
Distribution and Clonal Diversity of Staphylococcus aureus and Other Staphylococci in Surface Waters: Detection of ST425-t742 and ST130-t843 mecC-Positive MRSA Strains

 ,
, 
 , ,
, ,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Distribution of Staphylococci in Surface Waters
2.2. Characterization of CoPS
2.3. Characterization of CoNS
3. Discussion
4. Materials and Methods
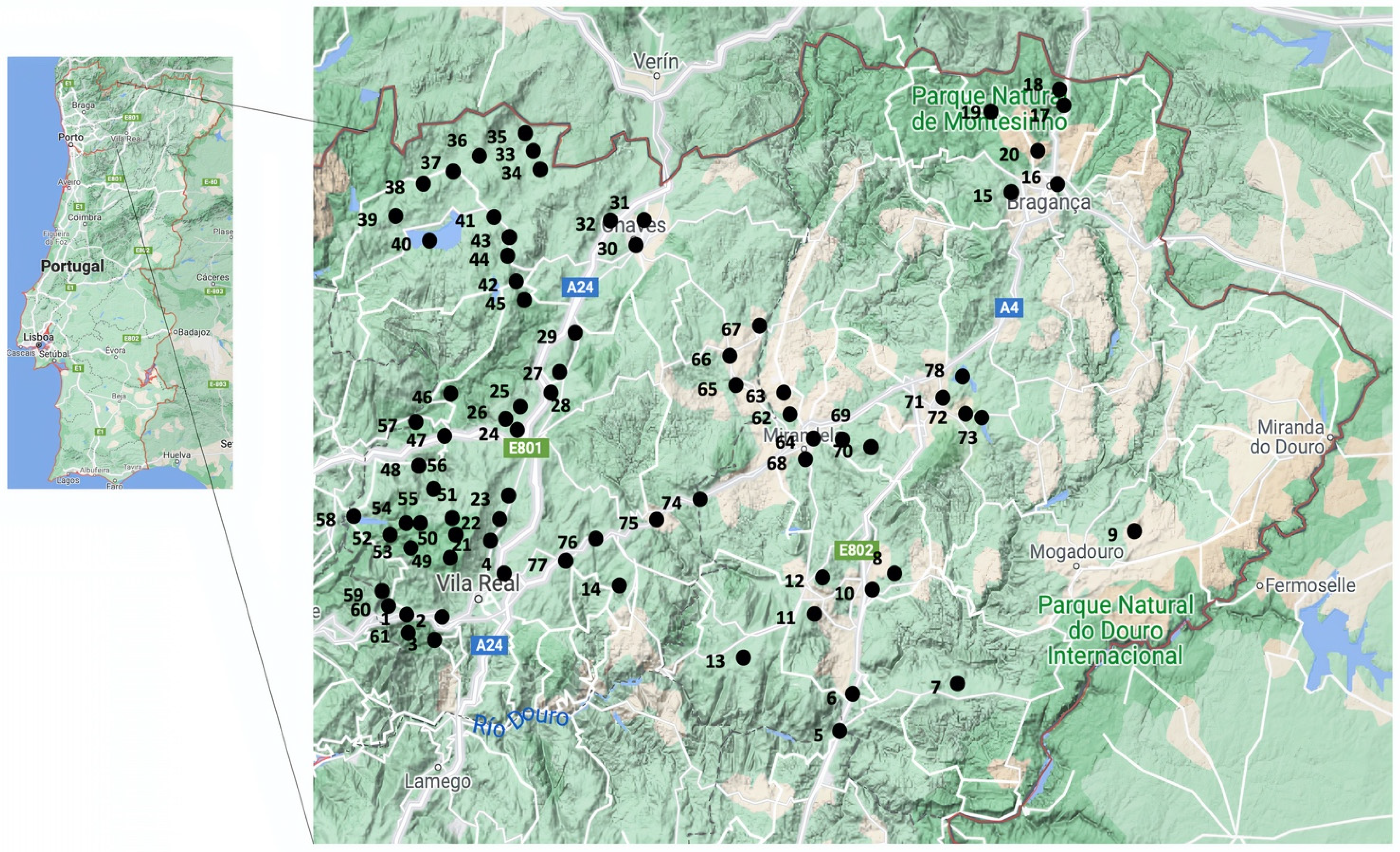
4.1. Study Area
4.2. Sample Collection
4.3. Bacterial Isolation
4.4. Antimicrobial Susceptibility Testing
4.5. Antimicrobial Resistance Genes and Virulence Factors
4.6. Molecular Typing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- O’Neill, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations: December 2014; Review on Antimicrobial Resistance. 2014. Available online: https://wellcomecollection.org/works/rdpck35v (accessed on 31 August 2021).
- Swift, B.M.C.; Bennett, M.; Waller, K.; Dodd, C.; Murray, A.; Gomes, R.L.; Humphreys, B.; Hobman, J.L.; Jones, M.A.; Whitlock, S.E.; et al. Anthropogenic environmental drivers of antimicrobial resistance in wildlife. Sci. Total Environ. 2019, 649, 12–20. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolejska, M.; Literak, I. Wildlife Is Overlooked in the Epidemiology of Medically Important Antibiotic-Resistant Bacteria. Antimicrob. Agents Chemother. 2021, 63, e01167-19. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Song, W.; Lin, H.; Wang, W.; Du, L.; Xing, W. Antibiotics and antibiotic resistance genes in global lakes: A review and meta-analysis. Environ. Int. 2018, 116, 60–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.A.; Ahmed, S.; Farooqi, I.H.; Ali, I.; Vambol, V.; Changani, F.; Yousefi, M.; Vambol, S.; Khan, S.U.; Khan, A.H. Occurrence, sources and conventional treatment techniques for various antibiotics present in hospital wastewaters: A critical review. TrAC Trends Anal. Chem. 2020, 129, 115921. [Google Scholar] [CrossRef]
- Senta, I.; Krizman-Matasic, I.; Terzic, S.; Ahel, M. Comprehensive determination of macrolide antibiotics, their synthesis intermediates and transformation products in wastewater effluents and ambient waters by liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2017, 1509, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, Z.; Xing, S.; Liao, X. The correlation between antibiotic resistance gene abundance and microbial community resistance in pig farm wastewater and surrounding rivers. Ecotoxicol. Environ. Saf. 2019, 182, 109452. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, C.; Cao, X.; Lin, H.; Wang, J. Antibiotic resistance genes in surface water of eutrophic urban lakes are related to heavy metals, antibiotics, lake morphology and anthropic impact. Ecotoxicology 2017, 26, 831–840. [Google Scholar] [CrossRef]
- Sabri, N.A.; Schmitt, H.; Van der Zaan, B.; Gerritsen, H.W.; Zuidema, T.; Rijnaarts, H.H.M.; Langenhoff, A.A.M. Prevalence of antibiotics and antibiotic resistance genes in a wastewater effluent-receiving river in the Netherlands. J. Environ. Chem. Eng. 2020, 8, 102245. [Google Scholar] [CrossRef]
- Garner, E.; Benitez, R.; von Wagoner, E.; Sawyer, R.; Schaberg, E.; Hession, W.C.; Krometis, L.-A.H.; Badgley, B.D.; Pruden, A. Stormwater loadings of antibiotic resistance genes in an urban stream. Water Res. 2017, 123, 144–152. [Google Scholar] [CrossRef]
- Savin, M.; Bierbaum, G.; Hammerl, J.A.; Heinemann, C.; Parcina, M.; Sib, E.; Voigt, A.; Kreyenschmidt, J. Antibiotic-resistant bacteria and antimicrobial residues in wastewater and process water from German pig slaughterhouses and their receiving municipal wastewater treatment plants. Sci. Total Environ. 2020, 727, 138788. [Google Scholar] [CrossRef]
- Finley, R.L.; Collignon, P.; Larsson, D.G.J.; McEwen, S.A.; Li, X.-Z.; Gaze, W.H.; Reid-Smith, R.; Timinouni, M.; Graham, D.W.; Topp, E. The Scourge of Antibiotic Resistance: The Important Role of the Environment. Clin. Infect. Dis. 2013, 57, 704–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thapaliya, D.; Hellwig, E.J.; Kadariya, J.; Grenier, D.; Jefferson, A.J.; Dalman, M.; Kennedy, K.; DiPerna, M.; Orihill, A.; Taha, M.; et al. Prevalence and Characterization of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus on Public Recreational Beaches in Northeast Ohio. GeoHealth 2017, 1, 320–332. [Google Scholar] [CrossRef] [Green Version]
- Zieliński, W.; Korzeniewska, E.; Harnisz, M.; Hubeny, J.; Buta, M.; Rolbiecki, D. The prevalence of drug-resistant and virulent Staphylococcus spp. in a municipal wastewater treatment plant and their spread in the environment. Environ. Int. 2020, 143, 105914. [Google Scholar] [CrossRef] [PubMed]
- Gerken, T.J.; Roberts, M.C.; Dykema, P.; Melly, G.; Lucas, D.; De Los Santos, V.; Gonzalez, J.; Butaye, P.; Wiegner, T.N. Environmental Surveillance and Characterization of Antibiotic Resistant Staphylococcus aureus at Coastal Beaches and Rivers on the Island of Hawai’i. Antibiotics 2021, 10, 980. [Google Scholar] [CrossRef]
- Silva, V.; Caniça, M.; Capelo, J.L.; Igrejas, G.; Poeta, P. Diversity and genetic lineages of environmental staphylococci: A surface water overview. FEMS Microbiol. Ecol. 2020, 96, fiaa191. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, G.; Di Bonaventura, G.; Savini, V. Staphylococcal Taxonomy; Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 9780128135488. [Google Scholar]
- Kim, E.; Kim, H.-J.; Yang, S.-M.; Kim, C.-G.; Choo, D.-W.; Kim, H.-Y. Rapid Identification of Staphylococcus Species Isolated from Food Samples by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. J. Microbiol. Biotechnol. 2019, 29, 548–557. [Google Scholar] [CrossRef] [PubMed]
- May, L.; Klein, E.Y.; Rothman, R.E.; Laxminarayan, R. Trends in antibiotic resistance in coagulase-negative staphylococci in the United States, 1999 to 2012. Antimicrob. Agents Chemother. 2014, 58, 1404–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, P.; Casado, C.; Sáenz, Y.; Ruiz-Ripa, L.; Estepa, V.; Zarazaga, M.; Torres, C. Diversity of species and antimicrobial resistance determinants of staphylococci in superficial waters in Spain. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, K.; van Alen, S.; Idelevich, E.A.; Schleimer, N.; Seggewiß, J.; Mellmann, A.; Kaspar, U.; Peters, G. Plasmid-Encoded Transferable mecB-Mediated Methicillin Resistance in Staphylococcus aureus. Emerg. Infect. Dis. 2018, 24, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shore, A.C.; Deasy, E.C.; Slickers, P.; Brennan, G.; O’Connell, B.; Monecke, S.; Ehricht, R.; Coleman, D.C. Detection of staphylococcal cassette chromosome mec type XI carrying highly divergent mecA, mecI, mecR1, blaZ, and ccr genes in human clinical isolates of clonal complex 130 methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 3765–3773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, V.; Capelo, J.L.; Igrejas, G.; Poeta, P. Molecular Epidemiology of Staphylococcus aureus Lineages in Wild Animals in Europe: A Review. Antibiotics 2020, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- MacFadyen, A.C.; Harrison, E.M.; Ellington, M.J.; Parkhill, J.; Holmes, M.A.; Paterson, G.K. A highly conserved mecC-encoding SCCmec type XI in a bovine isolate of methicillin-resistant Staphylococcus xylosus. J. Antimicrob. Chemother. 2018, 73, 3516–3518. [Google Scholar] [CrossRef] [PubMed]
- Porrero, M.C.; Harrison, E.; Fernández-Garayzábal, J.F.; Paterson, G.K.; Díez-Guerrier, A.; Holmes, M.A.; Domínguez, L. Detection of mecC-Methicillin-resistant Staphylococcus aureus isolates in river water: A potential role for water in the environmental dissemination. Environ. Microbiol. Rep. 2014, 6, 705–708. [Google Scholar] [CrossRef]
- Lepuschitz, S.; Mach, R.; Springer, B.; Allerberger, F.; Ruppitsch, W. Draft Genome Sequence of a Community-Acquired Methicillin-Resistant Staphylococcus aureus USA300 Isolate from a River Sample. Genome Announc. 2017, 5, e01166-17. [Google Scholar] [CrossRef] [Green Version]
- Heß, S.; Gallert, C. Demonstration of staphylococci with inducible macrolide–lincosamide–streptogramin B (MLSB) resistance in sewage and river water and of the capacity of anhydroerythromycin to induce MLSB. FEMS Microbiol. Ecol. 2014, 88, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Ramessar, K.; Olaniran, A.O. Antibiogram and molecular characterization of methicillin-resistant Staphylococcus aureus recovered from treated wastewater effluent and receiving surface water in Durban, South Africa. World J. Microbiol. Biotechnol. 2019, 35, 142. [Google Scholar] [CrossRef]
- Hatcher, S.M.; Myers, K.W.; Heaney, C.D.; Larsen, J.; Hall, D.; Miller, M.B.; Stewart, J.R. Occurrence of methicillin-resistant Staphylococcus aureus in surface waters near industrial hog operation spray fields. Sci. Total Environ. 2016, 565, 1028–1036. [Google Scholar] [CrossRef] [Green Version]
- Sood, A.; Pandey, P.; Bisht, S.; Sharma, S. Anthropogenic activities as a source of high prevalence of antibiotic resistant Staphylococcus aureus in the River Ganga. Appl. Ecol. Env. Res. 2014, 12, 33–48. [Google Scholar] [CrossRef]
- Cho, S.; Jackson, C.R.; Frye, J.G. The prevalence and antimicrobial resistance phenotypes of Salmonella, Escherichia coli and Enterococcus sp. in surface water. Lett. Appl. Microbiol. 2020, 71, 3–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, P.; González-Barrio, D.; Benito, D.; García, J.T.; Viñuela, J.; Zarazaga, M.; Ruiz-Fons, F.; Torres, C. Detection of methicillin-resistant Staphylococcus aureus (MRSA) carrying the mecC gene in wild small mammals in Spain. J. Antimicrob. Chemother. 2014, 69, 2061–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin-Edens, E.; Soge, O.O.; No, D.; Stiffarm, A.; Meschke, J.S.; Roberts, M.C. Methicillin-resistant Staphylococcus aureus from Northwest marine and freshwater recreational beaches. FEMS Microbiol. Ecol. 2012, 79, 412–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akanbi, O.E.; Njom, H.A.; Fri, J.; Otigbu, A.C.; Clarke, A.M. Antimicrobial susceptibility of Staphylococcus aureus isolated from recreational waters and beach sand in Eastern Cape Province of South Africa. Int. J. Environ. Res. Public Health 2017, 14, 1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, J.E.; Moore, R.E.; Shteinberg, M.; Kis-Papo, T.; Millar, B.C. Survival of Mycobacterium abscessus and Staphylococcus aureus in saline waters of the Dead Sea: Implications for health tourists. J. Travel Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Faria, C.; Vaz-Moreira, I.; Serapicos, E.; Nunes, O.C.; Manaia, C.M. Antibiotic resistance in coagulase negative staphylococci isolated from wastewater and drinking water. Sci. Total Environ. 2009, 407, 3876–3882. [Google Scholar] [CrossRef]
- Rey Pérez, J.; Zálama Rosa, L.; García Sánchez, A.; Hermoso de Mendoza Salcedo, J.; Alonso Rodríguez, J.M.; Cerrato Horrillo, R.; Zurita, S.G.; Gil Molino, M. Multiple Antimicrobial Resistance in Methicillin-Resistant Staphylococcus sciuri Group Isolates from Wild Ungulates in Spain. Antibiotics 2021, 10, 920. [Google Scholar] [CrossRef]
- Silva, V.; Gabriel, S.I.; Borrego, S.B.; Tejedor-Junco, M.T.; Manageiro, V.; Ferreira, E.; Reis, L.; Caniça, M.; Capelo, J.L.; Igrejas, G.; et al. Antimicrobial Resistance and Genetic Lineages of Staphylococcus aureus from Wild Rodents: First Report of mecC-Positive Methicillin-Resistant S. aureus (MRSA) in Portugal. Animals 2021, 11, 1537. [Google Scholar] [CrossRef]
- Monecke, S.; Gavier-Widén, D.; Hotzel, H.; Peters, M.; Guenther, S.; Lazaris, A.; Loncaric, I.; Müller, E.; Reissig, A.; Ruppelt-Lorz, A.; et al. Diversity of Staphylococcus aureus Isolates in European Wildlife. PLoS ONE 2016, 11, e0168433. [Google Scholar]
- Bengtsson, B.; Persson, L.; Ekström, K.; Unnerstad, H.E.; Uhlhorn, H.; Börjesson, S. High occurrence of mecC-MRSA in wild hedgehogs (Erinaceus europaeus) in Sweden. Vet. Microbiol. 2017, 207, 103–107. [Google Scholar] [CrossRef]
- García-Álvarez, L.; Holden, M.T.G.; Lindsay, H.; Webb, C.R.; Brown, D.F.J.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet. Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Giacinti, G.; Carfora, V.; Caprioli, A.; Sagrafoli, D.; Marri, N.; Giangolini, G.; Amoruso, R.; Iurescia, M.; Stravino, F.; Dottarelli, S.; et al. Prevalence and characterization of methicillin-resistant Staphylococcus aureus carrying mecA or mecC and methicillin-susceptible Staphylococcus aureus in dairy sheep farms in central Italy. J. Dairy Sci. 2017, 100, 7857–7863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porrero, M.C.; Valverde, A.; Fernández-Llario, P.; Díez-Guerrier, A.; Mateos, A.; Lavín, S.; Cantón, R.; Fernández-Garayzabal, J.-F.; Domínguez, L. Staphylococcus aureus carrying mecC gene in animals and urban wastewater, Spain. Emerg. Infect. Dis. 2014, 20, 899–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monecke, S.; Gavier-Widen, D.; Mattsson, R.; Rangstrup-Christensen, L.; Lazaris, A.; Coleman, D.C.; Shore, A.C.; Ehricht, R. Detection of mecC-Positive Staphylococcus aureus (CC130-MRSA-XI) in Diseased European Hedgehogs (Erinaceus europaeus) in Sweden. PLoS ONE 2013, 8, e66166. [Google Scholar]
- Kerschner, H.; Harrison, E.M.; Hartl, R.; Holmes, M.A.; Apfalter, P. First report of mecC MRSA in human samples from Austria: Molecular characteristics and clinical data. New Microbes New Infect. 2015, 3, 4–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worthing, K.A.; Coombs, G.W.; Pang, S.; Abraham, S.; Saputra, S.; Trott, D.J.; Jordan, D.; Wong, H.S.; Abraham, R.J.; Norris, J.M. Isolation of mecC MRSA in Australia. J. Antimicrob. Chemother. 2016, 71, 2348–2349. [Google Scholar] [CrossRef] [Green Version]
- Tavares, A.; Faria, N.A.; de Lencastre, H.; Miragaia, M. Population structure of methicillin-susceptible Staphylococcus aureus (MSSA) in Portugal over a 19-year period (1992–2011). Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 423–432. [Google Scholar] [CrossRef]
- Satta, G.; Ling, C.L.; Cunningham, E.S.; McHugh, T.D.; Hopkins, S. Utility and limitations of Spa-typing in understanding the epidemiology of staphylococcus aureus bacteraemia isolates in a single University Hospital. BMC Res. Notes 2013, 6, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, W.L.; Sohn, M.B.; Taffner, S.; Chatterjee, P.; Dunman, P.M.; Pecora, N.; Wozniak, R.A.F. Genomics of Staphylococcus aureus ocular isolates. PLoS ONE 2021, 16, e0250975. [Google Scholar] [CrossRef]
- David, M.Z.; Boyle-Vavra, S.; Zychowski, D.L.; Daum, R.S. Methicillin-Susceptible Staphylococcus aureus as a Predominantly Healthcare-Associated Pathogen: A Possible Reversal of Roles? PLoS ONE 2011, 6, e18217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plano, L.R.W.; Shibata, T.; Garza, A.C.; Kish, J.; Fleisher, J.M.; Sinigalliano, C.D.; Gidley, M.L.; Withum, K.; Elmir, S.M.; Hower, S.; et al. Human-Associated Methicillin-Resistant Staphylococcus aureus from a Subtropical Recreational Marine Beach. Microb. Ecol. 2013, 65, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Hofer, U. Squirrel-killing Staphylococcus aureus. Nat. Rev. Microbiol. 2021, 19, 481. [Google Scholar] [CrossRef] [PubMed]
- Fountain, K.; Blackett, T.; Butler, H.; Carchedi, C.; Schilling, A.-K.; Meredith, A.; Gibbon, M.J.; Lloyd, D.H.; Loeffler, A.; Feil, E.J. Fatal exudative dermatitis in island populations of red squirrels (Sciurus vulgaris): Spillover of a virulent Staphylococcus aureus clone (ST49) from reservoir hosts. Microb. Genom. 2021, 7, 565. [Google Scholar] [CrossRef]
- Mrochen, D.M.; Schulz, D.; Fischer, S.; Jeske, K.; El Gohary, H.; Reil, D.; Imholt, C.; Trübe, P.; Suchomel, J.; Tricaud, E.; et al. Wild rodents and shrews are natural hosts of Staphylococcus aureus. Int. J. Med. Microbiol. 2018, 308, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Deplano, A.; Vandendriessche, S.; Nonhoff, C.; Denis, O. Genetic diversity among methicillin-resistant Staphylococcus aureus isolates carrying the mecC gene in Belgium. J. Antimicrob. Chemother. 2014, 69, 1457–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haenni, M.; Châtre, P.; Dupieux, C.; Métayer, V.; Maillard, K.; Bes, M.; Madec, J.-Y.; Laurent, F. mecC-positive MRSA in horses. J. Antimicrob. Chemother. 2015, 70, 3401–3402. [Google Scholar] [CrossRef] [Green Version]
- Porrero, M.; Valverde, A.; Mateos, A.; Cantón, R.; Gortázar, C.; Fernández-Garayzábal, J.-F.; Domínguez, L. Staphylococcus aureus Genetic Lineages Found in Urban Effluents and River Water. Int. J. Water Wastewater Treat. 2016, 2, 1–5. [Google Scholar]
- Cuny, C.; Layer, F.; Hansen, S.; Werner, G.; Witte, W. Nasal Colonization of Humans with Occupational Exposure to Raw Meat and to Raw Meat Products with Methicillin-Susceptible and Methicillin-Resistant Staphylococcus aureus. Toxins (Basel) 2019, 11, 190. [Google Scholar] [CrossRef] [Green Version]
- Little, S.V.; Hillhouse, A.E.; Lawhon, S.D.; Bryan, L.K. Analysis of Virulence and Antimicrobial Resistance Gene Carriage in Staphylococcus aureus Infections in Equids Using Whole-Genome Sequencing. mSphere 2021, 6, e00196-20. [Google Scholar] [CrossRef]
- Mama, O.M.; Morales, L.; Ruiz-Ripa, L.; Zarazaga, M.; Torres, C. High prevalence of multidrug resistant S. aureus-CC398 and frequent detection of enterotoxin genes among non-CC398 S. aureus from pig-derived food in Spain. Int. J. Food Microbiol. 2020, 320, 108510. [Google Scholar] [CrossRef]
- Tolba, O.; Loughrey, A.; Goldsmith, C.E.; Millar, B.C.; Rooney, P.J.; Moore, J.E. Survival of epidemic strains of healthcare (HA-MRSA) and community-associated (CA-MRSA) meticillin-resistant Staphylococcus aureus (MRSA) in river-, sea- and swimming pool water. Int. J. Hyg. Environ. Health 2008, 211, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Umeda, K.; Nakamura, H.; Yamamoto, K.; Nishina, N.; Yasufuku, K.; Hirai, Y.; Hirayama, T.; Goto, K.; Hase, A.; Ogasawara, J. Molecular and epidemiological characterization of staphylococcal foodborne outbreak of Staphylococcus aureus harboring seg, sei, sem, sen, seo, and selu genes without production of classical enterotoxins. Int. J. Food Microbiol. 2017, 256, 30–35. [Google Scholar] [CrossRef]
- Rossi, B.F.; Bonsaglia, E.C.R.; Castilho, I.G.; Dantas, S.T.A.; Salina, A.; Langoni, H.; Pantoja, J.C.F.; Budri, P.E.; Fitzgerald-Hughes, D.; Júnior, A.F.; et al. Genotyping of long term persistent Staphylococcus aureus in bovine subclinical mastitis. Microb. Pathog. 2019, 132, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Lahuerta-Marin, A.; Guelbenzu-Gonzalo, M.; Pichon, B.; Allen, A.; Doumith, M.; Lavery, J.F.; Watson, C.; Teale, C.J.; Kearns, A.M. First report of lukM-positive livestock-associated methicillin-resistant Staphylococcus aureus CC30 from fattening pigs in Northern Ireland. Vet. Microbiol. 2016, 182, 131–134. [Google Scholar] [CrossRef]
- Pérez-Montarelo, D.; Viedma, E.; Larrosa, N.; Gómez-González, C.; Ruiz de Gopegui, E.; Muñoz-Gallego, I.; San Juan, R.; Fernández-Hidalgo, N.; Almirante, B.; Chaves, F. Molecular Epidemiology of Staphylococcus aureus Bacteremia: Association of Molecular Factors With the Source of Infection. Front. Microbiol. 2018, 9, 2210. [Google Scholar] [CrossRef]
- Sousa, M.; Silva, N.; Manageiro, V.; Ramos, S.; Coelho, A.; Gonçalves, D.; Caniça, M.; Torres, C.; Igrejas, G.; Poeta, P. First report on MRSA CC398 recovered from wild boars in the north of Portugal. Are we facing a problem? Sci. Total Environ. 2017, 596–597, 26–31. [Google Scholar] [CrossRef]
- Porrero, M.C.; Mentaberre, G.; Sánchez, S.; Fernández-Llario, P.; Gómez-Barrero, S.; Navarro-Gonzalez, N.; Serrano, E.; Casas-Díaz, E.; Marco, I.; Fernández-Garayzabal, J.F.; et al. Methicillin resistant Staphylococcus aureus (MRSA) carriage in different free-living wild animal species in Spain. Vet. J. 2013, 198, 127–130. [Google Scholar] [CrossRef]
- Shuaibu, S.A.; Onaolapo, J.A.; Olayinka, B.O. Towards Understanding the Clonality of Methicillin Resistant Staphylococcus aureus in Sokoto State Nigeria. Rep. Heal. Care 2019, 5, 44–53. [Google Scholar]
- Aggarwal, S.; Jena, S.; Panda, S.; Sharma, S.; Dhawan, B.; Nath, G.; Singh, N.P.; Nayak, K.C.; Singh, D.V. Antibiotic Susceptibility, Virulence Pattern, and Typing of Staphylococcus aureus Strains Isolated From Variety of Infections in India. Front. Microbiol. 2019, 10, 2763. [Google Scholar] [CrossRef]
- Overesch, G.; Büttner, S.; Rossano, A.; Perreten, V. The increase of methicillin-resistant Staphylococcus aureus(MRSA) and the presence of an unusual sequence type ST49 in slaughter pigs in Switzerland. BMC Vet. Res. 2011, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, V.R.; Davison, N.J.; Kearns, A.M.; Pichon, B.; Hudson, L.O.; Koylass, M.; Blackett, T.; Butler, H.; Rasigade, J.P.; Whatmore, A.M. Association of a lukM-positive clone of Staphylococcus aureus with fatal exudative dermatitis in red squirrels (Sciurus vulgaris). Vet. Microbiol. 2013, 162, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Seyedmonir, E.; Yilmaz, F.; Icgen, B. mecA Gene Dissemination Among Staphylococcal and Non-staphylococcal Isolates Shed in Surface Waters. Bull. Environ. Contam. Toxicol. 2015, 95, 131–138. [Google Scholar] [CrossRef]
- Čuvalová, Z.; Pipová, M.; Kantíková, M.; Brtková, A.; Fajber, J. Virulence factors and antimicrobial resistance of coagulase-negative staphylococci isolated from drinking water. Open Life Sci. 2015, 10, 328–338. [Google Scholar] [CrossRef]
- Yilmaz, F.; Orman, N.; Serim, G.; Kochan, C.; Ergene, A.; Icgen, B. Surface Water-Borne Multidrug and Heavy Metal-Resistant Staphylococcus Isolates Characterized by 16S rDNA Sequencing. Bull. Environ. Contam. Toxicol. 2013, 91, 697–703. [Google Scholar] [CrossRef]
- Silva, V.; Almeida, F.; Carvalho, J.A.; Castro, A.P.; Ferreira, E.; Manageiro, V.; Tejedor-Junco, M.T.; Caniça, M.; Igrejas, G.; Poeta, P. Emergence of community-acquired methicillin-resistant Staphylococcus aureus EMRSA-15 clone as the predominant cause of diabetic foot ulcer infections in Portugal. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 179–186. [Google Scholar] [CrossRef]
- Cuny, C.; Layer, F.; Strommenger, B.; Witte, W. Rare occurrence of methicillin-resistant Staphylococcus aureus CC130 with a novel mecA homologue in humans in Germany. PLoS ONE 2011, 6, e24360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Sparling, J.; Chow, B.L.; Elsayed, S.; Hussain, Z.; Church, D.L.; Gregson, D.B.; Louie, T.; Conly, J.M. New quadriplex PCR assay for detection of methicillin and mupirocin resistance and simultaneous discrimination of Staphylococcus aureus from coagulase-negative staphylococci. J. Clin. Microbiol. 2004, 42, 4947–4955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, V.; Vieira-Pinto, M.; Saraiva, C.; Manageiro, V.; Reis, L.; Ferreira, E.; Caniça, M.; Capelo, J.L.; Igrejas, G.; Poeta, P. Prevalence and Characteristics of Multidrug-Resistant Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) CC398 Isolated from Quails (Coturnix Coturnix Japonica) Slaughtered for Human Consumption. Animals 2021, 11, 2038. [Google Scholar] [CrossRef]
- van Wamel, W.J.B.; Rooijakkers, S.H.M.; Ruyken, M.; van Kessel, K.P.M.; van Strijp, J.A.G. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta-hemolysin-converting bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Harmsen, D.; Claus, H.H.H.H.; Witte, W.; Rothgänger, J.; Claus, H.H.H.H.; Turnwald, D.; Vogel, U. Typing of Methicillin-Resistant Staphylococcus aureus in a University Hospital Setting by Using Novel Software for spa Repeat Determination and Database Management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shopsin, B.; Mathema, B.; Alcabes, P.; Said-Salim, B.; Lina, G.; Matsuka, A.; Martinez, J.; Kreiswirth, B.N. Prevalence of agr specificity groups among Staphylococcus aureus strains colonizing children and their guardians. J. Clin. Microbiol. 2003, 41, 456–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Source | Number of Samples | Number of CoPS | Number of CoNS |
|---|---|---|---|
| Rivers | 29 | 11 | 22 |
| Streams | 19 | 6 | 14 |
| Irrigation ditches | 12 | 8 | 8 |
| Dams | 10 | 5 | 4 |
| Fountains | 4 | 1 | 1 |
| Lakes | 4 | 3 | 2 |
| Total | 78 | 34 | 51 |
| Isolate | Water Source | Antimicrobial Resistance | Virulence Factors | Molecular Typing | ||||
|---|---|---|---|---|---|---|---|---|
| Phenotype | Genotype | IEC System | Other Genes | ST (CC) | spa | agr | ||
| VS2847 | River | PEN, FOX | blaZ-SCCmecIX | - | hld | 425 | t742 | III |
| VS2848 | River | PEN, FOX | blaZ-SCCmecIX | - | hld | 425 | t742 | III |
| VS2846 | Irrigation ditch | PEN, FOX | blaZ-SCCmecIX | - | hld | 130 | t843 | III |
| VS2849 | River | PEN, FOX | blaZ-SCCmecIX | - | hld | 425 | t742 | III |
| VS2850 | Dam | PEN | blaZ | A | hla, hlb, hld | 30 (30) | t9413 | III |
| VS2851 | Lake | PEN | blaZ | A | hla, hlb, hld | 8 (8) | t008 | I |
| VS2852 | Lake | PEN | blaZ | D | hla, hlb, hld | 8 (8) | t008 | I |
| VS2853 | Dam | PEN | blaZ | D | hla, hlb, hld | 8 (8) | t008 | I |
| VS2854 | River | PEN | blaZ | - | hla, hlb, hld | 8 (8) | t008 | I |
| VS2855 | River | PEN | blaZ | D | hla, hlb, hld | 8 (8) | t008 | I |
| VS2856 | Dam | PEN | blaZ | D | hla, hlb, hld | 8 (8) | t008 | I |
| VS2857 | Fountain | PEN | blaZ | - | hla, hlb, hld | 8 (8) | t008 | I |
| VS2858 | Dam | PEN | blaZ | B | hla, hld | 617 (45) | t350 | I |
| VS2859 | River | ERY | ermT | B | hla, hld | 398 | t571 | I |
| VS2860 | Stream | PEN, ERY | blaZ, ermT, msr(A/B) | C | hla, hld | 398 | t1451 | I |
| VS2861 | Irrigation ditch | Susceptible | - | - | hla, hlb, hld | 49 (49) | t208 | II |
| VS2862 | River | Susceptible | - | - | hla, hlb, hld | 49 (49) | t208 | II |
| VS2863 | Irrigation ditch | Susceptible | - | - | hla, hlb, hld | 49 (49) | t208 | n.d. |
| VS2864 | River | Susceptible | - | - | hla, hlb, hld | 3223 | t742 | n.d. |
| VS2865 | Irrigation ditch | Susceptible | - | - | hla, hlb, hld | 6832 (1) | t098 | III |
| VS2866 | Stream | PEN | - | - | hla, hlb, hld | 352 (97) | t267 | I |
| VS2867 | Irrigation ditch | Susceptible | - | - | hla, hlb, hld | 6833 | t098 | IV |
| VS2868 | Stream | Susceptible | - | - | hla, hlb, hld | 352 (97) | t267 | I |
| VS2869 | Irrigation ditch | Susceptible | - | - | hla, hlb | 6834 | t208 | II |
| VS2870 | Irrigation ditch | Susceptible | - | - | hla, hlb, hld | 6833 | t098 | IV |
| VS2871 | Stream | PEN, TET | blaZ, tetL | - | hla, hlb, hld, tst | 425 | t742 | II |
| VS2872 | River | Susceptible | - | - | hla, hlb, hld | 6835 | t098 | IV |
| VS2873 | River | ERY | vgaA | - | hla, hlb, hld | 133 | t4735 | I |
| VS2874 | River | Susceptible | - | - | - | 6836 | t8083 | n.d. |
| VS2875 | River | Susceptible | - | - | hla, hlb | 133 | t4735 | I |
| VS2876 | Stream | Susceptible | - | - | hla, | 130 | t1532 | III |
| VS2877 | Dam | PEN | blaZ | B | hla, hld, eta | 582 (15) | t1877 | II |
| VS2878 | Lake | Susceptible | - | - | - | 243 (30) | t021 | n.d. |
| Species | Number of Isolates | Antimicrobial Resistance | Virulence | |
|---|---|---|---|---|
| Phenotype | Genotype | |||
| S. sciuri | 28 | PEN 12, CD 9, FD 11 | MecA 27, ermT 2, msr(A/B) 2 | Hld2 |
| S. lentus | 5 | PEN 2, CD 2, TET 1 | MecA 5, msr(A/B) 1, mph(C) 2, tetK 1 | - |
| S. xylosus | 5 | PEN 1, CD 1, TET 1 | MecA 4, tetK 1 | - |
| S. epidermidis | 4 | PEN 3, FOX 2, CIP 1, LNZ 1, CN 2, TOB 2, KAN 2, ERY 1, CD 2, C 1, FD 1, SXT 1 | MecA 4, aac(6′)-Ie–aph(2′’)-Ia 1, aph(3’)-IIIa 2, msr(A/B) 3, mph(C) 2, catpC221 1, fusB 1, dfrA 1 | - |
| S. cohnii spp. urealyticus | 2 | PEN 2, ERY 1, CD 1, FD 1 | MecA 2, msr(A/B), mph(C) | - |
| S. vitulinus | 2 | Susceptible | mecA2 | - |
| S. caprae | 1 | FD | mecA | - |
| S. succinus | 1 | Susceptible | mecA | - |
| S. carnosus spp. carnosus | 1 | Susceptible | mecA | - |
| S. equorum | 1 | Susceptible | mecA | - |
| S. simulans | 1 | Susceptible | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, V.; Ferreira, E.; Manageiro, V.; Reis, L.; Tejedor-Junco, M.T.; Sampaio, A.; Capelo, J.L.; Caniça, M.; Igrejas, G.; Poeta, P. Distribution and Clonal Diversity of Staphylococcus aureus and Other Staphylococci in Surface Waters: Detection of ST425-t742 and ST130-t843 mecC-Positive MRSA Strains. Antibiotics 2021, 10, 1416. https://doi.org/10.3390/antibiotics10111416
Silva V, Ferreira E, Manageiro V, Reis L, Tejedor-Junco MT, Sampaio A, Capelo JL, Caniça M, Igrejas G, Poeta P. Distribution and Clonal Diversity of Staphylococcus aureus and Other Staphylococci in Surface Waters: Detection of ST425-t742 and ST130-t843 mecC-Positive MRSA Strains. Antibiotics. 2021; 10(11):1416. https://doi.org/10.3390/antibiotics10111416
Chicago/Turabian StyleSilva, Vanessa, Eugénia Ferreira, Vera Manageiro, Lígia Reis, María Teresa Tejedor-Junco, Ana Sampaio, José Luis Capelo, Manuela Caniça, Gilberto Igrejas, and Patrícia Poeta. 2021. "Distribution and Clonal Diversity of Staphylococcus aureus and Other Staphylococci in Surface Waters: Detection of ST425-t742 and ST130-t843 mecC-Positive MRSA Strains" Antibiotics 10, no. 11: 1416. https://doi.org/10.3390/antibiotics10111416
APA StyleSilva, V., Ferreira, E., Manageiro, V., Reis, L., Tejedor-Junco, M. T., Sampaio, A., Capelo, J. L., Caniça, M., Igrejas, G., & Poeta, P. (2021). Distribution and Clonal Diversity of Staphylococcus aureus and Other Staphylococci in Surface Waters: Detection of ST425-t742 and ST130-t843 mecC-Positive MRSA Strains. Antibiotics, 10(11), 1416. https://doi.org/10.3390/antibiotics10111416