An Acinetobacter non-baumannii Population Study: Antimicrobial Resistance Genes (ARGs)

 ,
,  and
and 
Abstract
:1. Introduction
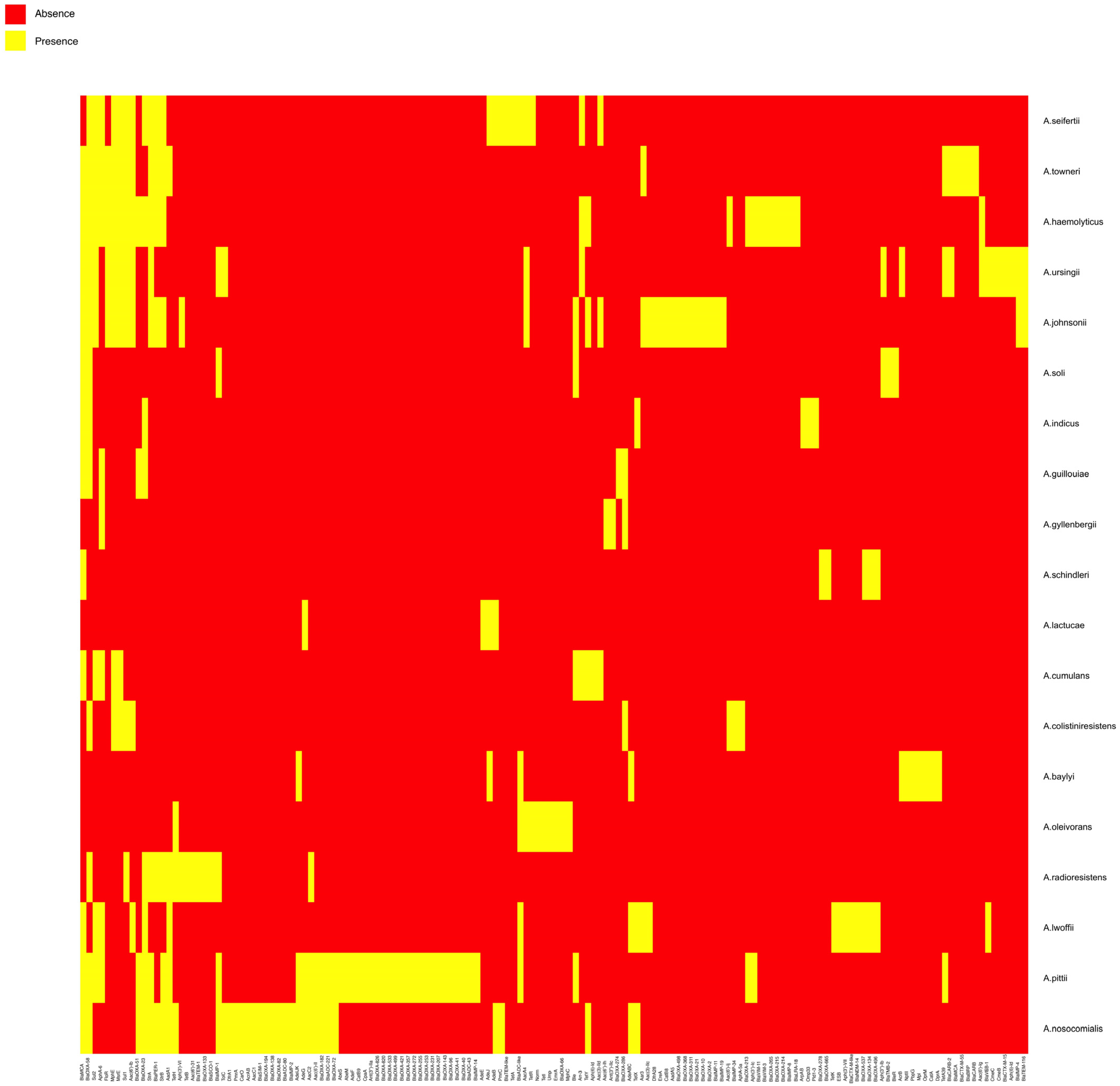
2. Results and Discussion
3. Materials and Methods
3.1. Literature Search
3.2. Genome Sequences Collection and Antibiotic Resistance Genes Prediction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- CDC. Antibiotic Resistance Threats in the United States; Centers for Disease Control: Atlanta, GA, USA, 2019. [Google Scholar]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and pathophysiological overview of Acinetobacter infections: A century of challenges. Clin. Microbiol. Rev. 2017, 30, 409–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavares, L.C.B.; Cunha, M.P.V.; Vasconcellos, F.M.; Bertani, A.M.J.; Barcellos, T.A.F.; Bueno, M.S.; Santos, C.A.; Sant’Ana, D.A.; Ferreira, A.M.; Mondelli, A.L.; et al. Genomic and clinical characterization of IMP-1-producing multidrug-resistant Acinetobacter bereziniae isolates from bloodstream infections in a brazilian tertiary hospital. Microb. Drug Resist. 2020. [Google Scholar] [CrossRef] [PubMed]
- Al Atrouni, A.; Joly-Guillou, M.L.; Hamze, M.; Kempf, M. Reservoirs of non-baumannii Acinetobacter species. Front. Microbiol. 2016, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, Z.; Zhang, X. blaNDM-1-carrying Acinetobacter johnsonii detected in hospital sewage. J. Antimicrob. Chemother. 2013, 68, 1007–1010. [Google Scholar] [CrossRef] [Green Version]
- Montana, S.; Palombarani, S.; Carulla, M.; Kunst, A.; Rodriguez, C.H.; Nastro, M.; Vay, C.; Ramirez, M.S.; Almuzara, M. First case of bacteraemia due to Acinetobacter schindleri harbouring blaNDM-1 in an immunocompromised patient. New Microbes New Infect. 2018, 21, 28–30. [Google Scholar] [CrossRef]
- Kiyasu, Y.; Hitomi, S.; Funayama, Y.; Saito, K.; Ishikawa, H. Characteristics of invasive Acinetobacter infection: A multicenter investigation with molecular identification of causative organisms. J. Infect. Chemother. 2020, 26, 475–482. [Google Scholar] [CrossRef]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM metallo-beta-lactamases and their bacterial producers in health care settings. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Bonomo, R.A.; Tolmasky, M.E. Carbapenemases: Transforming Acinetobacter baumannii into a yet more dangerous menace. Biomolecules 2020, 10, 720. [Google Scholar] [CrossRef]
- Poirel, L.; Marque, S.; Heritier, C.; Segonds, C.; Chabanon, G.; Nordmann, P. OXA-58, a novel class D {beta}-lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2005, 49, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Dozzo, P.; Moser, H.E. New aminoglycoside antibiotics. Expert Opin. Ther. Pat. 2010, 20, 1321–1341. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.; Perez, F.; Bonomo, R.A. Carbapenem-resistant Enterobacteriaceae and Acinetobacter baumannii: Assessing their impact on organ transplantation. Curr. Opin. Organ. Transplant. 2010, 15, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updates 2010, 13, 151–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delmar, J.A.; Su, C.C.; Yu, E.W. Bacterial multidrug efflux transporters. Ann. Rev. Biophys. 2014, 43, 93–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolmasky, M.E. Strategies to prolong the useful life of existing antibiotics and help overcoming the antibiotic resistance crisis. In Frontiers in Clinical Drug Research-Anti Infectives; Atta-ur-Rhaman, Ed.; Bentham Books: Sharjah, UAE, 2017; Volume 1, pp. 1–27. [Google Scholar]
- Coyne, S.; Courvalin, P.; Perichon, B. Efflux-mediated antibiotic resistance in Acinetobacter spp. Antimicrob. Agents Chemother. 2011, 55, 947–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damier-Piolle, L.; Magnet, S.; Bremont, S.; Lambert, T.; Courvalin, P. AdeIJK, a resistance-nodulation-cell division pump effluxing multiple antibiotics in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2008, 52, 557–562. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, E.; Nikaido, H. Properties of AdeABC and AdeIJK efflux systems of Acinetobacter baumannii compared with those of the AcrAB-TolC system of Escherichia coli. Antimicrob. Agents Chemother. 2014, 58, 7250–7257. [Google Scholar] [CrossRef] [Green Version]
- Parte, A.C.; Sarda Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Goker, M. List of prokaryotic names with standing in nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020. [Google Scholar] [CrossRef]
- Carvalheira, A.; Gonzales-Siles, L.; Salva-Serra, F.; Lindgren, A.; Svensson-Stadler, L.; Thorell, K.; Pineiro-Iglesias, B.; Karlsson, R.; Silva, J.; Teixeira, P.; et al. Acinetobacter portensis sp. nov. and Acinetobacter guerrae sp. nov., isolated from raw meat. Int. J. Syst. Evol. Microbiol. 2020, 70, 4544–4554. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020. [Google Scholar] [CrossRef] [PubMed]
- Murtagh, F.; Legendre, P. Ward’s hierarchical agglomerative clustering method: Which algorithms implement Ward’s criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, T.L. Hierarchical sets: Analyzing pangenome structure through scalable set visualizations. Bioinformatics 2017, 33, 1604–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Number of ARG in Acinetobacter non-baumannii Isolates Reported in the Literature | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β-lactamases | Aminoglycosides | Efflux Pump Genes | Sulfonamides | Tetracyclines | Macrolides | ABC-F | Rifampicin | Florfenicol | Bleomycin | Chloramphenicol | Carbapenem (CarO Specific) | OMP | Fluoroquinolone | PBPs | Pmr | Trimhoprim |
| A. baylys (n = 1) | A. baylys (n = 1) | A. apis (n = 1) | A. colistiniresistens (n = 1) | A. indicus (n = 1) | A. colistiniresistens (n = 1) | A. colistiniresistens (n = 1) | A. cumulans (n = 1) | A. haemolyticus (n = 1) | A. cululans (n = 1) | A. baylyi (n = 1) | A. apis (n = 1) | A. apis (n = 1) | A. baylyi (n = 1) | A. baylyi (n = 1) | A. nosocomialis (n = 1) | A. nosocomialis (n = 1) |
| A. bereziniae (n = 1) | A. calcoaceticus (n = 1) | A. baylys (n = 6) | A. cumulans (n = 1) | A. lwoffii (n = 1) | A. cumulans (n = 1) | A. cumulans (n = 1) | A. haemolyticus (n = 1) | A. johnsonii (n = 1) | A. johnsonii (n = 1) | A. johnsonii (n = 1) | A. nosocomialis (n = 1) | A. indicus (n = 1) | A. ursingii (n = 1) | A. indicus (n = 1) | A. seifertii (n = 1) | A. johnsonii (n = 1) |
| A. beijerinckii (n = 1) | A. colistiniresistens (n = 3) | A. cumulans (n = 1) | A. gandensis (n = 1) | A. nosocomialis (n = 2) | A. haemolyticus (n = 1) | A. haemolyticus (n = 1) | A. johnsonii (n = 1) | A. seifertii (n = 1) | A. pittii (n = 1) | A. modestus (n = 1) | ||||||
| A. bohemicus (n = 1) | A. cumulans (n = 3) | A. haemolyticus (n = 1) | A. haemolyticus (n = 2) | A. oleivorans (n = 3) | A. indicus (n = 1) | A. johnsonii (n = 1) | A. lwoffii (n = 1) | A. towneri (n = 1) | A. soli (n = 2) | A. pittii (n = 1) | ||||||
| A. calcoaceticus (n = 3) | A. gerneri (n = 2) | A. johnsonii (n = 1) | A. johnsonii (n = 2) | A. pitii (n = 1) | A. johnsonii (n = 2) | A. seifertii (n = 1) | A. seifertii (n = 1) | A. ursingii (n = 1) | ||||||||
| A. chinensis (n = 1) | A. guillouiae (n = 1) | A. lactucae (n = 4) | A. lwoffii (n = 1) | A. radioresistens (n = 2) | A. oleivorans (n = 1) | A. towneri (n = 1) | A. towneri (n = 1) | |||||||||
| A. colistiniresistens (n = 3) | A. gyllenbergii (n = 3) | A. lwoffii (n = 3) | A. pittii (n = 1) | A. seifertii (n = 2) | A. seifertii (n = 1) | A. ursingii (n = 1) | A. ursingii (n = 1) | |||||||||
| A. cumulans (n = 1) | A. haemolyticus (n = 7) | A. nosocomialis (n = 8) | A. radioresistens (n = 1) | A. towneri (n = 1) | A. towneri (n = 1) | |||||||||||
| A. dijkshoorniae (n = 3) | A. johnsonii (n = 9) | A. oleivorans (n = 3) | A. seifertii (n = 2) | A. ursingii (n = 1) | A. ursingii (n = 1) | |||||||||||
| A. disperusus (n = 1 | A. lwoffii (n = 5) | A. pittii (n = 5) | A. towneri (n = 2) | |||||||||||||
| A. gandensis (n = 2) | A. nosocomialis (n = 7) | A. seifertii (n = 2) | A. ursingii (n = 2) | |||||||||||||
| A. guillouiae (n = 6) | A. oleivorans (n = 1) | A. ursingii (n = 3) | ||||||||||||||
| A. gyllenbergii (n = 1) | A. parvus (n = 1) | |||||||||||||||
| A. haemolyticus (n = 14) | A. pittii (n = 9) | |||||||||||||||
| A. indicus (n = 3) | A. radioresistens (n = 6) | |||||||||||||||
| A. johnsonii (n = 13) | A. rudis (n = 1) | |||||||||||||||
| A. kyonggiensis (n = 1) | A. seifertii (n = 6) | |||||||||||||||
| A. lwoffii (n = 9) | A. soli (n = 1) | |||||||||||||||
| A. nosocomialis (n = 15) | A. ursingii (n = 5) | |||||||||||||||
| A. oleivorans (n = 2) | A. towneri (n = 5) | |||||||||||||||
| A. parvus (n = 1) | ||||||||||||||||
| A. pitti (n = 28) | ||||||||||||||||
| A. proteolyticus (n = 1) | ||||||||||||||||
| A. radioresistens (n = 7) | ||||||||||||||||
| A. schindleri (n = 6) | ||||||||||||||||
| A. seifertii (n = 5) | ||||||||||||||||
| A. soli (n = 4) | ||||||||||||||||
| A. tandoii (n = 1) | ||||||||||||||||
| A. towneri (n = 8) | ||||||||||||||||
| A. ursingii (n = 8) | ||||||||||||||||
| A. variabilis (n = 2) | ||||||||||||||||
| Total Number of ARGs | ||||||||||||||||
| N = 153 | N = 77 | N = 38 | N = 16 | N = 14 | N = 10 | N = 7 | N = 7 | N = 5 | N = 4 | N = 4 | N = 2 | N = 2 | N = 2 | N = 2 | N = 2 | N = 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations All authors have read and agreed to the published version of the manuscript.. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baraka, A.; Traglia, G.M.; Montaña, S.; Tolmasky, M.E.; Ramirez, M.S. An Acinetobacter non-baumannii Population Study: Antimicrobial Resistance Genes (ARGs). Antibiotics 2021, 10, 16. https://doi.org/10.3390/antibiotics10010016
Baraka A, Traglia GM, Montaña S, Tolmasky ME, Ramirez MS. An Acinetobacter non-baumannii Population Study: Antimicrobial Resistance Genes (ARGs). Antibiotics. 2021; 10(1):16. https://doi.org/10.3390/antibiotics10010016
Chicago/Turabian StyleBaraka, Adam, German M. Traglia, Sabrina Montaña, Marcelo E. Tolmasky, and Maria Soledad Ramirez. 2021. "An Acinetobacter non-baumannii Population Study: Antimicrobial Resistance Genes (ARGs)" Antibiotics 10, no. 1: 16. https://doi.org/10.3390/antibiotics10010016
APA StyleBaraka, A., Traglia, G. M., Montaña, S., Tolmasky, M. E., & Ramirez, M. S. (2021). An Acinetobacter non-baumannii Population Study: Antimicrobial Resistance Genes (ARGs). Antibiotics, 10(1), 16. https://doi.org/10.3390/antibiotics10010016