
Effects of Essential Oils from Cymbopogon spp. and Cinnamomum verum on Biofilm and Virulence Properties of Escherichia coli O157:H7

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Components of Essential Oils
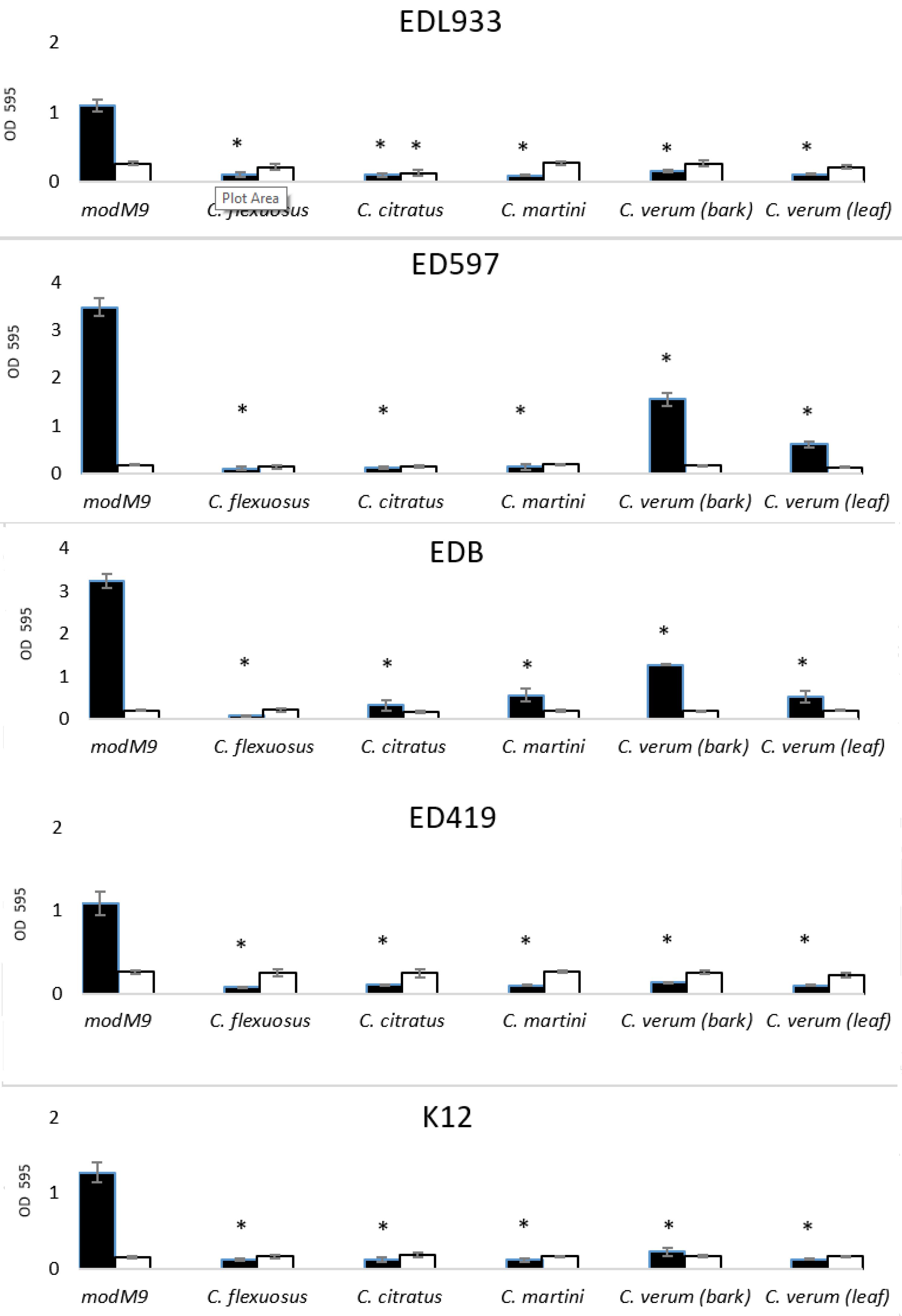
2.2. Antibacterial and Antibiofilm Activity of EOs
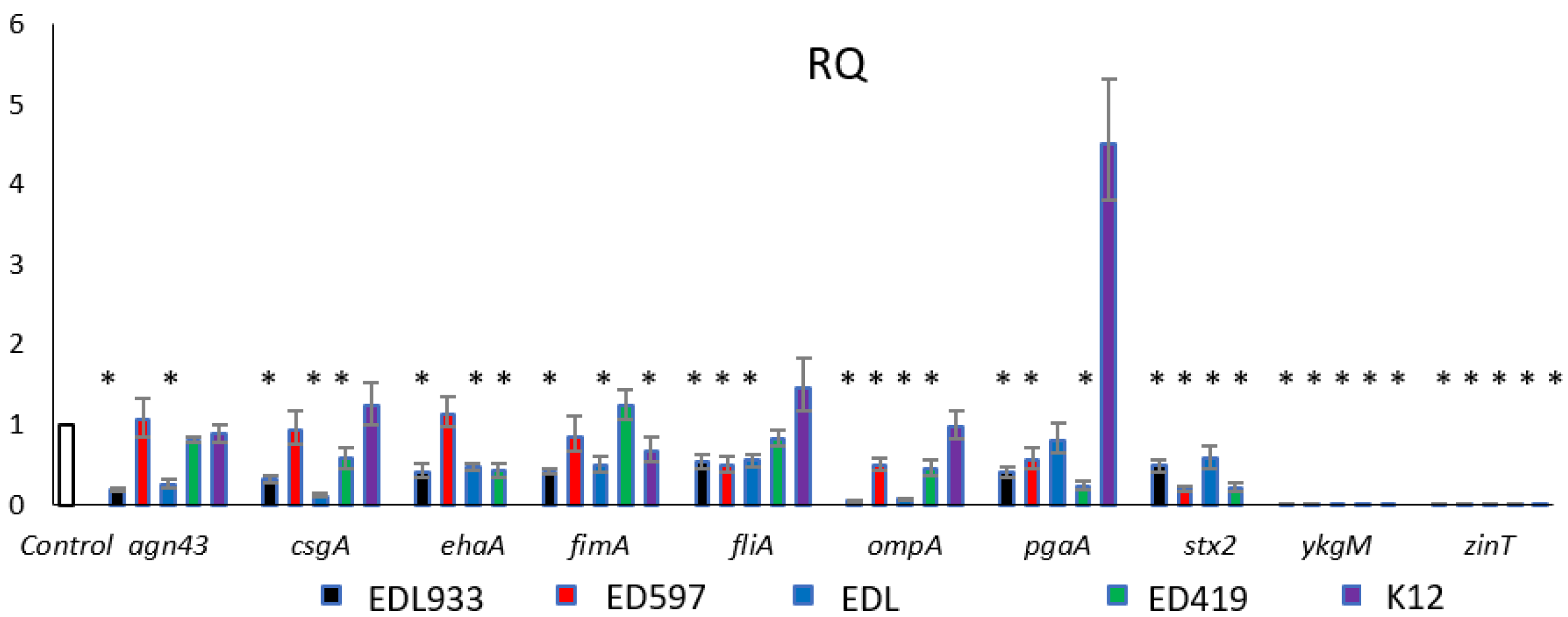
2.3. Gene Expression Analysis
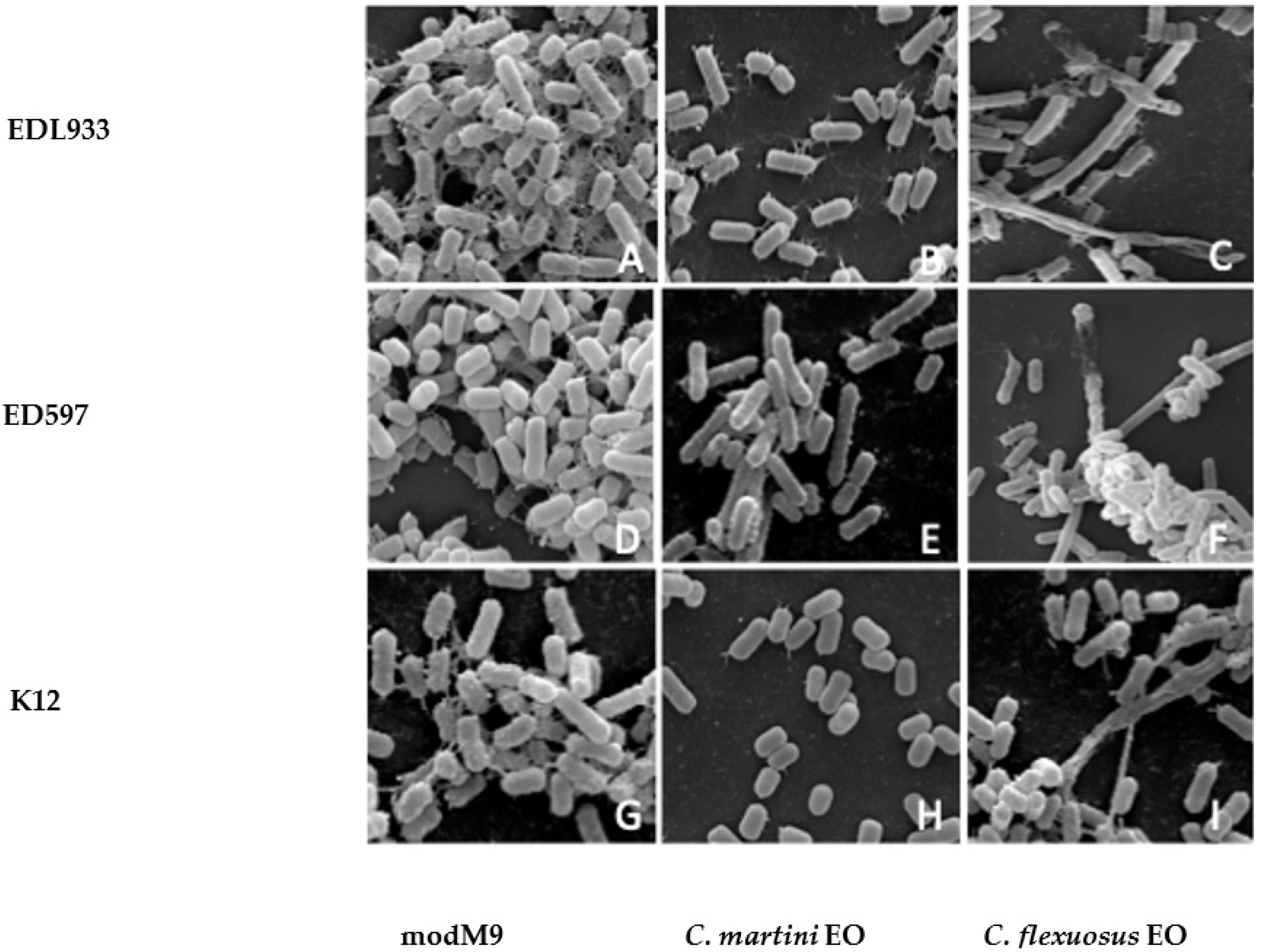
2.4. Scanning Electron Microscope Analysis
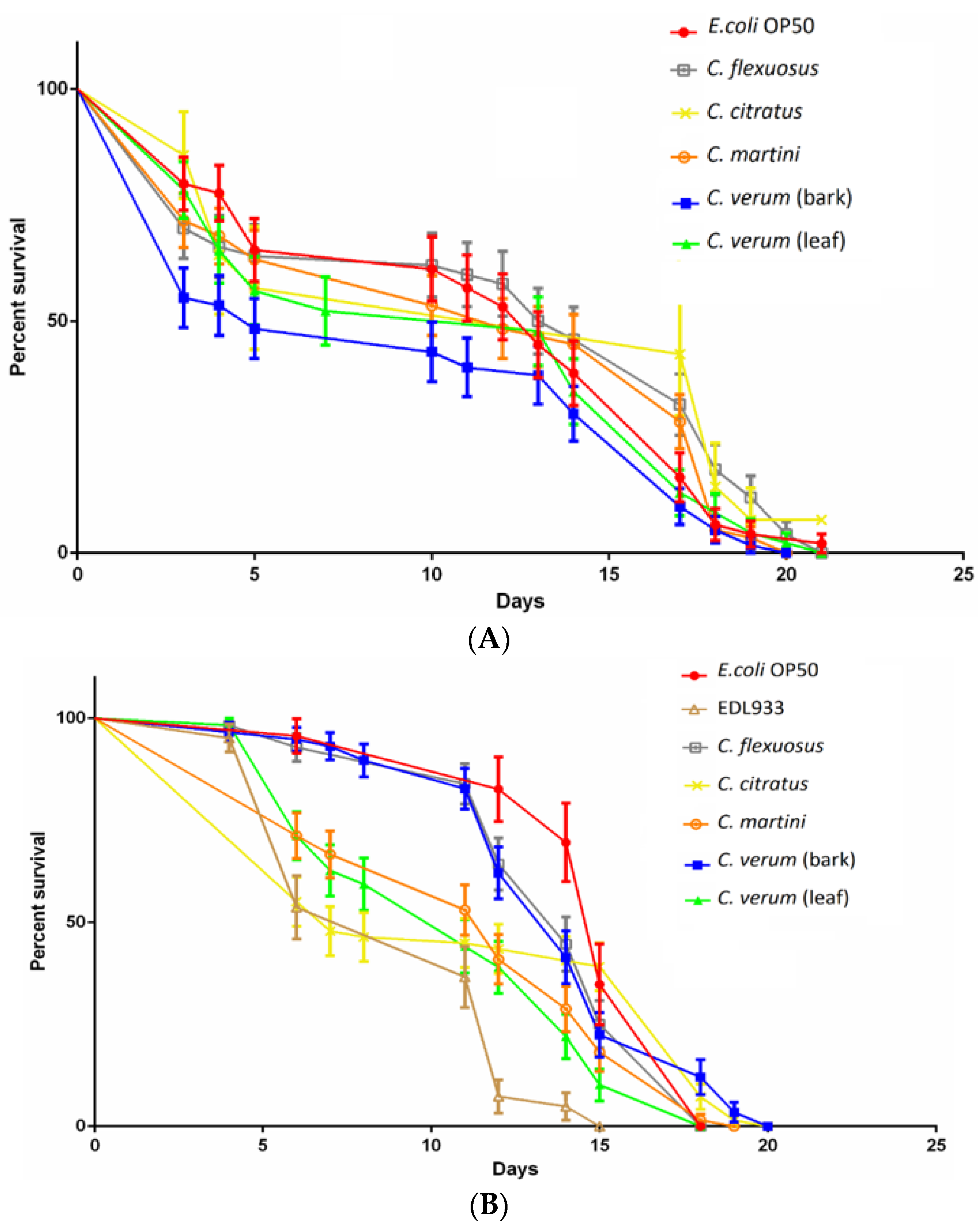
2.5. EOs Antibacterial Activity in the Nematode Model
3. Materials and Methods
3.1. Essential Oils
3.2. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
3.3. Bacterial Strains
3.4. MIC and MBC Assay
3.5. Biofilm Formation Assay
3.6. Scanning Electron Microscopy (SEM)
3.7. RNA Isolation and Quantitative Real-Time RT-PCR
3.8. Caenorhabditis Elegans Killing Assay
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elcock, E.R.; Spencer-Phillips, P.T.N.; Adukwu, E.C. Rapid bactericidal effect of cinnamon bark essential against Pseudomonas aeruginosa. J. Appl. Microbiol. 2019, 128, 1025–1037. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilm, an emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef] [PubMed]
- Paton, J.C.; Paton, A.W. Pathogenesis and diagnosis of Shiga toxin-producing Escherichia coli Infections. Clin. Microbiol. Rev. 1998, 11, 450–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, L.; Rasco, B.; Zhu, M.J. Cinnamon Oil Inhibits Shiga Toxin Type 2 Phage Induction and Shiga Toxin Type 2 Production in Escherichia coli O157:H7. Appl. Environ. Microbiol. 2016, 82, 6531–6540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial activity of some essential oils—Present status and future perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Burt, S.A.; Reinders, R.D. Antibacterial activity of selected plant essential oils against Escherichia coli O157:H7. Lett. Appl. Microb. 2003, 36, 162–167. [Google Scholar] [CrossRef] [Green Version]
- De Martino, L.; de Feo, V.; Nazzaro, F. Chemical composition and in vitro antimicrobial and mutagenic activities of seven Lamiaceae essential oils. Molecules 2009, 14, 4213–4230. [Google Scholar] [CrossRef] [Green Version]
- Raybaudi-Massilia, R.M.; Mosqueda-Melgar, J.; Martin-Belloso, O. Antimicrobial activity of essential oils on Salmonella enteritidis, Escherichia coli, and Listeria innocua in fruit juices. J. Food Prot. 2006, 69, 1579–1586. [Google Scholar] [CrossRef] [PubMed]
- Velluti, A.; Sanchis, V.; Ramos, A.J.; Egido, J.; Marın, S. Inhibitory effect of cinnamon, clove, lemongrass, oregano and palmarose essential oils on growth and fumonisin B1 production by Fusarium proliferatum in maize grain. Int. J. Food Microbiol. 2003, 89, 145–154. [Google Scholar] [CrossRef]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A comprehensive review of the antibacterial, antifungal and antiviral potential of essential oils and their chemical constituents against drug-resistant microbial pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef] [PubMed]
- Mutlu-Ingok, A.; Devecioglu, D.; Dikmetas, D.N.; Karbancioglu-Guler, F.; Capanoglu, E. Antibacterial, Antifungal, Antimycotoxigenic, and Antioxidant Activities of Essential Oils: An Updated Review. Molecules 2020, 25, 4711. [Google Scholar] [CrossRef] [PubMed]
- Thapa, D.; Louis, P.; Losa, R.; Zweifel, B.; Wallace, R.J. Essential oils have different effects on human pathogenic and commensal bacteria in mixed faecal fermentations compared with pure cultures. Microbiology 2015, 161, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Mith, H.; Duré, R.; Delcenserie, V.; Zhiri, A.; Daube, G.; Clinquart, A. Antimicrobial activities of commercial essential oils and their components against food-borne pathogens and food spoilage bacteria. Food Sci. Nutr. 2014, 2, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, W.; Gong, J.; Tsao, R.; Zhou, T.; Yu, H.; Poppe, C.; Johnson, R.; Du, Z. Antimicrobial activity of essential oils and structurally related synthetic food additives towards selected pathogenic and beneficial gut bacteria. J. Appl. Microbiol. 2006, 161, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kang, D.H.; Kim, J.K.; Ha, J.G.; Hwang, J.Y.; Kim, T.; Lee, S.H. Antimicrobial activity of plant extracts against Salmonella Typhimurium, Escherichia coli O157:H7, and Listeria monocytogenes on fresh lettuce. J. Food Sci. 2011, 76, M41–M46. [Google Scholar] [CrossRef]
- Kim, Y.G.; Lee, J.H.; Gwon, G.; Kim, S.I.; Park, J.G.; Lee, J. Essential Oils and Eugenols Inhibit Biofilm Formation and the Virulence of Escherichia coli O157:H7. Sci. Rep. 2016, 6, 36377. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kim, Y.G.; Cho, H.S.; Ryu, S.Y.; Cho, M.H.; Lee, L. Coumarins reduce biofilm formation and the virulence of Escherichia coli O157:H7. Phytomedicine 2014, 21, 1037–1042. [Google Scholar] [CrossRef]
- Maggi, F.; Fortuné Randriana, R.; Rasoanaivo, P.; Nicoletti, M.; Quassinti, L.; Bramucci, M.; Lupidi, G.; Petrelli, D.; Vitali, L.A.; Papa, F.; et al. Chemical composition and in vitro biological activities of the essential oil of Vepris macrophylla (Baker) I. Verd. endemic to Madagascar. Chem. Biodivers. 2013, 10, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.A.C.; Borges, A.C.; Brighenti, F.L.; Salvador, M.J.; Gontijo, A.V.L.; Koga-Ito, C.Y. Cymbopogon citratus essential oil: Effect on polymicrobial caries-related biofilm with low cytotoxicity. Braz. Oral Res. 2017, 31, e89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helander, I.M.; Alakomi, H.L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; von Wrigh, A. Characterization of the Action of Selected Essential Oil Components on Gram-Negative Bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Brnawi, W.I.; Hettiarachchy, N.S.; Horax, R.; Kumar-Philips, G.; Seo, H.S.; Marcy, J. Comparison of Cinnamon Essential Oils from Leaf and Bark with Respect to Antimicrobial Activity and Sensory Acceptability in Strawberry Shake. J. Food Sci. 2018, 83, 475–480. [Google Scholar] [CrossRef]
- Giamperi, L.; Bucchini, A.E.A.; Ricci, D.; Tirillini, B.; Nicoletti, M.; Rakotosaona, R.; Maggi, F. Vepris macrophylla (Baker) I. Verd Essential Oil: An Antifungal Agent against Phytopathogenic Fungi. Int. J. Mol. Sci. 2020, 21, 2776. [Google Scholar] [CrossRef] [PubMed]
- Laekeman, G.M.; van Hoof, L.; Haemers, A.; Vanden Berghe, D.A.; Herman, A.G.; Vlietinck, A.J. Eugenol a valuable compound for in vitro experimental research and worthwhile for further in vivo investigation. Phytother. Res. 1990, 4, 90–96. [Google Scholar] [CrossRef]
- Ouattara, B.; Simard, R.E.; Holley, R.A.; Piette, G.J.; Bégin, A. Antibacterial activity of selected fatty acids and essential oils against six meat spoilage organisms. Int. J. Food Microbiol. 1997, 37, 155–162. [Google Scholar] [CrossRef]
- Kim, J.; Marshall, M.R.; Wei, C. Antibacterial Activity of Some Essential Oil Components against Five Foodborne Pathogens. J. Agric. Food Chem. 1995, 43, 2839–2845. [Google Scholar] [CrossRef]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial Activity of Terpenes and Terpenoids Present in Essential Oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.G.; Lee, J.H.; Kim, S.I.; Baek, K.H.; Lee, J. Cinnamon bark oil and its components inhibit biofilm formation and toxin production. Int. J. Food Microbiol. 2015, 195, 30–39. [Google Scholar] [CrossRef]
- Ortega-Ramirez, L.A.; Silva-Espinoza, B.A.; Vargas-Arispuro, I.; Gonzalez-Aguilar, G.A.; Cruz-Valenzuela, M.R.; Nazzaro, F.; Ayala-Zavala, J.F. Combination of Cymbopogon citratus and Allium cepa essential oils increased antibacterial activity in leafy vegetables. J. Sci. Food Agric. 2017, 97, 2166–2173. [Google Scholar] [CrossRef] [PubMed]
- Firmino, D.F.; Cavalcante, T.T.A.; Gomes, G.A.; Firmino, N.C.S.; Rosa, L.D.; de Carvalho, M.G.; Catunda, F.E.A., Jr. Antibacterial and Antibiofilm Activities of Cinnamomum Sp. Essential Oil and Cinnamaldehyde: Antimicrobial Activities. Sci. World J. 2018, 2018, 7405736. [Google Scholar] [CrossRef] [Green Version]
- Vasireddy, L.; Bingle, L.E.H.; Davies, M.S. Antimicrobial activity of essential oils against multidrug-resistant clinical isolates of the Burkholderia cepacia complex. PLoS ONE 2018, 13, e0201835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cáceres, M.; Hidalgo, W.; Stashenko, E.; Torres, R.; Ortiz, C. Essential Oils of Aromatic Plants with Antibacterial, Anti-Biofilm and Anti-Quorum Sensing Activities against Pathogenic Bacteria. Antibiotics 2020, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.E.; Jung, M.; Lee, S.C.; Huh, M.J.; Seo, S.M.; Park, I.K. Antibacterial mode of action of trans-cinnamaldehyde derived from cinnamon bark (Cinnamomum verum) essential oil against Agrobacterium tumefaciens. Pestic. Biochem. Physiol. 2020, 165, 104546. [Google Scholar] [CrossRef]
- Zulfa, Z.; Chia, C.; Rukayadi, Y. In vitro antimicrobial activity of Cymbopogon citratus (lemongrass) extracts against selected foodborne pathogens. Int. Food Res. J. 2016, 23, 1262–1267. [Google Scholar]
- Ortega-Ramirez, L.A.; Gutiérrez-Pacheco, M.M.; Vargas-Arispuro, I.; Gonzales- Aguilar, G.A.; Martínez-Téllez, M.A.; Ayala-Zavala, J.F. Inhibition of Glucosyltransferase Activity and Glucan Production as an Antibiofilm Mechanism of Lemongrass Essential Oil against Escherichia coli O157:H7. Antibiotics 2020, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.; Yuk, H.G. Effects of Sublethal Thymol, Carvacrol, and trans-Cinnamaldehyde Adaptation on Virulence Properties of Escherichia coli O157:H7. Appl. Environ. Microbiol. 2019, 85, e00271-19. [Google Scholar] [CrossRef] [Green Version]
- Matthysse, A.G.; Deora, R.; Mishra, M.; Torres, A.G. Polysaccharides cellulose, poly-beta-1,6-n- acetyl-D-glucosamine, and colanic acid are required for optimal binding of Escherichia coli O157:H7 strains to alfalfa sprouts and K-12 strains to plastic but not for binding to epithelial cells. Appl. Environ. Microbiol. 2008, 74, 2384–2390. [Google Scholar] [CrossRef] [Green Version]
- Orme, R.; Douglas, C.W.; Rimmer, S.; Webb, M. Proteomic analysis of Escherichia coli biofilms reveals the overexpression of the outer membrane protein OmpA. Proteomics 2006, 6, 4269–4277. [Google Scholar] [CrossRef]
- Qun, M.; Wood, T.K. OmpA influences Escherichia coli biofilm formation by repressing cellulose production through the CpxRA two-component system. Environ. Microbiol. 2009, 11, 2735–2746. [Google Scholar] [CrossRef]
- Barrios, A.F.; Zuo, R.; Ren, D.; Wood, T.K. Hha, YbaJ, and OmpA regulate Escherichia coli K12 biofilm formation and conjugation plasmids abolish motility. Biotechnol. Bioeng. 2006, 93, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Kaper, J.B. Multiple elements controlling adherence of enterohemorrhagic Escherichia coli O157:H7 to HeLa cells. Infect. Immun. 2003, 71, 4985–4995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mith, H.; Clinquart, A.; Zhiri, A.; Daube, G.; Delcenserie, V. The impact of oregano (Origanum heracleoticum) essential oil and carvacrol on virulence gene transcription by Escherichia coli O157:H7. FEMS Microbiol. Lett. 2015, 362, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Khatibi, S.A.; Misaghi, A.; Moosavy, M.H.; Akhondzadeh Basti, A.; Mohamadian, S.; Khanjari, A. Effect of nanoliposomes containing Zataria multiflora Boiss. essential oil on gene expression of Shiga toxin 2 in Escherichia coli O157:H7. J. Appl. Microbiol. 2018, 124, 389–397. [Google Scholar] [CrossRef]
- Takemasa, N.; Ohnishi, S.; Tsuji, M.; Shikata, T.; Yokoigawa, K. Screening and analysis of spices with ability to suppress verocytotoxin production by Escherichia coli O157:H7. J. Food Sci. 2009, 74, M461–M466. [Google Scholar] [CrossRef]
- Lim, J.; Lee, K.M.; Kim, S.H.; Kim, Y.; Kim, S.H.; Park, W.; Park, S. YkgM and ZinT proteins are required for maintaining intracellular zinc concentration and producing curli in enterohemorrhagic Escherichia coli (EHEC) O157:H7 under zinc deficient conditions. Int. J. Food Microbiol. 2011, 149, 159–170. [Google Scholar] [CrossRef]
- Crane, J.K.; Broome, J.E.; Reddinger, R.M.; Werth, B.B. Zinc protects against shiga-toxigenic Escherichia coli by acting on host tissues as well as on bacteria. BMC Microbiol. 2014, 14, 145. [Google Scholar] [CrossRef] [Green Version]
- Kumaran, A.M.; D’Souza, P.; Agarwal, A.; Bokkolla, R.M.; Balasubramaniam, M. Geraniol, the putative anthelmintic principle of Cymbopogon martini. Phytother. Res. 2003, 17, 957. [Google Scholar] [CrossRef]
- Chou, T.C.; Chiu, C.; Kuo, C.J.; Wu, C.M.; Syu, W.J.; Chiu, W.T.; Chen, C.S. Enterohaemorrhagic Escherichia coli O157:H7 Shiga-like toxin 1 is required for full pathogenicity and activation of the p38 mitogen-activated protein kinase pathway in Caenorhabditis elegans. Cell. Microbiol. 2013, 15, 82–97. [Google Scholar] [CrossRef]
- Erfan, A.M.; Marouf, S. Cinnamon oil downregulates virulence genes of poultry respiratory bacterial agents and revealed significant bacterial inhibition: An in vitro perspective. Vet. World 2019, 12, 1707–1715. [Google Scholar] [CrossRef] [Green Version]
- Maggi, F.; Cecchini, C.; Cresci, A.; Coman, M.M.; Tirillini, B.; Sagratini, G.; Papa, F.; Vittori, S. Chemical composition and antimicrobial activity of the essential oils from several Hypericum taxa (Guttiferae) growing in central Italy (Appennino Umbro-Marchigiano). Chem. Biodivers. 2010, 7, 447–466. [Google Scholar] [CrossRef] [PubMed]
- Ornano, L.; Venditti, A.; Ballero, M.; Sanna, C.; Quassinti, L.; Bramucci, M.; Lupidi, G.; Papa, F.; Vittori, S.; Maggi, F.; et al. Chemopreventive and antioxidant activity of the chamazulene-rich essential oil obtained from Artemisia arborescens L. growing on the Isle of La Maddalena, Sardinia, Italy. Chem. Biodivers. 2013, 10, 1464–1474. [Google Scholar] [CrossRef] [PubMed]
- Scotti, R.; Nicolini, L.; Gabbianelli, R. Sod C genes expression in Escherichia coli O157:H7 strains. Ann. Ist. Super. Sanità 2016, 52, 309–312. [Google Scholar] [CrossRef]
- Gabbianelli, R.; Scotti, R.; Ammendola, S.; Petrarca, P.; Nicolini, L.; Battistoni, A. Role of ZnuABC and ZinT in Escherichia coli O157:H7 zinc acquisition and interactior with epithelial cells. BMC Microbiol. 2011, 11, 11–36. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institutes (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; M07 Ed.11; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Scotti, R.; Nicolini, L.; Stringaro, A.; Gabbianelli, R. A study on prophagic and chromosomal sodC genes involvement in Escherichia coli O157:H7 biofilm formation and biofilm resistance to H2O2. Ann. Ist. Super. Sanità 2015, 51, 62–66. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Amrit, F.R.; Ratnappan, R.; Keith, S.A.; Ghazi, A. The C. elegans lifespan assay toolkit. Methods 2014, 68, 465–475. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EO | Main Components |
|---|---|
| Cymbopogon flexuosus | geranial (41.5%), neral (30.0%), geranyl acetate (6.1%), geraniol (5.5%), caryophyllene oxide (2.8%), γ-cadinene (2.6%) |
| Cymbopogon citratus | geranial (48.2%), neral (32%), geranyl acetate (3.8%), geraniol (3.1%), camphene (1.9%), (E)-caryophyllene |
| Cymbopogon martini | geraniol (82.5%), geranyl acetate (11.1%), (E)-caryophyllene (2.2%), linalool (2.1%) |
| Cinnamomum verum (bark) | (E)-cinnamaldehyde (85.4%), eugenol (7.0%), (E)-cinnamyl acetate (4.5%) |
| Cinnamomum verum (leaf) | eugenol (83.5%), benzyl benzoate (2.9%), (E)-caryophyllene (2.8%), eugenol acetate (1.9%), α-humulene (1.2%), (E)-cinnamyl acetate (1.2%) |
| Strain/EO | C. flexuosus | C. citratus | C. martini | C. verum (Bark) | C. verum (Leaf) |
|---|---|---|---|---|---|
| EDL933 | MIC 0.075 | MIC 0.075 | MIC 0.100 | MIC 0.0075 | MIC 0.175 |
| MBC 0.400 | MBC 0.200 | MBC 0.400 | MBC 0.0800 | MBC 0.200 | |
| ED597 | MIC 0.075 | MIC 0.075 | MIC 0.075 | MIC 0.0075 | MIC 0.150 |
| MBC 0.300 | MBC 0.200 | MBC 0.300 | MBC 0.0200 | MBC 0.200 | |
| EDB | MIC 0.075 | MIC 0.200 | MIC 0.100 | MIC 0.0075 | MIC 0.175 |
| MBC 0.400 | MBC 0.400 | MBC 0.200 | MBC 0.0600 | MBC 0.200 | |
| ED419 | MIC 0.100 | MIC 0.100 | MIC 0.100 | MIC 0.0075 | MIC 0.175 |
| MBC 0.300 | MBC 0.200 | MBC 0.200 | MBC 0.0800 | MBC 0.300 | |
| K12 | MIC 0.150 | MIC 0.200 | MIC 0.300 | MIC 0.0075 | MIC 0.175 |
| MBC 0.400 | MBC 0.400 | MBC 0.400 | MBC 0.0400 | MBC 0.300 |
| Oligo Name | Sequence 5’-3’ |
|---|---|
| ant43 F/R | ACAAATGGTCGTCAGGTCGT |
| csgA F/R | CCCGTATACGAGTTGTCAGA |
| GCTCAATCGATCTGACCCAA | |
| ehaA F/R | TTACCAAAGCCAACCTGAGT |
| CAGCCGTTTGTAGAAGTGAA | |
| fimA F/R | GCAGAGGTGTCATTATATCCC |
| CGTTCAGTTAGGACAGGTTC | |
| fliA F/R | TTATTCAGGGTTGTTTGCTCA |
| GTAAGTTGTAAATGCCGTTCC | |
| pgaA F/R | GCTGAAGGTGTAATGGATAAAC |
| AGGGACTGCGCATTGATTAC | |
| ompA F/R | GTTCACGTTCGACAACATCG |
| GTTGTAAGCGTCAGAACCGA | |
| stx2 F/R | ACAGACCAAGCACTTCACTC |
| CGTTCCGGAATGCAAATCAG | |
| ykgM F/R | GCGTCATCGTATACACAGGA |
| TACTGTGGTGTTCCACGACACC | |
| zinT F/R | CCTGTATAGAACGGGTGCGATT |
| ACGGCAAACCCTTAACAGA | |
| 16 s F/R | CTCCATCCCAGTCACTGAG |
| CATCCACAGAACTTTCCAGAG | |
| CCAACATTTCACAACACGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scotti, R.; Stringaro, A.; Nicolini, L.; Zanellato, M.; Boccia, P.; Maggi, F.; Gabbianelli, R. Effects of Essential Oils from Cymbopogon spp. and Cinnamomum verum on Biofilm and Virulence Properties of Escherichia coli O157:H7. Antibiotics 2021, 10, 113. https://doi.org/10.3390/antibiotics10020113
Scotti R, Stringaro A, Nicolini L, Zanellato M, Boccia P, Maggi F, Gabbianelli R. Effects of Essential Oils from Cymbopogon spp. and Cinnamomum verum on Biofilm and Virulence Properties of Escherichia coli O157:H7. Antibiotics. 2021; 10(2):113. https://doi.org/10.3390/antibiotics10020113
Chicago/Turabian StyleScotti, Raffaella, Annarita Stringaro, Laura Nicolini, Miriam Zanellato, Priscilla Boccia, Filippo Maggi, and Roberta Gabbianelli. 2021. "Effects of Essential Oils from Cymbopogon spp. and Cinnamomum verum on Biofilm and Virulence Properties of Escherichia coli O157:H7" Antibiotics 10, no. 2: 113. https://doi.org/10.3390/antibiotics10020113
APA StyleScotti, R., Stringaro, A., Nicolini, L., Zanellato, M., Boccia, P., Maggi, F., & Gabbianelli, R. (2021). Effects of Essential Oils from Cymbopogon spp. and Cinnamomum verum on Biofilm and Virulence Properties of Escherichia coli O157:H7. Antibiotics, 10(2), 113. https://doi.org/10.3390/antibiotics10020113