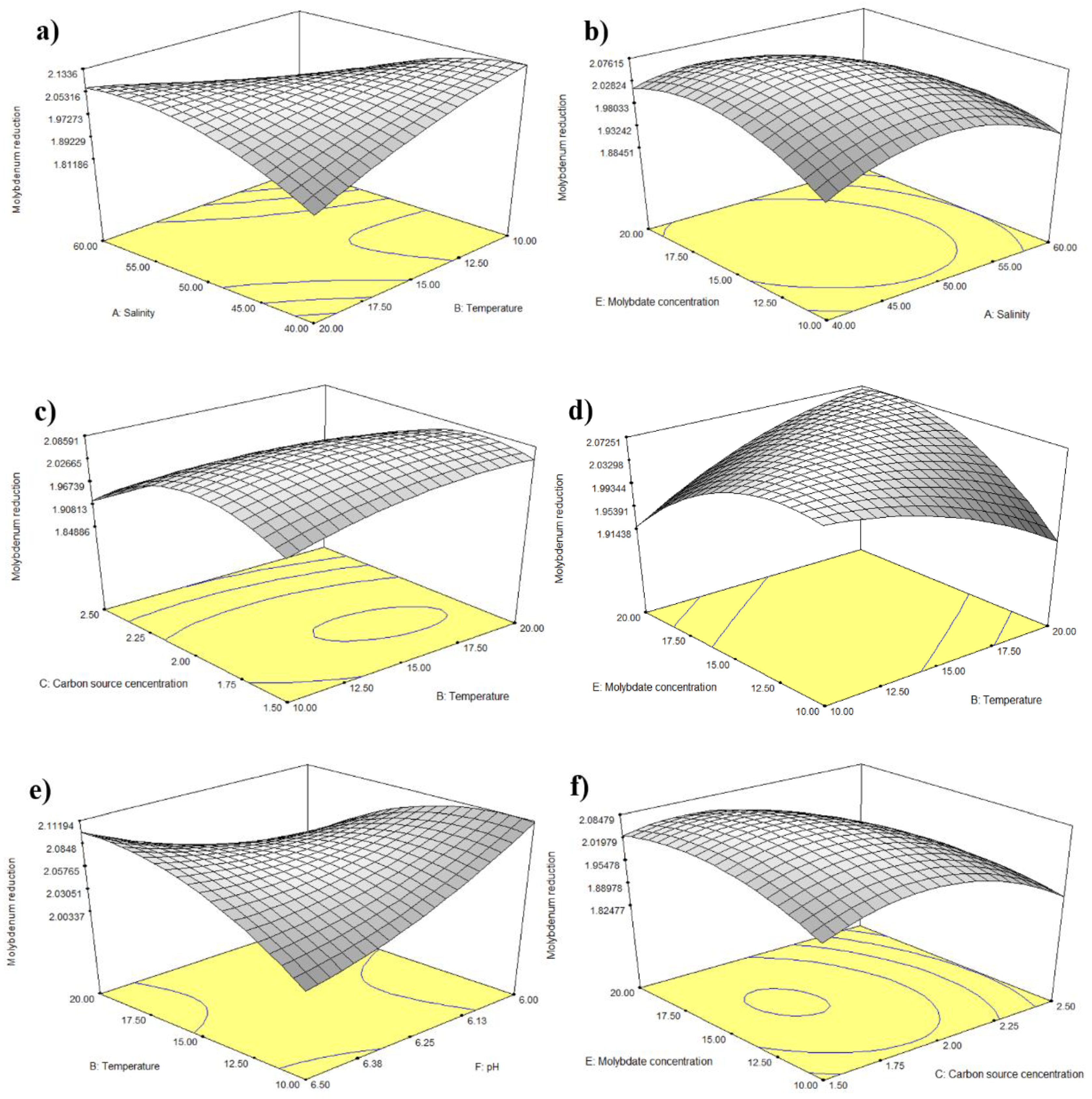
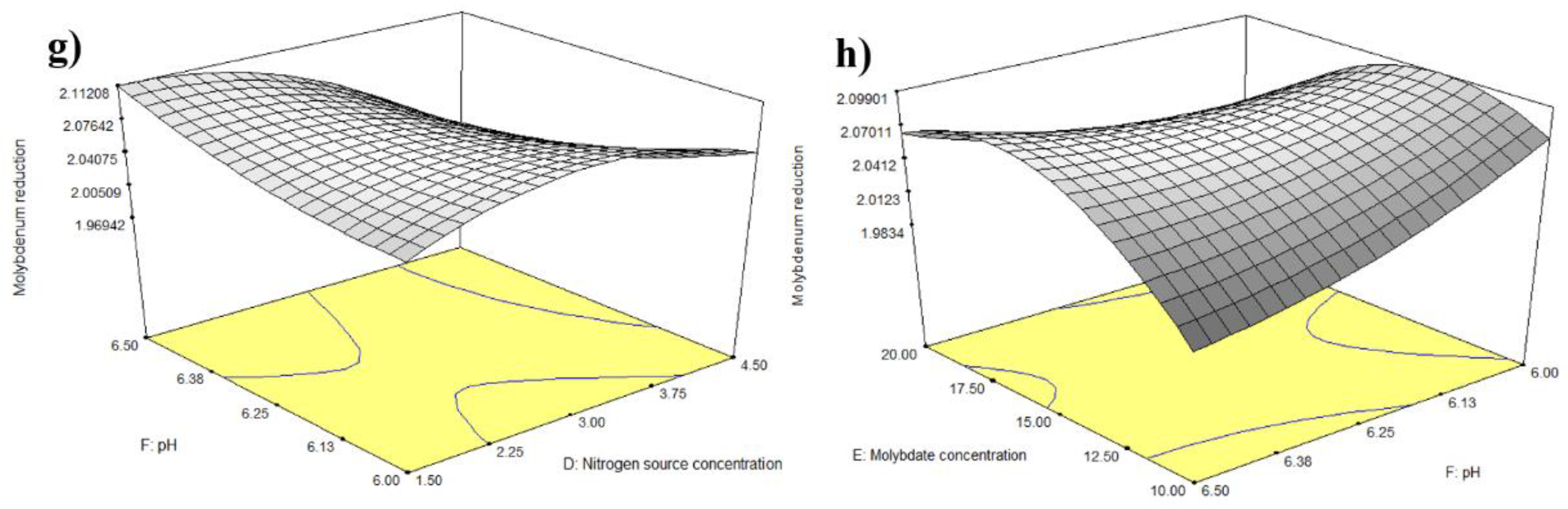
3.2. Statistical Optimisation of Mo Reduction Using RSM
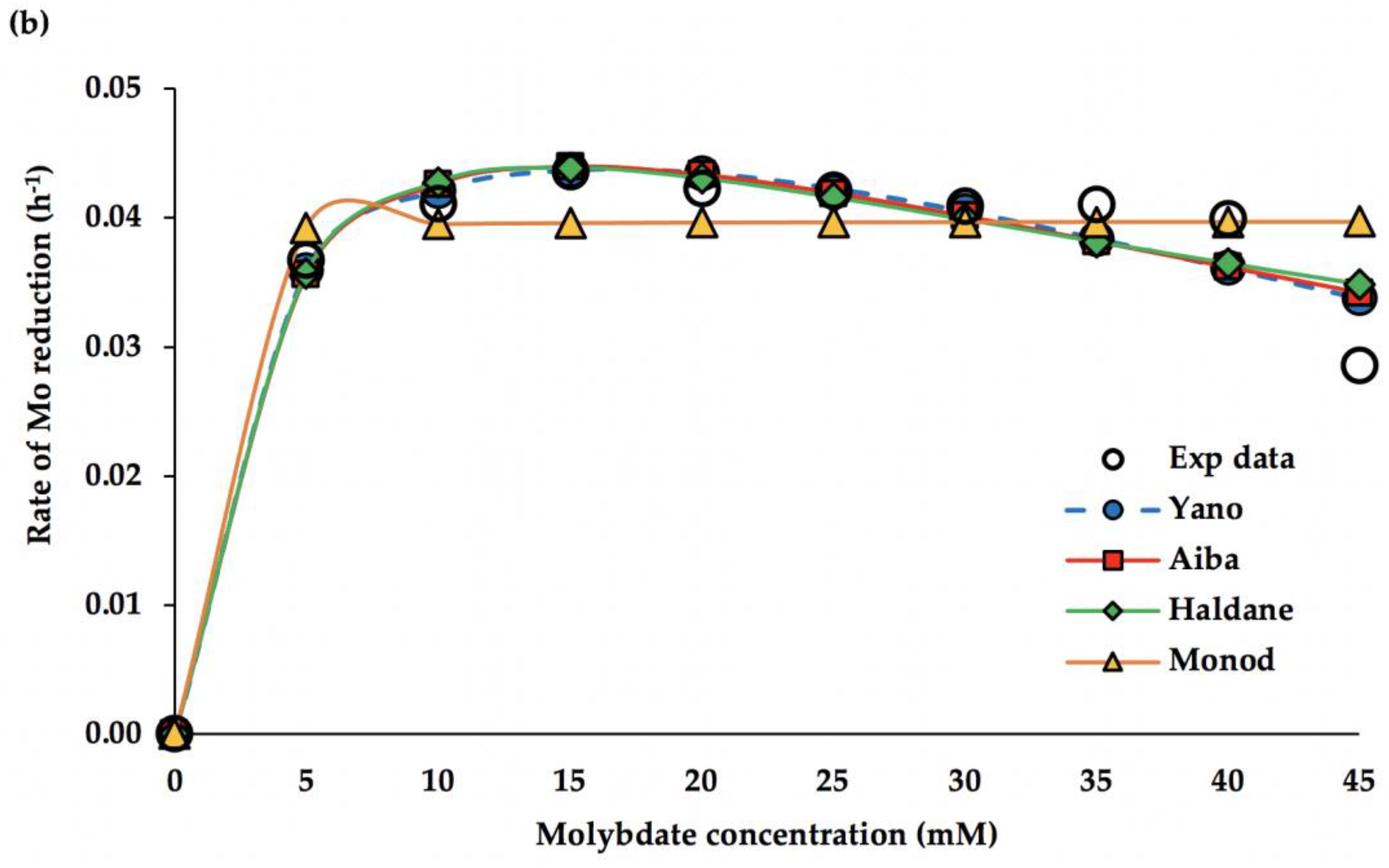
3.3. Kinetic Modelling
4. Discussion
4.1. Optimisation of Mo Reduction Using OFAT
4.2. Optimisation of Mo Reduction Using RSM
4.2.1. Statistical Analyses of RSM
4.2.2. Response Surface Plot of Pairwise Parameter Interactions
4.3. Comparison of Optimised Conditions between OFAT and RSM
4.4. Kinetic Study of Mo Reduction
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United Nations Environmental Protection/Global Program of Action. Why the Marine Environment Needs Protection from Heavy Metals; UNEP/GPA Coordination Officer, United Nations Environment Programme: Nairobi, Kenya, 2004. [Google Scholar]
- Zakaria, N.N.; Roslee, A.F.A.; Gomez-Fuentes, C.; Zulkharnain, A.; Abdulrasheed, M.; Sabri, S.; Ramirez-Moreno, N.; Calisto-Ulloa, N.; Ahmad, S.A. Kinetic studies of marine psychrotolerant microorganisms capable of degrading diesel in the presence of heavy metals. Rev. Mex. Ing. Quím. 2020, 19, 1375–1388. [Google Scholar] [CrossRef]
- Rowley, P.D.; Williams, P.L.; Pride, D.E. Petroleum and mineral resources of Antarctica. In Mineral Occurrences of Antarctica; U.S. Geological Survey Circular; Behrendt, J.C., Ed.; United States Department of the Interior: Washington, DC, USA, 1983; pp. 25–49. [Google Scholar]
- Hong, S.; Soyol-Erdene, T.O.; Hwang, H.J.; Hong, S.B.; Hur, S.D.; Motoyama, H. Evidence of global-scale As, Mo, Sb, and Tl atmospheric pollution in the Antarctic snow. Environ. Sci. Technol. 2012, 46, 11550–11557. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Welch, K.A.; Mohajerin, T.J.; Telfeyan, K.; Chevis, D.A.; Grimm, D.A.; Lyons, W.B.; White, C.D.; Johannesson, K.H. Comparison of arsenic and molybdenum geochemistry in meromictic lakes of the McMurdo dry valleys, Antarctica: Implications for oxyanion-forming trace element behavior in permanently stratified lakes. Chem. Geol. 2015, 404, 110–125. [Google Scholar] [CrossRef]
- Santos, I.R.; Silva-Filho, E.V.; Schaefer, C.E.G.R.; Albuquerque-Filho, M.R.; Campos, L.S. Heavy metal contamination in coastal sediments and soils near the Brazilian Antarctic station, King George island. Mar. Pollut. Bull. 2005, 50, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Barra, F.; Alcota, H.; Rivera, S.; Valencia, V.A.; Munizaga, F.; Maksaev, V. Timing and formation of porphyry Cu-Mo mineralization in the Chuquicamata district, northern Chile: New constraints from the Toki cluster. Miner. Deposita 2013, 48, 629–651. [Google Scholar] [CrossRef]
- Claridge, G.G.; Campbell, I.B.; Powell, H.K.; Amin, Z.H.; Balks, M.R. Heavy metal contamination in some soils of the McMurdo Sound region, Antarctica. Antarct. Sci. 1995, 7, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Novotny, J.A.; Peterson, C.A. Molybdenum. Adv. Nutr. 2018, 9, 272–273. [Google Scholar] [CrossRef]
- Bompart, G.; Pécher, C.; Prévot, D.; Girolami, J.P. Mild renal failure induced by subchronic exposure to molybdenum: Urinary kallikrein excretion as a marker of distal tubular effect. Toxicol. Lett. 1990, 52, 293–300. [Google Scholar] [CrossRef]
- Fungwe, T.V.; Buddingh, F.; Demick, D.S.; Lox, C.D.; Yang, M.T.; Yang, S.P. The role of dietary molybdenum on estrous activity, fertility, reproduction, and molybdenum and copper enzyme activities of female rats. Nutr. Res. 1990, 10, 515–524. [Google Scholar] [CrossRef]
- Kovalskiy, V.V.; Yarovaya, G.A.; Shmavonyan, D.M. Changes of purine metabolism in man and animals under conditions of molybdenum biogeochemical provinces. Zhurnal Obs. Biol. 1961, 22, 179–191. (In Russian) [Google Scholar]
- Walravens, P.A.; Moure-Eraso, R.; Solomons, C.C.; Chappell, W.R.; Bentley, G. Biochemical abnormalities in workers exposed to molybdenum dust. Arch. Environ. Health 1979, 34, 302–308. [Google Scholar] [CrossRef]
- Tengku-Mazuki, T.A.; Subramanian, K.; Zakaria, N.N.; Convey, P.; Khalil, K.A.; Lee, G.L.Y.; Zulkharnain, A.; Shaharuddin, N.A.; Ahmad, S.A. Optimization of phenol degradation by Antarctic bacterium Rhodococcus sp. Antarc. Sci. 2020, 32, 486–495. [Google Scholar] [CrossRef]
- Roslee, A.F.A.; Zakaria, N.N.; Convey, P.; Zulkharnain, A.; Lee, G.L.Y.; Gomez-Fuentes, C.; Ahmad, S.A. Statistical optimisation of growth and diesel degradation by the Antarctic bacterium, Rhodococcus sp. strain AQ5-07. Extremophiles 2020, 24, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Verasoundarapandian, G.; Darham, S.; Ahmad, S.A. Toxicity of molybdenum and microbial application in molybdenum reduction for bioremediation: A mini review. Malays. J. Biochem. Mol. Biol. 2019, 22, 46–51. [Google Scholar]
- Ahmad, S.A.; Shukor, M.Y.; Shamaan, N.A.; MacCormack, W.P.; Syed, M.A. Molybdate reduction to molybdenum blue by an Antarctic bacterium. BioMed Res. Int. 2013, 2013, 871941. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.M.; del Campillo-Campbell, A.; Villaret, D.B. Molybdate reduction by Escherichia coli K-12 and its chl mutants. Proc. Natl. Acad. Sci. USA 1985, 82, 227–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghani, B.; Takai, M.; Hisham, N.Z.; Kishimoto, N.; Ismail, A.K.M.; Tano, T.; Sugio, T. Isolation and characterization of a Mo6+ reducing bacterium. Appl. Environ. Microbiol. 1993, 59, 1176–1180. [Google Scholar] [CrossRef] [Green Version]
- Darham, S.; Gomez-Fuentes, C.; Zulkharnain, A.; Sabri, S.; Calisto-Ulloa, N.; Ramírez-Moreno, N.; Ahmad, S.A. Isolation and identification of molybdenum-reducing cold-adapted marine bacteria isolated from Bernardo O’Higgins riquelme base station, Antarctica. Malays. J. Biochem. Mol. Biol. 2019, 22, 8–15. [Google Scholar]
- Gupta, P.; Chaturvedi, P.; Pradhan, S.; Delille, D.; Shivaji, S. Marinomonas polaris sp. nov., a psychrohalotolerant strain isolated from coastal sea water off the subantarctic Kerguelen islands. Int. J. Syst. Bacteriol. 2006, 56, 361–364. [Google Scholar] [CrossRef]
- Zhang, D.C.; Li, H.R.; Xin, Y.H.; Liu, H.C.; Chen, B.; Chi, Z.M.; Zhou, P.J.; Yu, Y. Marinomonas arctica sp. nov., a psychrotolerant bacterium isolated from the Arctic. Int. J. Syst. Evolut. Microbiol. 2008, 58, 1715–1718. [Google Scholar] [CrossRef]
- Khuri, A.I.; Mukhopadhyay, S. Response surface methodology. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 128–149. [Google Scholar] [CrossRef]
- Ibrahim, S.; Khalil, K.A.; Zahri, K.N.M.; Gomez-Fuentes, C.; Convey, P.; Zulkharnain, A.; Sabri, S.; Alias, S.A.; González-Rocha, G.; Ahmad, S.A. Biosurfactant production and growth kinetics studies of the waste canola oil-degrading bacterium Rhodococcus erythropolis AQ5-07 from Antarctica. Molecules 2020, 25, 3878. [Google Scholar] [CrossRef] [PubMed]
- Abdulrasheed, M.; Zulkharnain, A.; Zakaria, N.N.; Roslee, A.F.A.; Khalil, K.A.; Napis, S.; Convey, P.; Gomez-Fuentes, C.; Ahmad, S.A. Response surface methodology optimization and kinetics of diesel degradation by a cold-adapted Antarctic bacterium, Arthrobacter sp. strain AQ5-05. Sustainability 2020, 12, 6966. [Google Scholar] [CrossRef]
- Haldane, J.B.S. Enzymes. In Monographs and Biochemistry; Plimmer, R.H.A., Hopkins, F.G., Eds.; Longmans and Green: London, UK, 1930; pp. 905–942. [Google Scholar]
- Monod, J. The growth of bacterial cultures. Annu. Rev. Microbiol. 1949, 3, 371–394. [Google Scholar] [CrossRef] [Green Version]
- Aiba, S.; Shoda, M.; Nagatani, M. Kinetics of product inhibition in alcohol fermentation. Biotechnol. Bioeng. 1968, 10, 845–864. [Google Scholar] [CrossRef]
- Yano, T.; Koga, S. Dynamic behavior of the chemostat subject to substrate inhibition. J. Gen. Appl. Microbiol. 1973, 19, 97–114. [Google Scholar] [CrossRef]
- Lee, G.L.Y.; Ahmad, S.A.; Yasid, N.A.; Zulkharnain, A.; Convey, P.; Johari, W.L.W.; Alias, S.A.; Gonzalez-Rocha, G.; Shukor, M.Y. Biodegradation of phenol by cold-adapted bacteria from Antarctic soils. Polar Biol. 2018, 41, 553–562. [Google Scholar] [CrossRef]
- Macián, M.C.; Arahal, D.R.; Garay, E.; Pujalte, M.J. Marinomonas aquamarina sp. nov., isolated from oysters and sea water. Syst. Appl. Microbiol. 2005, 28, 145–150. [Google Scholar] [CrossRef]
- Gordon, A.L.; Visbeck, M.; Huber, B. Export of Weddell sea deep and bottom water. J. Geophys. Res. 2001, 106, 9005–9017. [Google Scholar] [CrossRef]
- Sandle, T.; Skinner, K. Study of psychrophilic and psychrotolerant microorganisms isolated in cold rooms used for pharmaceutical processing. J. Appl. Microbiol. 2012, 114, 1166–1174. [Google Scholar] [CrossRef]
- Gilbert, J.A.; Davies, P.L.; Laybourn-Parry, J. A hyperactive, Ca2+-dependent antifreeze protein in an Antarctic bacterium. FEMS Microbiol. Lett. 2005, 245, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Bai, X.; Lai, Q.; Xie, Y.; Chen, X.; Shao, Z. Draft genome sequence of Marinomonas sp. strain D104, a plycyclic aromatic hydrocarbon-degrading bacterium from the deep-sea sediment of the Arctic Ocean. Genome Announc. 2014, 2, e01211–e01213. [Google Scholar] [CrossRef] [Green Version]
- Halmi, M.I.E.; Zuhainis, S.W.; Yusof, M.T.; Shaharuddin, N.A.; Helmi, W.; Shukor, Y.; Syed, M.A.; Ahmad, S.A. Hexavalent molybdenum reduction to Mo-blue by a sodium-dodecyl-sulfate-degrading Klebsiella oxytoca strain DRY14. BioMed Res. Int. 2013, 2013, e384541. [Google Scholar] [CrossRef] [PubMed]
- Sabullah, M.K.; Rahman, M.F.; Ahmad, S.A.; Sulaiman, M.R.; Shukor, M.S.; Shamaan, N.A.; Shukor, M.Y. Assessing resistance and bioremediation ability of Enterobacter sp. strain saw-1 on molybdenum in various heavy metals and pesticides. J. Math. Fund. Sci. 2017, 49, 193–210. [Google Scholar] [CrossRef] [Green Version]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial diversity in extreme marine habitats and their biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Adnan, A.S.M.; Zeid, I.M.A.; Ahmad, S.A.; Halmi, M.I.E.; Abdullah, S.R.S.; Masdor, N.A.; Shukor, M.S.; Shukor, M.Y. A molybdenum-reducing Bacillus sp. strain Zeid 14 in soils from Sudan that could grow on amides and acetonitrile. Malays. J. Soil Sci. 2016, 20, 111–134. [Google Scholar]
- Shukor, M.Y.; Habib, S.H.M.; Rahman, M.F.A.; Jirangon, H.; Abdullah, M.P.A.; Shamaan, N.A.; Syed, M.A. Hexavalent molybdenum reduction to molybdenum blue by S. marcescens strain Dr. Y6. Appl. Biochem. Biotechnol. 2008, 149, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Glenn, J.L.; Crane, F.L. Studies on metalloflavoproteins: V. The action of silicomolybdate in the reduction of cytochrome c by aldehyde oxidase. Biochim. Biophys. Acta 1956, 22, 111–115. [Google Scholar] [CrossRef]
- Krishnan, C.V.; Garnett, M.; Chu, B. Influence of pH and acetate on the self-assembly process of (NH4)42[MoVI72MoV60O372(CH3COO)30(H2O)72].ca.300H2O. Int. J. Electrochem. Sci. 2008, 3, 1299–1315. [Google Scholar]
- Sabullah, M.K.; Rahman, M.F.; Ahmad, S.A.; Sulaiman, M.R.; Shukor, M.S.; Shamaan, N.A.; Shukor, M.Y. Isolation and characterization of a molybdenum-reducing and glyphosate-degrading Klebsiella oxytoca strain saw-5 in soils from Sarawak. Agrivita 2016, 38, 1–13. [Google Scholar] [CrossRef]
- Rubio, V.; Linhares, F.; Solano, R.; Martín, A.C.; Iglesias, J.; Leyva, A.; Paz-Ares, J. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes Develop. 2020, 15, 2122–2133. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.; Skou, J.C.; Boyer, P.D. Adenosine Triphosphate. Available online: https://www.britannica.com/science/adenosine-triphosphate (accessed on 12 March 2020).
- Lee, J.Y.; Lim, H.S.; Yoon, H.I. Thermal characteristics of soil and water during summer at King Sejong station, King George Island, Antarctica. Geosci. J. 2016, 20, 503–516. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, X.; Ding, C.; Wang, Z.; Zhou, Q.; Pao, H.; Cai, W. Optimization of phenol degradation by Candida tropicalis Z-04 using Plackett-Burman design and response surface methodology. J. Environ. Sci. 2011, 23, 22–30. [Google Scholar] [CrossRef]
- Yan, N.; Marschner, P.; Cao, W.; Zuo, C.; Qin, W. Influence of salinity and water content on soil microorganisms. Int. Soil. Water Conserv. Res. 2015, 3, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Morita, R.Y.; Moyer, C.L. Origins of psychrophiles. In Encyclopedia of Biodiversity; Levin, S.A., Colwell, R., Daily, G., Lubchenco, J., Mooney, H.A., Schulze, E.D., Tilman, G.D., Eds.; Academic Press: San Diego, CA, USA, 2001; Volume 4, pp. 917–924. [Google Scholar]
- Sidgwick, N.V. The Chemical Elements and Their Compounds; Clarendon Press: Oxford, UK, 1984. [Google Scholar]
- Ivanova, E.P.; Onyshchenko, O.M.; Christen, R.; Lysenko, A.M.; Zhukova, N.V.; Shevchenko, L.S.; Kiprianova, E.A. Marinomonas pontica sp. nov., isolated from the Black Sea. Int. J. Syst. Evolut. Microbiol. 2005, 55, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, L.M.; Pfaender, F.K. The effect of inorganic and organic supplements on the microbial degradation of phenanthrene and pyrene in soils. Biodegradation 1997, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chaîneau, C.H.; Rougeux, G.; Yéprémian, C.; Oudot, J. Effects of nutrient concentration on the biodegradation of crude oil and associated microbial populations in the soil. Soil Biol. Biochem. 2005, 37, 1490–1497. [Google Scholar] [CrossRef]
- Dutta, K.; Dasu, V.; Mahanty, B.; Prabhu, A. Substrate inhibition growth kinetics for cutinase producing Pseudomonas cepacia using tomato-peel extracted cutin. Chem. Biochem. Eng. Q. 2015, 29, 437–445. [Google Scholar] [CrossRef]
- Muloiwa, M.; Nyende-Byakika, S.; Dinka, M. Comparison of unstructured kinetic bacterial growth models. S. Afr. J. Chem. Eng. 2020, 33, 141–150. [Google Scholar] [CrossRef]
- Othman, A.R.; Bakar, N.A.; Halmi, M.I.E.; Johari, W.L.W.; Ahmad, S.A.; Jirangon, H.; Syed, M.A.; Shukor, M.Y. Kinetics of molybdenum reduction to molybdenum blue by Bacillus sp. strain A.rzi. BioMed Res. Int. 2013, 2013, 371058. [Google Scholar] [CrossRef] [Green Version]
- Mulchandani, A.; Luong, H.T.; Groom, C. Substrate inhibition kinetics for microbial growth and synthesis of poly-β-hydroxybutyric acid by Alcaligenes eutrophus ATCC 17697. Appl. Microbiol. Technol. 1989, 33, 11–17. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Symbol | Experimental Values | ||||
|---|---|---|---|---|---|---|
| −2 | −1 | 0 | +1 | +2 | ||
| Salinity (ppt) | A | 21.72 | 40.0 | 50.0 | 60.0 | 78.28 |
| Temperature (°C) | B | 0.86 | 10.0 | 15.0 | 20.0 | 29.14 |
| Carbon concentration (%) | C | 0.59 | 1.5 | 2.0 | 2.5 | 3.41 |
| Nitrogen concentration (g/L) | D | −1.24 | 1.5 | 3.0 | 4.5 | 7.24 |
| Mo concentration (mM) | E | 0.86 | 10.0 | 15.0 | 20.0 | 29.14 |
| pH | F | 5.54 | 6.0 | 6.25 | 6.5 | 6.96 |
| Source | Sum of Squares | (Degrees of Freedom) DF | Mean Square | F-Value | Prob > F |
|---|---|---|---|---|---|
| Model | 4.12 | 16 | 0.16 | 15.46 | <0.0001 *** |
| A | 0.089 | 1 | 0.089 | 9.12 | 0.0035 ** |
| B | 0.00015 | 1 | 0.00015 | 0.015 | 0.9041 |
| C | 0.36 | 1 | 0.36 | 36.99 | <0.0001 *** |
| D | 0.09 | 1 | 0.09 | 9.21 | 0.0034 ** |
| E | 0.00067 | 1 | 0.00067 | 0.0066 | 0.7977 |
| F | 0.0045 | 1 | 0.0045 | 0.45 | 0.5067 |
| A2 | 0.54 | 1 | 0.54 | 53.63 | <0.0001 *** |
| B2 | 0.085 | 1 | 0.085 | 8.38 | 0.0053 ** |
| C2 | 0.97 | 1 | 0.97 | 95.69 | <0.0001 *** |
| D2 | 0.16 | 1 | 0.16 | 15.8 | 0.0002 *** |
| E2 | 0.37 | 1 | 0.37 | 0.37 | <0.0001 *** |
| F2 | 0.023 | 1 | 0.023 | 2.29 | 0.1359 |
| AB | 1.04 | 1 | 1.04 | 103.07 | <0.0001 *** |
| AC | 0.026 | 1 | 0.026 | 2.58 | 0.1135 |
| AD | 0.00165 | 1 | 0.00165 | 0.16 | 0.6873 |
| AE | 0.063 | 1 | 0.063 | 6.48 | 0.0132 * |
| AF | 0.00081 | 1 | 0.00081 | 0.08 | 0.7785 |
| BC | 0.082 | 1 | 0.082 | 8.38 | 0.0051 ** |
| BE | 0.34 | 1 | 0.34 | 34.75 | <0.0001 *** |
| BF | 0.14 | 1 | 0.14 | 14.01 | 0.0004 *** |
| CD | 0.0084 | 1 | 0.0084 | 0.83 | 0.3654 |
| CE | 0.073 | 1 | 0.073 | 7.27 | 0.0091 ** |
| DE | 0.012 | 1 | 0.012 | 1.15 | 0.2883 |
| DF | 0.091 | 1 | 0.091 | 9.29 | 0.0039 ** |
| EF | 0.088 | 1 | 0.088 | 9.03 | 0.0044 ** |
| Residual | 0.59 | 58 | 0.01 | ||
| Lack of Fit | 0.52 | 49 | 0.011 | 1.37 | 0.3194 |
| Pure Error | 0.069 | 9 | 0.0077 | ||
| Cor Total | 4.8 | 85 | |||
| Standard deviation | 0.10 | R2 | 0.8780 | ||
| Mean | 1.82 | Adjusted R2 | 0.8213 | ||
| Coefficient variance | 5.52 | Predicted R2 | 0.6272 | ||
| Predicted residual error sum of square | 1.79 | Adequate Precision | 19.347 | ||
| Model | DF | RMSE | R2 | Adj R2 | µmax (h−1) | Ks (mM) | Ki (mM) | K | AICc |
|---|---|---|---|---|---|---|---|---|---|
| Aiba | 7 | 0.0029 | 0.952 | 0.952 | 0.067 | 3.85 | 75.6 | - | −114.7 |
| Haldane | 7 | 0.0031 | 0.947 | 0.947 | 0.069 | 4.16 | 50.7 | - | −113.7 |
| Yano | 6 | 0.0030 | 0.952 | 0.952 | 0.053 | 2.33 | 8338 | 0.47 | −111.8 |
| Monod | 8 | 0.0045 | 0.896 | 0.883 | 0.040 | 0.06 | - | - | −107.7 |
| Variables | OFAT | RSM |
|---|---|---|
| Salinity (ppt) | 50.0 | 47.0 |
| Temperature (°C) | 15.0 | 16.0 |
| Carbon concentration (%) | 2.0 | 1.8 |
| Nitrogen concentration (g/L) | 3.0 | 2.25 |
| Molybdate concentration (mM) | 15.0 | 16.0 |
| pH (phosphate buffer) | 6.0 | 6.0 |
| Incubation period (day) | 8 | 4 |
| Mo-blue production (OD865nm) | 2.459 | 2.201 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darham, S.; Zahri, K.N.M.; Zulkharnain, A.; Sabri, S.; Gomez-Fuentes, C.; Convey, P.; Khalil, K.A.; Ahmad, S.A. Statistical Optimisation and Kinetic Studies of Molybdenum Reduction Using a Psychrotolerant Marine Bacteria Isolated from Antarctica. J. Mar. Sci. Eng. 2021, 9, 648. https://doi.org/10.3390/jmse9060648
Darham S, Zahri KNM, Zulkharnain A, Sabri S, Gomez-Fuentes C, Convey P, Khalil KA, Ahmad SA. Statistical Optimisation and Kinetic Studies of Molybdenum Reduction Using a Psychrotolerant Marine Bacteria Isolated from Antarctica. Journal of Marine Science and Engineering. 2021; 9(6):648. https://doi.org/10.3390/jmse9060648
Chicago/Turabian StyleDarham, Syazani, Khadijah Nabilah Mohd Zahri, Azham Zulkharnain, Suriana Sabri, Claudio Gomez-Fuentes, Peter Convey, Khalilah Abdul Khalil, and Siti Aqlima Ahmad. 2021. "Statistical Optimisation and Kinetic Studies of Molybdenum Reduction Using a Psychrotolerant Marine Bacteria Isolated from Antarctica" Journal of Marine Science and Engineering 9, no. 6: 648. https://doi.org/10.3390/jmse9060648
APA StyleDarham, S., Zahri, K. N. M., Zulkharnain, A., Sabri, S., Gomez-Fuentes, C., Convey, P., Khalil, K. A., & Ahmad, S. A. (2021). Statistical Optimisation and Kinetic Studies of Molybdenum Reduction Using a Psychrotolerant Marine Bacteria Isolated from Antarctica. Journal of Marine Science and Engineering, 9(6), 648. https://doi.org/10.3390/jmse9060648