
Discretization and Analysis of HIV-1 and HTLV-I Coinfection Model with Latent Reservoirs


Abstract
:1. Introduction
2. The Discrete-Time Model
3. Preliminaries
4. Equilibria
5. Global Stability
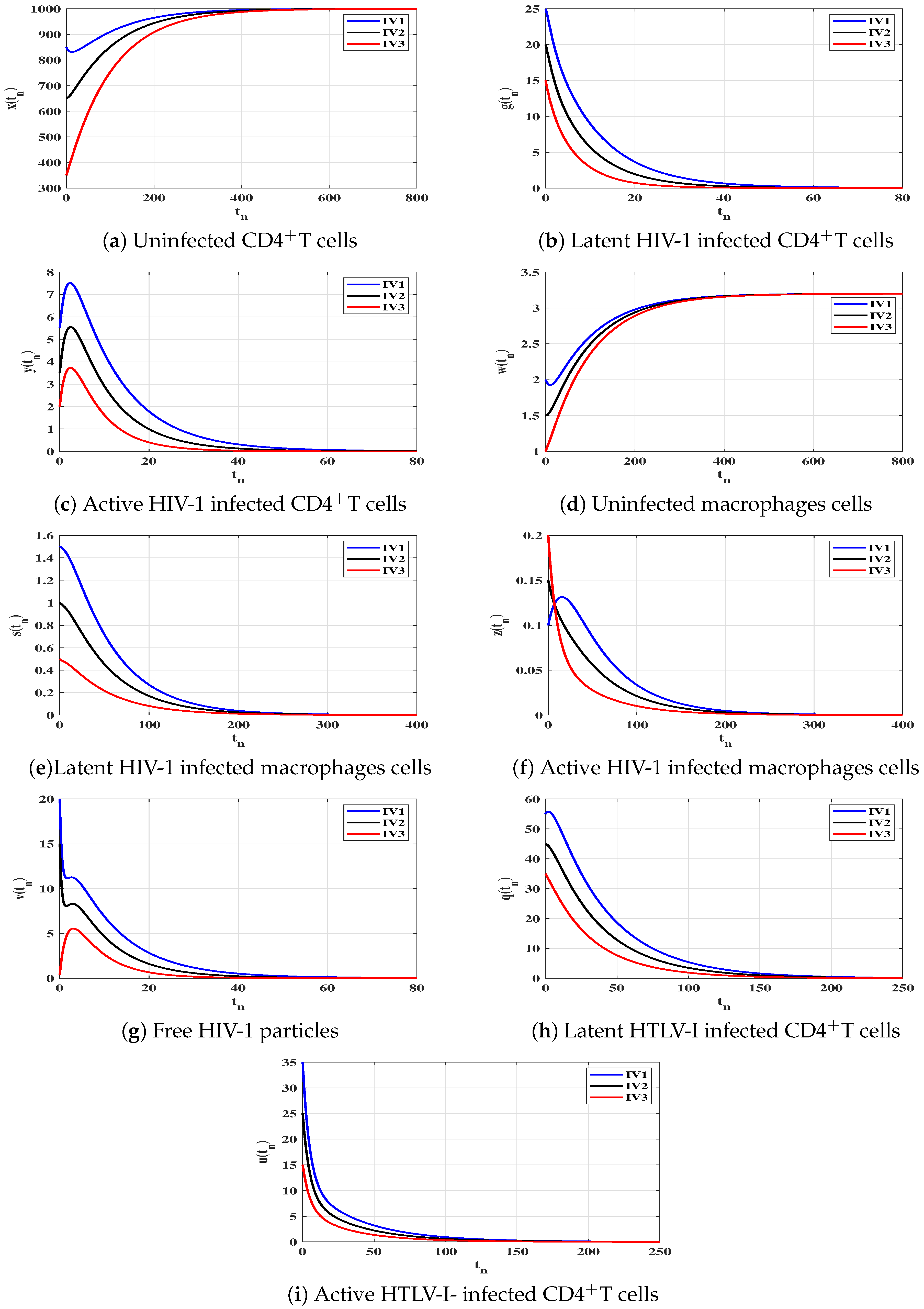
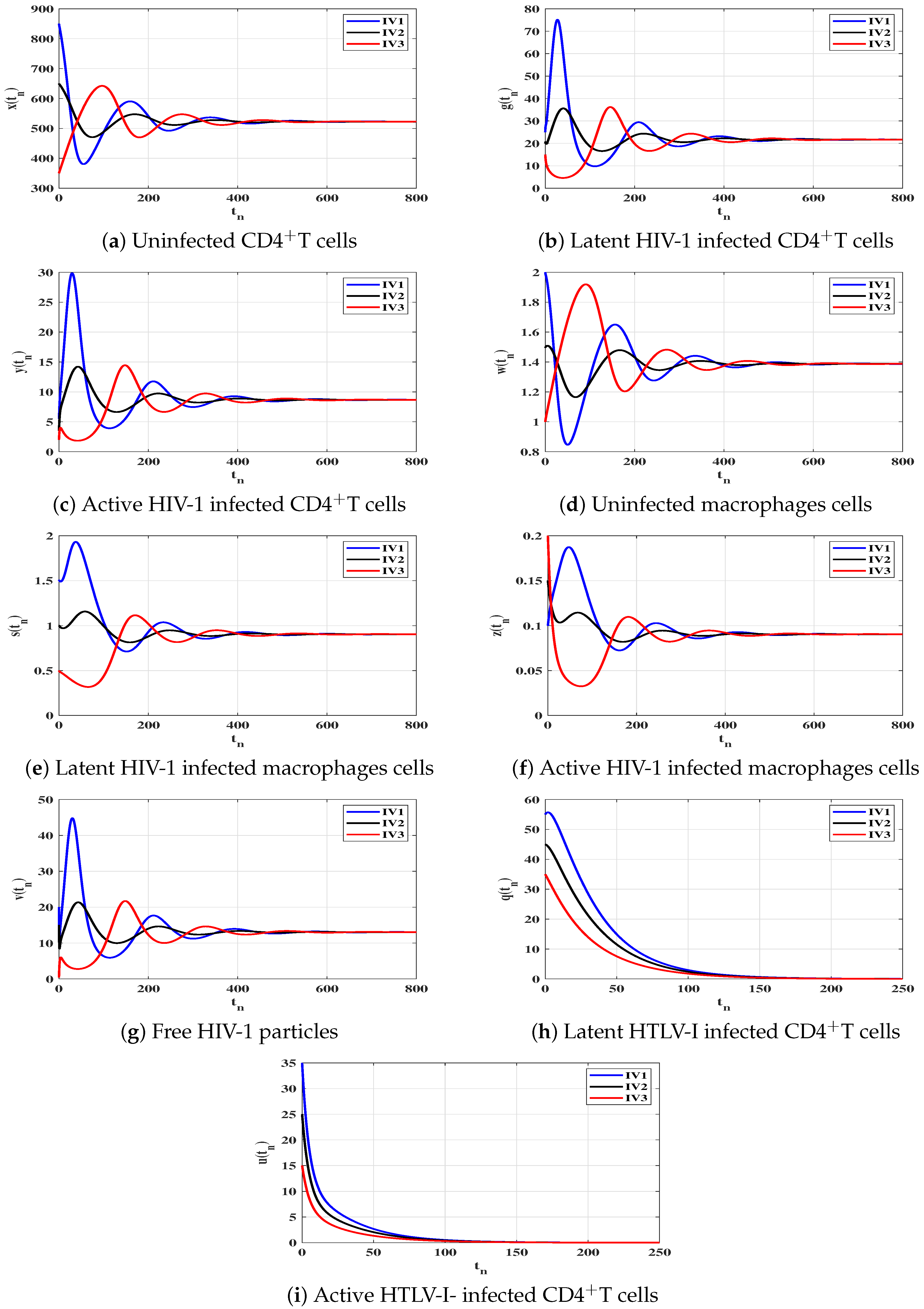
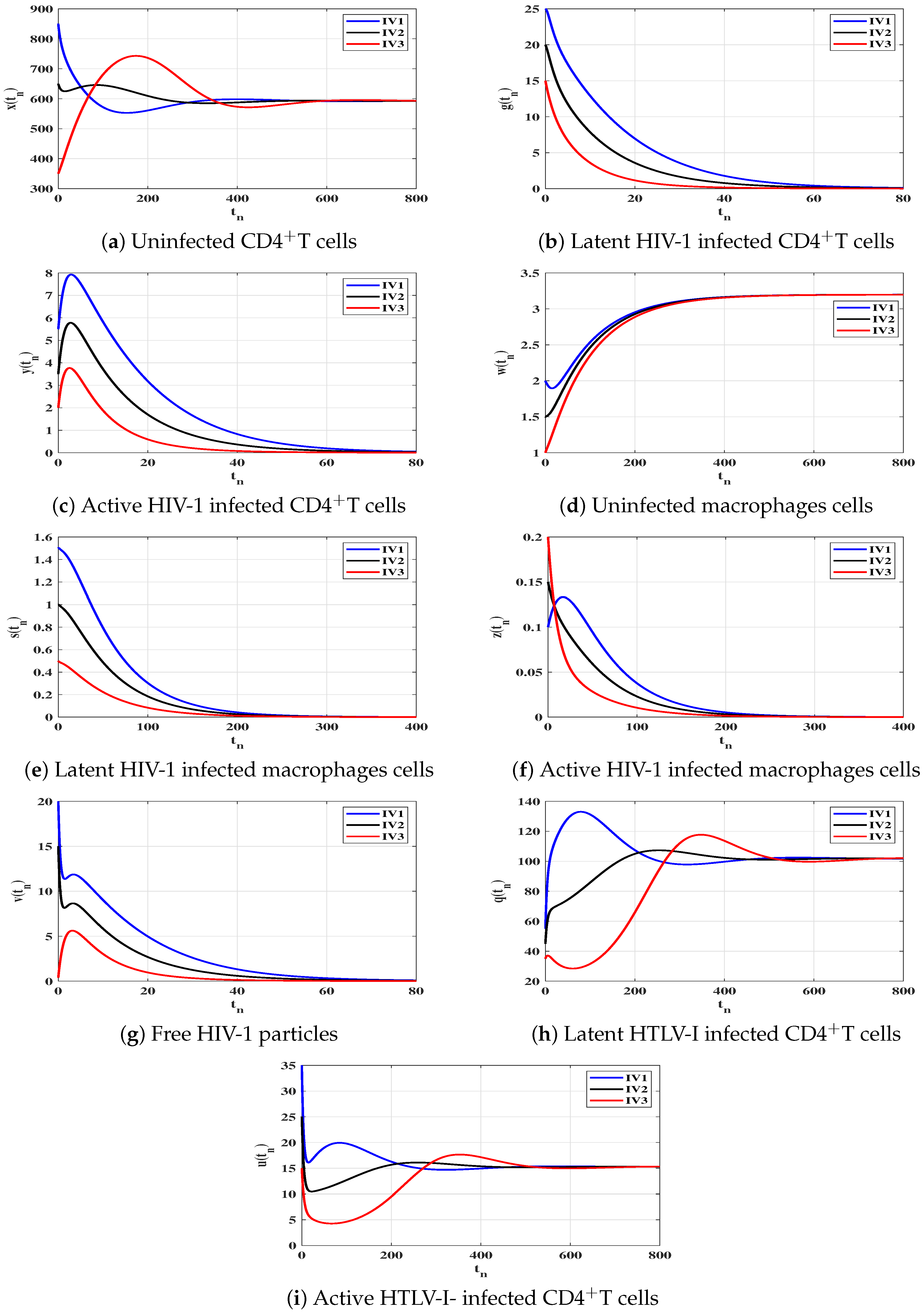
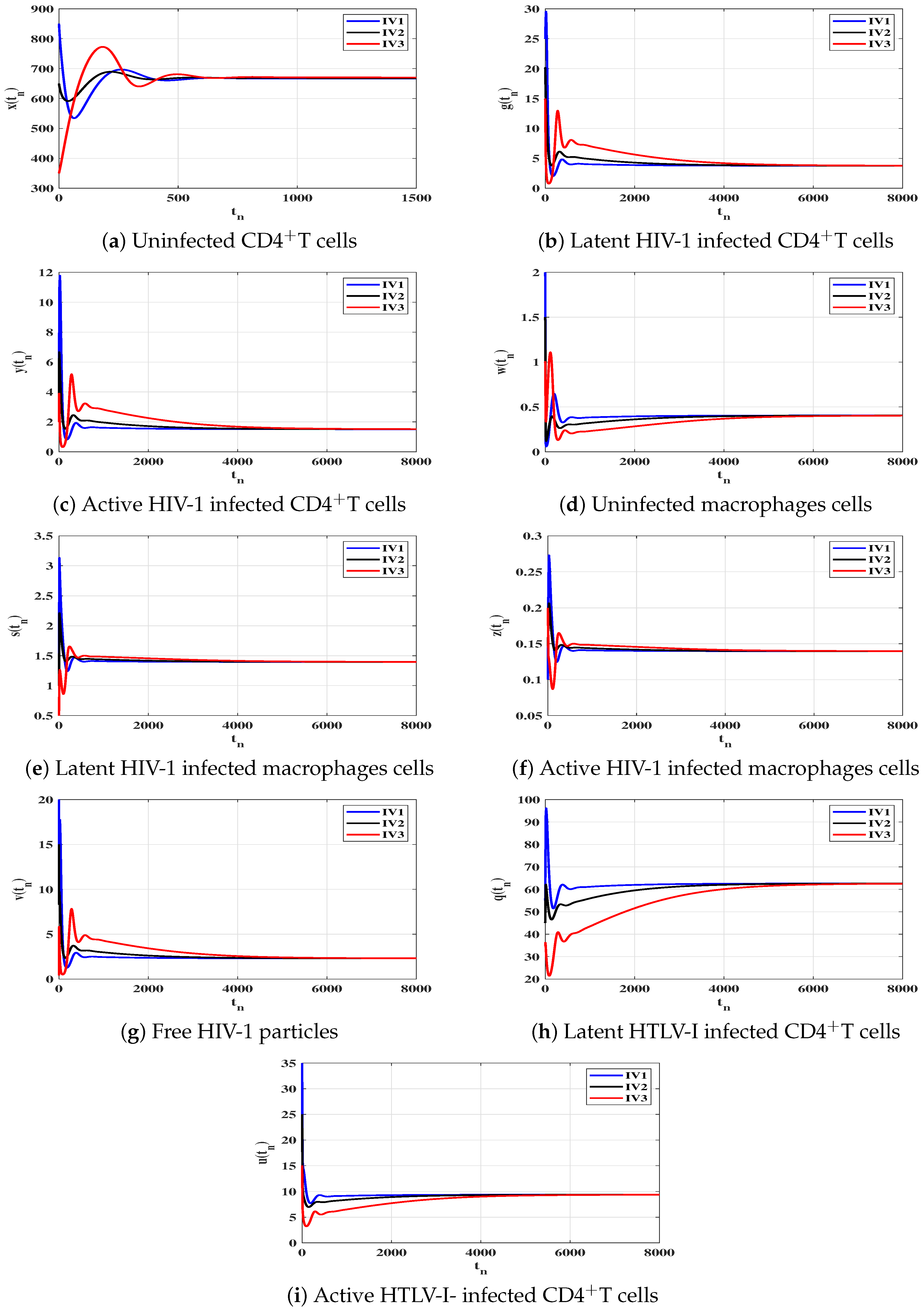
6. Numerical Simulations
6.1. Stability of the Equilibria
6.2. Impact of Latent Reservoirs on the HIV-1 and HTLV-I Co-Dynamics
Lengthening of the Latent Phase
7. Discussion and Conclusions
- always exists, further, if and , then is GAS. This result recommends that when and , both HIV-1 and HTLV-I infections will be cleared regardless of the starting points.
- exists when , and it is GAS when and . This result recommends obtain when and , the HIV-1 single-infection is always established regardless of the starting points.
- exists when , and it is GAS when and . This result recommends that when and , the HTLV-I single-infection is always established regardless of the starting points.
- exists, and it is GAS when , and . This result recommends that the HIV-1 and HTLV-I coinfection is always established regardless of the starting points.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Definition |
| AIDS | Acquired immunodeficiency syndrome |
| ATL | Adult T-cell leukemia |
| CTLs | Cytotoxic T lymphocytes |
| GAS | Globally asymptotically stable |
| HAM/TSP | HTLV-I-associated myelopathy/tropical spastic paraparesis |
| HBV | Hepatitis B virus |
| HCV | Hepatitis C virus |
| HIV-1 | Human immunodeficiency virus type 1 |
| HTLV-I | Human T-lymphotropic virus type I |
| LAS | Locally asymptotically stable |
| NSFD | Non-standard finite difference |
| IAV | Influenza A virus |
| RTI | Reverse transcriptase inhibitor |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| sup | Supremum (least upper bound) |
References
- Nowak, M.A.; Bangham, C.R.M. Population dynamics of immune responses to persistent viruses. Science 1996, 272, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Stilianakis, N.I.; Seydel, J. Modeling the T-cell dynamics and pathogenesis of HTLV-I infection. Bull. Math. Biol. 1999, 61, 935–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciupe, S.M.; Ribeiro, R.M.; Nelson, P.W.; Perelson, A.S. Modeling the mechanisms of acute hepatitis B virus infection. J. Theor. Biol. 2007, 247, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagawa, K.; Kuniya, T.; Nakaoka, S.; Asai, Y.; Watashi, K.; Iwami, S. Mathematical analysis of a transformed ODE from a PDE multiscale model of hepatitis C virus infection. Bull. Math. Biol. 2019, 81, 1427–1441. [Google Scholar] [CrossRef]
- Baccam, P.; Beauchemin, C.; Macken, C.A.; Hayden, F.G.; Perelson, A.S. Kinetics of influenza A virus infection in humans. J. Virol. 2006, 80, 7590–7599. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Vargas, E.A.; Velasco-Hernandez, J.X. In-host mathematical modelling of COVID-19 in humans. Annu. Rev. Control 2020, 50, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Pan, Y.; Wang, Q.; Miao, H.; Brown, A.N.; Rong, L. Modeling the viral dynamics of SARS-CoV-2 infection. Math. Biosci. 2020, 328, 108438. [Google Scholar] [CrossRef]
- Comez, M.C.; Yang, H.M. Mathematical model of the immune response to dengue virus. J. Appl. Math. Comput. 2020, 63, 455–478. [Google Scholar]
- Wang, Y.; Liu, X. Stability and Hopf bifurcation of a within-host chikungunya virus infection model with two delays. Math. Comput. Simul. 2017, 138, 31–48. [Google Scholar] [CrossRef]
- Best, K.; Perelson, A.S. Mathematical modeling of within-host Zika virus dynamics. Immunol. Rev. 2018, 285, 81–96. [Google Scholar] [CrossRef]
- Tang, B.; Xiao, Y.; Sander, B.; Kulkarni, M.A.; RADAM-LAC Research Team; Wu, J. Modelling the impact of antibody-dependent enhancement on disease severity of Zika virus and dengue virus sequential and co-infection. R. Soc. Open Sci. 2020, 7, 191749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elaiw, A.M.; AlShamrani, N.H. Analysis of a within-host HTLV-I/HIV-1 co-infection model with immunity. Virus Res. 2021, 295, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Elaiw, A.M.; AlShamrani, N.H. HTLV/HIV dual infection: Modeling and analysis. Mathematics 2021, 9, 51. [Google Scholar] [CrossRef]
- Pinky, L.; Dobrovolny, H.M. SARS-CoV-2 coinfections: Could influenza and the common cold be beneficial? J. Med. Virol. 2020, 92, 2623–2630. [Google Scholar] [CrossRef] [PubMed]
- Elaiw, A.M.; Alsulami, R.S.; Hobiny, A.D. Modeling and stability analysis of within-host IAV/SARS-CoV-2 coinfection with antibody immunity. Mathematics 2022, 10, 4382. [Google Scholar] [CrossRef]
- Agha, A.D.A.; Elaiw, A.M.; Ramadan, S.A.A.E. Stability analysis of within-host SARS-CoV-2/HIV coinfection model. Math. Methods Appl. Sci. 2022, 45, 11403–11422. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Shflot, A.S.; Hobiny, A.D. Global stability of delayed SARS-CoV-2 and HTLV-I coinfection models within a host. Mathematics 2022, 10, 4756. [Google Scholar] [CrossRef]
- Birger, R.; Kouyos, R.; Dushoff, J.; Grenfell, B. Modeling the effect of HIV coinfection on clearance and sustained virologic response during treatment for hepatitis C virus. Epidemics 2015, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nampala, H.; Livingstone, S.; Luboobi, L.; Mugisha, J.Y.T.; Obua, C.; Jablonska-Sabuka, M. Modelling hepatotoxicity and antiretroviral therapeutic effect in HIV/HBV coinfection. Math. Biosci. 2018, 302, 67–79. [Google Scholar] [CrossRef]
- Nelson, P.W.; Murray, J.D.; Perelson, A.S. A model of HIV-1 pathogenesis that includes an intracellular delay. Math. Biosci. 2000, 163, 201–215. [Google Scholar] [CrossRef]
- Nelson, P.W.; Perelson, A.S. Mathematical analysis of delay differential equation models of HIV-1 infection. Math. Biosci. 2002, 179, 73–94. [Google Scholar] [CrossRef]
- Perelson, A.S.; Nelson, P.W. Mathematical analysis of HIV-1 dynamics in vivo. SIAM Rev. 1999, 41, 3–44. [Google Scholar] [CrossRef] [Green Version]
- Lv, C.; Huang, L.; Yuan, Z. Global stability for an HIV-1 infection model with Beddington-DeAngelis incidence rate and CTL immune response. Commun. Nonlinear Sci. Numer. Simul. 2014, 19, 121–127. [Google Scholar] [CrossRef]
- Lin, J.; Xu, R.; Tian, X. Threshold dynamics of an HIV-1 model with both viral and cellular infections, cell-mediated and humoral immune responses. Math. Biosci. Eng. 2018, 16, 292–319. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, J. Threshold dynamics of a delayed nonlocal reaction-diffusion HIV infection model with both cell-free and cell-to-cell transmissions. J. Math. Anal. Appl. 2020, 488, 124047. [Google Scholar] [CrossRef]
- Feng, T.; Qiu, Z.; Meng, X.; Rong, L. Analysis of a stochastic HIV-1 infection model with degenerate diffusion. Appl. Math. Comput. 2019, 348, 437–455. [Google Scholar] [CrossRef]
- Callaway, D.S.; Perelson, A.S. HIV-1 infection and low steady state viral loads. Bull. Math. Biol. 2002, 64, 29–64. [Google Scholar] [CrossRef]
- Adams, B.M.; Banks, H.T.; Kwon, H.-D.; Tran, H.T. Dynamic multidrug therapies for HIV: Optimal and STI control approaches. Math. Biosci. Eng. 2004, 1, 223–241. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.M.; Banks, H.T.; Davidian, M.; Kwon, H.-D.; Tran, H.T.; Wynne, S.N.; Rosenberg, E.S. HIV dynamics: Modeling, data analysis, and optimal treatment protocols. J. Comput. Appl. Math. 2005, 184, 10–49. [Google Scholar] [CrossRef] [Green Version]
- Rong, L.; Perelson, A.S. Modeling HIV persistence, the latent reservoir, and viral blips. J. Theor. Biol. 2009, 260, 308–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perelson, A.S.; Essunger, P.; Cao, Y.; Vesanen, M.; Hurley, A.; Saksela, K.; Markowitz, M.; Ho, D.D. Decay characteristics of HIV-1-infected compartments during combination therapy. Nature 1997, 387, 188–191. [Google Scholar] [CrossRef]
- Korobeinikov, A. Global properties of basic virus dynamics models. Bull. Math. Biol. 2004, 66, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.G.; Maini, P.K. HTLV-I infection: A dynamic struggle between viral persistence and host immunity. J. Theor. Biol. 2014, 352, 92–108. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Ma, W. Global dynamics of a reaction and diffusion model for an HTLV-I infection with mitotic division of actively infected cells. J. Appl. Anal. Comput. 2017, 7, 899–930. [Google Scholar] [CrossRef]
- Li, F.; Ma, W. Dynamics analysis of an HTLV-1 infection model with mitotic division of actively infected cells and delayed CTL immune response. Math. Methods Appl. Sci. 2018, 41, 3000–3017. [Google Scholar] [CrossRef]
- Khajanchi, S.; Bera, S.; Roy, T.K. Mathematical analysis of the global dynamics of a HTLV-I infection model, considering the role of cytotoxic T-lymphocytes. Math. Comput. Simul. 2021, 180, 354–378. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Heffernan, J.M. Viral dynamics of an HTLV-I infection model with intracellular delay and CTL immune response delay. J. Math. Anal. Appl. 2018, 459, 506–527. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H.; Dahy, E.; Abdellatif, A. Stability of within host HTLV-I/HIV-1 co-infection in the presence of macrophages. Int. J. Biomath. 2022, 16, 2250066. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Aljahdali, A.K.; Hobiny, A.D. Dynamical [roperties of discrete-time HTLV-I and HIV-1 within-host coinfection model. Axioms 2023, 12, 201. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H.; Dahy, E.; Abdellatif, A.A.; Raezah, A.A. Effect of macrophages and latent reservoirs on the dynamics of HTLV-I and HIV-1 coinfection. Mathematics 2023, 11, 592. [Google Scholar] [CrossRef]
- Mickens, R.E. Nonstandard Finite Difference Models of Differential Equations; World Scientific: Singapore, 1994. [Google Scholar]
- Korpusik, A. A nonstandard finite difference scheme for a basic model of cellular immune response to viral infection. Commun. Nonlinear Sci. Numer. Simul. 2017, 43, 369–384. [Google Scholar] [CrossRef]
- Geng, Y.; Xu, J.; Hou, J. Discretization and dynamic consistency of a delayed and diffusive viral infection model. Appl. Math. Comput. 2018, 316, 282–295. [Google Scholar] [CrossRef]
- Xu, J.; Geng, Y. Stabilty preserving NSFD scheme for a delayed viral infection model with cell-to-cell transmission and general nonlinear incidence. J. Differ. Equ. Appl. 2017, 23, 893–916. [Google Scholar] [CrossRef]
- Manna, K. A non-standard finite difference scheme for a diffusive HBV infection model with capsids and time delay. J. Differ. Equ. Appl. 2017, 23, 1901–1911. [Google Scholar] [CrossRef]
- Vaz, S.; Torres, D.F.M. Discrete-time system of an intracellular delayed HIV model with CTL immune response. arXiv 2022, arXiv:2205.02199. [Google Scholar]
- Salman, S.M. A nonstandard finite difference scheme and optimal control for an HIV model with Beddington-DeAngelis incidence and cure rate. Eur. Phys. J. Plus 2020, 135, 808. [Google Scholar] [CrossRef]
- Liu, X.L.; Zhu, C.C. A non-standard finite difference scheme for a diffusive HIV-1 infection model with immune response and intracellular delay. Axioms 2022, 11, 129. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Alshaikh, M.A. Stability of discrete-time HIV dynamics models with three categories of infected CD4+ T-cells. Adv. Differ. Equ. 2019, 2019, 407. [Google Scholar] [CrossRef]
- Pasha, S.A.; Nawaz, Y.; Arif, M.S. On the nonstandard finite difference method for reaction–diffusion models. Chaos Solitons Fractals 2023, 166, 112929. [Google Scholar] [CrossRef]
- Maamar, M.H.; Ehrhardt, M.; Tabharit, L. A Nonstandard Finite Difference Scheme for a Time-Fractional Model of Zika Virus Transmission. 2022. Available online: https://www.imacm.uni-wuppertal.de/fileadmin/imacm/preprints/2022/imacm_22_21.pdf (accessed on 15 January 2023).
- Farooqi, A.; Ahmad, R.; Alotaibi, H.; Nofal, T.A.; Farooqi, R.; Khan, I. A comparative epidemiological stability analysis of predictor corrector type non-standard finite difference scheme for the transmissibility of measles. Results Phys. 2021, 21, 103756. [Google Scholar] [CrossRef]
- Shi, P.; Dong, L. Dynamical behaviors of a discrete HIV-1 virus model with bilinear infective rate. Math. Methods Appl. Sci. 2014, 37, 2271–2280. [Google Scholar] [CrossRef]
- Perelson, A.S.; Kirschner, D.E.; de Boer, R. Dynamics of HIV Infection of CD4+ T cells. Math. Biosci. 1993, 114, 81–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elaiw, A.M.; Raezah, A.A.; Azoz, S.A. Stability of delayed HIV dynamics models with two latent reservoirs and immune impairment. Adv. Differ. Equ. 2018, 50, 1–25. [Google Scholar] [CrossRef]
- Mohri, H.; Bonhoeffer, S.; Monard, S.; Perelson, A.S.; Ho, D. Rapid turnover of T lymphocytes in SIV-infected rhesus macaques. Science 1998, 279, 1223–1227. [Google Scholar] [CrossRef] [PubMed]
- Wodarz, D.; Nowak, M. Immune responses and viral phenotype: Do replication rate and cytopathogenicity influence virus load? J. Theor. Med. 1999, 2, 113. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Alshaikh, M.A. Stability preserving NSFD scheme for a general virus dynamics model with antibody and cell-mediated responses. Chaos Solitons Fractals 2020, 138, 109862. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Liu, L. Viral dynamics of an HIV model with latent infection incorporating antiretroviral therapy. Adv. Differ. Equ. 2016, 2016, 225. [Google Scholar] [CrossRef] [Green Version]
- Li, M.Y.; Lim, A.G. Modelling the role of Tax expression in HTLV-1 persisence in vivo. Bull. Math. Biol. 2011, 73, 3008–3029. [Google Scholar] [CrossRef] [PubMed]
- Raza, M.T.; Mizan, S.; Yasmin, F.; Akash, A.S.; Shahik, S. Epitope-based universal vaccine for Human T-lymphotropic virus-1 (HTLV-1). PLoS ONE 2021, 16, e0248001. [Google Scholar] [CrossRef]
- Beauchemin, A.A.C.; McSharry, J.J.; Drusano, G.L.; Nguyen, J.T.; Went, G.T.; Ribeiro, R.M.; Perelson, A.S. Modeling amantadine treatment of influenza A virus in vitro. J. Theor. Biol. 2008, 254, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Dobrovolny, H.M. Quantifying the effect of remdesivir in rhesus macaques infected with SARSCoV-2. Virology 2020, 550, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Bellomo, N.; Outada, N.; Soler, J.; Tao, Y.; Winkler, M. Chemotaxis and cross diffusion models in complex environments: Modeling towards a multiscale vision. Math. Model. Methods Appl. Sci. 2022, 32, 713–792. [Google Scholar] [CrossRef]
- Ali, I.; Khan, S.U. Dynamics and simulations of stochastic COVID-19 epidemic model using Legendre spectral collocation method. AIMS Math. 2023, 8, 4220–4236. [Google Scholar] [CrossRef]
- Qi, K.; Jiang, D.; Hayat, T.; Alsaedi, A. Virus dynamic behavior of a stochastic HIV/AIDS infection model including two kinds of target cell infections and CTL immune. Math. Comput. Simulat. 2021, 188, 548–570. [Google Scholar] [CrossRef]
- Ali, I.; Khan, S.U. Asymptotic behavior of three connected stochastic delay neoclassical growth systems using spectral technique. Mathematics 2022, 10, 3639. [Google Scholar] [CrossRef]
- Ali, I.; Khan, S.U. Threshold of stochastic SIRS epidemic model from infectious to susceptible class with saturated incidence rate using spectral method. Symmetry 2022, 14, 1838. [Google Scholar] [CrossRef]
- Chatterjee, A.N.; Basir, F.A.; Almuqrin, M.A.; Mondal, J.; Khan, I. SARS-CoV-2 infection with lytic and nonlytic immune responses: A fractional order optimal control theoretical study. Results Phys. 2021, 26, 104260. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Source |
|---|---|---|
| 10 cells mm day | [27,35,54] | |
| cells mm day | [28,29] | |
| day | [20,22] | |
| day | [13,55] | |
| day | [27,56] | |
| day | [28,29] | |
| 6 viruses cells | [55] | |
| 6 viruses cells | [55] | |
| 2 day | [13,57] | |
| day | [13,34] | |
| h | [58] | |
| (varied) viruses mm day | Assumed | |
| (varied) viruses mm day | Assumed | |
| (varied) cells mm day | Assumed | |
| day | [54,59] | |
| day | [55] | |
| day | [35] | |
| day | [59] | |
| day | Assumed | |
| day | [60] |
| Case | Equilibrium Point | Stability | |
|---|---|---|---|
| (I) | |||
| (II) | |||
| (III) | |||
| (IV) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elaiw, A.M.; Aljahdali, A.K.; Hobiny, A.D. Discretization and Analysis of HIV-1 and HTLV-I Coinfection Model with Latent Reservoirs. Computation 2023, 11, 54. https://doi.org/10.3390/computation11030054
Elaiw AM, Aljahdali AK, Hobiny AD. Discretization and Analysis of HIV-1 and HTLV-I Coinfection Model with Latent Reservoirs. Computation. 2023; 11(3):54. https://doi.org/10.3390/computation11030054
Chicago/Turabian StyleElaiw, Ahmed M., Abdualaziz K. Aljahdali, and Aatef D. Hobiny. 2023. "Discretization and Analysis of HIV-1 and HTLV-I Coinfection Model with Latent Reservoirs" Computation 11, no. 3: 54. https://doi.org/10.3390/computation11030054
APA StyleElaiw, A. M., Aljahdali, A. K., & Hobiny, A. D. (2023). Discretization and Analysis of HIV-1 and HTLV-I Coinfection Model with Latent Reservoirs. Computation, 11(3), 54. https://doi.org/10.3390/computation11030054