
Molecular Analysis of Polymyxin Resistance among Carbapenemase-Producing Klebsiella pneumoniae in Colombia

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility
2.2. Molecular Typing and Virulome Analysis
2.3. Resistome Analysis
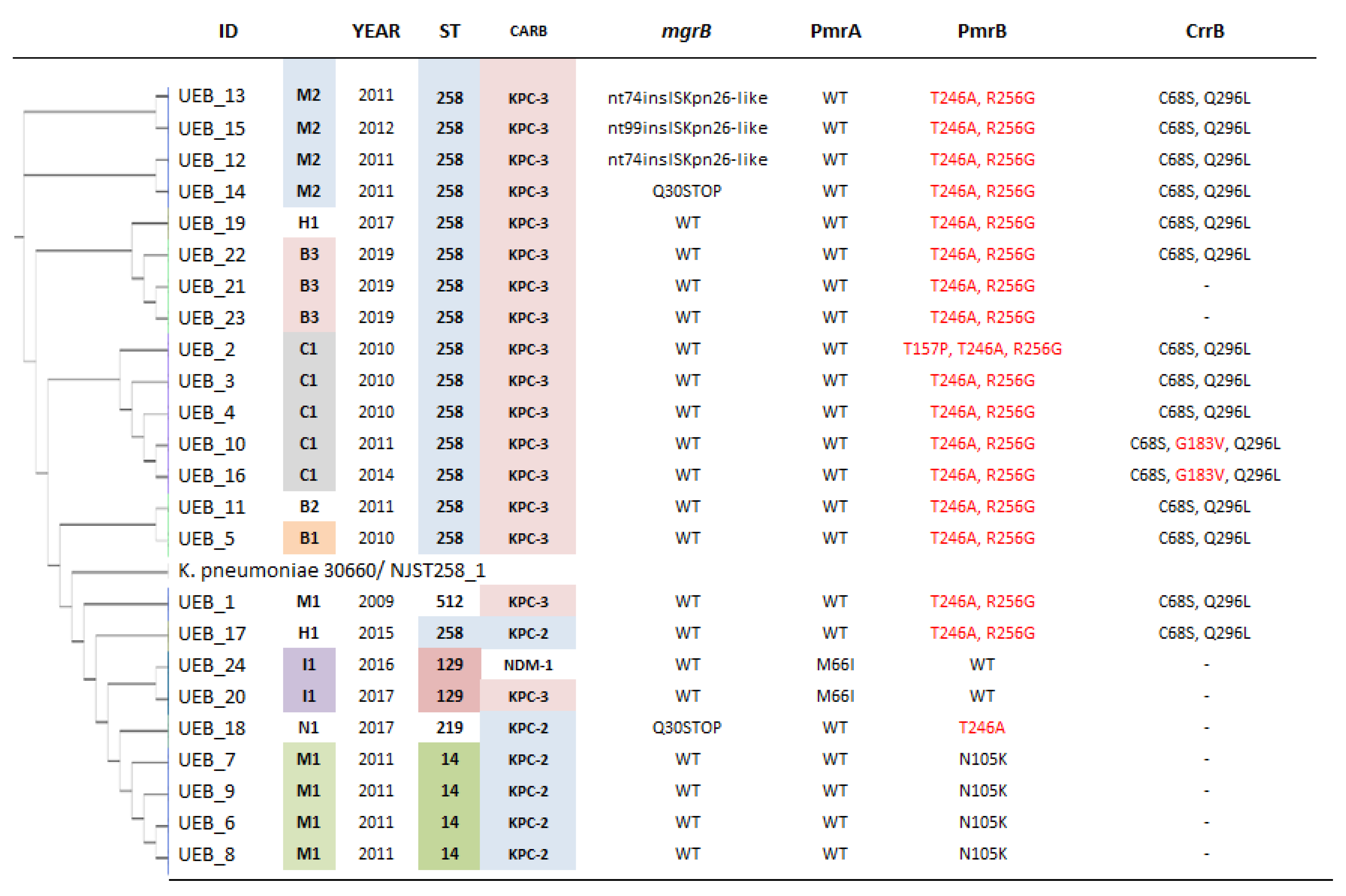
2.4. Mechanisms of Resistance towards Polymyxins
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates and Antimicrobial Susceptibility
4.2. Molecular Typing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Duin, D.; Perez, F.; Rudin, S.D.; Cober, E.; Hanrahan, J.; Ziegler, J.; Webber, R.; Fox, J.; Mason, P.; Richter, S.S.; et al. Surveillance of carbapenem-resistant Klebsiella pneumoniae: Tracking molecular epidemiology and outcomes through a regional network. Antimicrob. Agents Chemother. 2014, 58, 4035–4041. [Google Scholar] [CrossRef] [Green Version]
- Rojas, L.J.; Salim, M.; Cober, E.; Richter, S.S.; Perez, F.; Salata, R.A.; Kalayjian, R.C.; Watkins, R.R.; Marshall, S.; Rudin, S.D.; et al. Colistin Resistance in Car-bapenem-Resistant Klebsiella pneumoniae: Laboratory Detection and Impact on Mortality. Clin. Infect. Dis. 2017, 64, 711–718. [Google Scholar] [PubMed] [Green Version]
- Queenan, A.M.; Bush, K. Carbapenemases: The Versatile β-Lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Sun, X.; Ma, X. Systematic review and meta-analysis of mortality of patients infected with carbapenem-resistant Klebsiella pneumoniae. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavascki, A.P.; Goldani, L.Z.; Li, J.; Nation, R.L. Polymyxin B for the treatment of multidrug-resistant pathogens: A critical review. J. Antimicrob. Chemother. 2007, 60, 1206–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.; Brunel, J.M.; Dubus, J.C.; Reynaud-Gaubert, M.; Rolain, J.M. Colistin: An update on the antibiotic of the 21st century. Expert Rev. Anti-Infect. Ther. 2012, 10, 917–934. [Google Scholar] [CrossRef] [PubMed]
- Jeannot, K.; Bolard, A.; Plésiat, P. Resistance to polymyxins in Gram-negative organisms. Int. J. Antimicrob. Agents 2017, 49, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Cannatelli, A.; Giani, T.; D’Andrea, M.M.; Di Pilato, V.; Arena, F.; Conte, V.; Tryfinopoulou, K.; Vatopoulos, A.; Rossolini, G.M. MgrB Inactivation Is a Common Mechanism of Colistin Resistance in KPC-Producing Klebsiella pneumoniae of Clinical Origin. Antimicrob. Agents Chemother. 2014, 58, 5696–5703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jana, B.; Cain, A.K.; Doerrler, W.T.; Boinett, C.J.; Fookes, M.C.; Parkhill, J.; Guardabassi, L. The secondary resistome of multidrug-resistant Klebsiella pneumoniae. Sci. Rep. 2017, 7, srep42483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, L.J.; Weinstock, G.M.; De La Cadena, E.; Diaz, L.; Rios, R.; Hanson, B.M.; Brown, J.S.; Vats, P.; Phillips, D.S.; Nguyen, H.; et al. An Analysis of the Epidemic of Klebsiella pneumoniae Carbapenemase-Producing K. pneumoniae: Convergence of Two Evolutionary Mechanisms Creates the “Perfect Storm”. J. Infect. Dis. 2018, 217, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Jayol, A.; Bontron, S.; Villegas, M.-V.; Ozdamar, M.; Türkoglu, S.; Nordmann, P. The mgrB gene as a key target for acquired resistance to colistin in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2014, 70, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Bogdanovich, T.; Adams-Haduch, J.M.; Tian, G.-B.; Nguyen, M.H.; Kwak, E.J.; Muto, C.A.; Doi, Y. Colistin-Resistant, Klebsiella pneumoniae Carbapenemase (KPC)-Producing Klebsiella pneumoniae Belonging to the International Epidemic Clone ST258. Clin. Infect. Dis. 2011, 53, 373–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behera, B.; Mathur, P.; Das, A.; Kapil, A.; Gupta, B.; Bhoi, S.; Farooque, K.; Sharma, V.; Misra, M. Evaluation of susceptibility testing methods for polymyxin. Int. J. Infect. Dis. 2010, 14, e596–e601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multi-drug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2012, 18, 268–281. [Google Scholar]
- Cannatelli, A.; D’Andrea, M.M.; Giani, T.; Di Pilato, V.; Arena, F.; Ambretti, S.; Gaibani, P.; Rossolini, G.M. In vivo emergence of colistin resistance in Klebsiella pneumoniae producing KPC-type carbapenemases mediated by insertional inactivation of the PhoQ/PhoP mgrB regulator. Antimicrob. Agents Chemother. 2013, 57, 5521–5526. [Google Scholar] [CrossRef] [Green Version]
- Jaidane, N.; Bonnin, R.A.; Mansour, W.; Girlich, D.; Creton, E.; Cotellon, G.; Chaouch, C.; Boujaafar, N.; Bouallegue, O.; Naas, T. Genomic Insights into Colistin-Resistant Klebsiella pneumoniae from a Tunisian Teaching Hospital. Antimicrob. Agents Chemother. 2017, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayol, A.; Poirel, L.; Villegas, M.-V.; Nordmann, P. Modulation of mgrB gene expression as a source of colistin resistance in Klebsiella oxytoca. Int. J. Antimicrob. Agents 2015, 46, 108–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayol, A.; Nordmann, P.; Brink, A.; Villegas, M.-V.; Dubois, V.; Poirel, L. High-Level Resistance to Colistin Mediated by Various Mutations in the crrB Gene among Carbapenemase-Producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2017, 61, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Biswas, L.; Omgy, N.; Mohan, K.; Vinod, V.; Sajeev, A.; Nair, P.; Singh, S.; Biswas, R. Colistin resistance due to inser-tional inactivation of the mgrB in klebsiella pneumoniae of clinical origin: First report from India. Rev. Esp. Quimioter. 2018, 31, 406–410. [Google Scholar]
- Novović, K.; Trudić, A.; Brkić, S.; Vasiljević, Z.; Kojić, M.; Medić, D.; Ćirković, I.; Jovčić, B. Molecular Epidemiology of Colistin-Resistant, Carbapenemase-Producing Klebsiella pneumoniae in Serbia from 2013 to 2016. Antimicrob. Agents Chemother. 2017, 61, e02550-16. [Google Scholar] [CrossRef] [Green Version]
- Olaitan, A.O.; Diene, S.M.; Kempf, M.; Berrazeg, M.; Bakour, S.; Gupta, S.K.; Thongmalayvong, B.; Akkhavong, K.; Somphavong, S.; Paboriboune, P.; et al. Worldwide emergence of colistin resistance in Klebsiella pneumoniae from healthy humans and pa-tients in Lao PDR, Thailand, Israel, Nigeria and France owing to inactivation of the PhoP/PhoQ regulator mgrB: An epidemi-ological and molecular study. Int. J. Antimicrob. Agents 2014, 44, 500–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitt, M.E.; Elliot, A.G.; Cao, M.D.; Ganesamoorthy, D.; Karaiskos, I.; Giamarellou, H.; Abboud, C.S.; Blaskovich, M.A.T.; Cooper, M.A.; Coin, L.J.M. Multifactorial chromosomal variants regulate polymyxin resistance in extensively drug-resistant Klebsiella pneumoniae. Microb. Genom. 2018, 4, e000158. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-H.; Lin, T.-L.; Pan, Y.-J.; Wang, Y.-P.; Lin, Y.-T.; Wang, J.-T. Colistin Resistance Mechanisms in Klebsiella pneumoniae Strains from Taiwan. Antimicrob. Agents Chemother. 2015, 59, 2909–2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayol, A.; Poirel, L.; Brink, A.; Villegas, M.-V.; Yilmaz, M.; Nordmann, P. Resistance to Colistin Associated with a Single Amino Acid Change in Protein PmrB among Klebsiella pneumoniae Isolates of Worldwide Origin. Antimicrob. Agents Chemother. 2014, 58, 4762–4766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheong, H.S.; Kim, S.Y.; Seo, J.; Wi, Y.M.; Peck, K.R.; Ko, K.S. Colistin Resistance and Extensive Genetic Variations in PmrAB and PhoPQ in Klebsiella Pneumoniae Isolates from South Korea. Curr. Microbiol. 2020, 77, 2307–2311. [Google Scholar] [CrossRef]
- Jayol, A.; Poirel, L.; Dortet, L.; Nordmann, P. 2016. National survey of colistin resistance among carbapenemase-producing Enterobacteriaceae and outbreak caused by colistin-resistant OXA-48-producing Klebsiella pneumoniae, France, 2014. Eurosurveillance 2016, 21, 30339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojica, M.F.; Correa, A.; Vargas, D.A.; Maya, J.J.; Montealegre, M.C.; Rojas, L.J.; Ruiz, S.J.; Quinn, J.P.; Villegas, M.V.; Colombian Nosocomial Bacterial Resistance Study Group. Molecular correlates of the spread of KPC-producing Enterobacteriaceae in Colombia. Int. J. Antimicrob. Agents 2012, 40, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Ocampo, A.M.; Vargas, C.A.; Sierra, P.M.; Cienfuegos, A.V.; Jiménez, J.N. Caracterización molecular de un brote de Klebsiella pneumoniae resistente a carbapenémicos en un hospital de alto nivel de complejidad de Medellín, Colombia. Biomédica 2015, 35, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Lalaoui, R.; Bakour, S.; Livnat, K.; Assous, M.V.; Diene, S.M.; Rolain, J.-M. Spread of Carbapenem and Colistin-Resistant Klebsiella pneumoniae ST512 Clinical Isolates in Israel: A Cause for Vigilance. Microb. Drug Resist. 2019, 25, 63–71. [Google Scholar] [CrossRef]
- Giani, T.; Pini, B.; Arena, F.; Conte, V.; Bracco, S.; Migliavacca, R.; Pantosti, A.; Pagani, L.; Luzzaro, F.; Rossolini, G.M. Epidemic diffusion of KPC carbapenemase-producing Klebsiella pneumoniae in Italy: Results of the first countrywide survey, 15 May to 30 June 2011. Eurosurveillance 2013, 18, 1–9. [Google Scholar]
- Cienfuegos-Gallet, A.V.; Chen, L.; Kreiswirth, B.N.; Jiménez, J.N. Colistin Resistance in Carbapenem-Resistant Klebsiella pneumoniae Mediated by Chromosomal Integration of Plasmid DNA. Antimicrob. Agents Chemother. 2017, 61, e00404-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saavedra, S.Y.; Diaz, L.; Wiesner, M.; Correa, A.; Arévalo, S.A.; Reyes, J.; Hidalgo, A.M.; De La Cadena, E.; Perenguez, M.; Montaño, L.A.; et al. Genomic and Molecular Characterization of Clinical Isolates of Enterobacteriaceae Harboring mcr-1 in Colombia, 2002 to 2016. Antimicrob. Agents Chemother. 2017, 61, e00841-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berglund, B. Acquired Resistance to Colistin via Chromosomal And Plasmid-Mediated Mechanisms in Klebsiella pneumoniae. Infect. Microbes. Dis. 2019, 1, 10–19. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard-Thenth Edition; CLSI Document M07-A10; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Correa, A.; Del Campo, R.; Perenguez, M.; Blanco, V.M.; Rodríguez-Baños, M.; Perez, F.; Maya, J.J.; Rojas, L.; Cantón, R.; Arias, C.A.; et al. Dissemination of High-Risk Clones of Extensively Drug-Resistant Pseudomonas aeruginosa in Colombia. Antimicrob. Agents Chemother. 2015, 59, 2421–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding data and analysis capabilities. Nucleic Acids Res. 2019, 48, D606–D612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In SilicoDetection and Typing of Plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Chan, M.-S.; Maiden, M.C.J.; Spratt, B.G. Database-driven Multi Locus Sequence Typing (MLST) of bacterial pathogens. Bioinformatics 2001, 17, 1077–1083. [Google Scholar] [CrossRef]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequenc-es. Nucleic Acids Res. 2006, 34, 32–36. [Google Scholar] [CrossRef] [Green Version]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| ID | HOSPITAL | YEAR | SOURCE | ROOM | POL | CZA | CAZ | FEP | TZP | ETP | IMI | MEM | DOR | CIP | TGC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| UEB_1 | M1 | 2009 | BLOOD | HOSP | 4 | 4/4 | >32 | >64 | >64/4 | >32 | >32 | >32 | >16 | >2 | 4 |
| UEB_2 | C1 | 2010 | SKIN | ER | 8 | ≤1/4 | >32 | >64 | >64/4 | >32 | 32 | >32 | >16 | >2 | ≤0.5 |
| UEB_3 | C1 | 2010 | BLOOD | ICU | 8 | 4/4 | >32 | >64 | >128/4 | >32 | 32 | >32 | >16 | >2 | ≤0.5 |
| UEB_4 | C1 | 2010 | SKIN | ICU | >8 | 4/4 | >32 | >64 | >128/4 | >32 | >32 | >32 | >16 | >2 | 1 |
| UEB_5 | B1 | 2010 | OTHER | HOSP | 4 | ≤1/4 | >32 | 64 | >128/4 | 2 | 1 | 1 | 1 | >2 | ≤0.5 |
| UEB_6 | M1 | 2011 | GI | ICU | >8 | 2/4 | >32 | 16 | >128/4 | 16 | 4 | 4 | 4 | 1 | ≤0.5 |
| UEB_7 | M1 | 2011 | SECRETION | ICU | 8 | 4/4 | >32 | 16 | >128/4 | >32 | 16 | >32 | >16 | 1 | 1 |
| UEB_8 | M1 | 2011 | GI | ICU | >8 | 2/4 | >32 | >64 | >128/4 | >32 | 16 | 32 | >16 | 1 | 1 |
| UEB_9 | M1 | 2011 | GI | ICU | 8 | ≤1/4 | >32 | 16 | 128/4 | 2 | 1 | 1 | ≤0.5 | 1 | 1 |
| UEB_10 | C1 | 2011 | GI | ICU | >8 | ≤1/4 | >32 | 64 | >128/4 | 32 | 4 | 16 | 16 | >2 | 2 |
| UEB_11 | B2 | 2011 | BLOOD | ICU | >8 | 4/4 | >33 | >64 | >128/4 | >32 | >32 | >32 | >16 | >2 | 2 |
| UEB_12 | M2 | 2011 | RESP TRACT | ICU | >8 | 4/4 | >32 | >64 | >128/4 | >32 | >32 | >32 | >16 | >2 | 1 |
| UEB_13 | M2 | 2011 | BLOOD | ICU | >8 | 4/4 | >32 | 64 | >128/4 | >32 | >32 | >32 | >16 | >2 | ≤0.5 |
| UEB_14 | M2 | 2011 | GI | ICU | >8 | 2/4 | >32 | 64 | >128/4 | >32 | >32 | >32 | >16 | >2 | 1 |
| UEB_15 | M2 | 2012 | URINE | HOSP | >8 | 4/4 | >32 | 64 | >128/4 | >32 | >32 | >32 | >16 | >2 | 1 |
| UEB_16 | C1 | 2014 | BLOOD | HOSP | >8 | ≤1/4 | >32 | >64 | >128/4 | 32 | >32 | >32 | >16 | >2 | 4 |
| UEB_17 | H1 | 2015 | RESP TRACT | ICU | >8 | 2/4 | >32 | 64 | >128/4 | >32 | >32 | >32 | >16 | 1 | 2 |
| UEB_18 | N1 | 2017 | BLOOD | ER | >8 | ≤1/4 | >32 | 32 | >128/4 | 4 | 2 | 1 | 2 | ≤0.25 | 4 |
| UEB_19 | H1 | 2017 | URINE | HOSP | >8 | ≤1/4 | >32 | 64 | >128/4 | >32 | 32 | 32 | >16 | >2 | 4 |
| UEB_20 | I1 | 2017 | URINE | HOSP | >8 | ≤1/4 | >32 | >64 | >128/4 | 32 | 8 | 16 | 16 | 1 | 2 |
| UEB_21 | B3 | 2019 | RESP TRACT | HOSP | >8 | 4/4 | >32 | >64 | >128/4 | >32 | >32 | >32 | >16 | >2 | 1 |
| UEB_22 | B3 | 2019 | PERIT LIQ | HOSP | >8 | 2/4 | >32 | >64 | >128/4 | >32 | >32 | >32 | >16 | >2 | 1 |
| UEB_23 | B3 | 2019 | ASCIT LIQ | HOSP | >8 | 4/4 | >32 | >64 | >128/4 | >32 | >32 | >32 | >16 | >2 | 1 |
| UEB_24 | I1 | 2016 | RESP TRACT | HOSP | >8 | >64/4 | >32 | >64 | >128/4 | >32 | >32 | 32 | >16 | >2 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De La Cadena, E.; Mojica, M.F.; García-Betancur, J.C.; Appel, T.M.; Porras, J.; Pallares, C.J.; Solano-Gutiérrez, J.S.; Rojas, L.J.; Villegas, M.V. Molecular Analysis of Polymyxin Resistance among Carbapenemase-Producing Klebsiella pneumoniae in Colombia. Antibiotics 2021, 10, 284. https://doi.org/10.3390/antibiotics10030284
De La Cadena E, Mojica MF, García-Betancur JC, Appel TM, Porras J, Pallares CJ, Solano-Gutiérrez JS, Rojas LJ, Villegas MV. Molecular Analysis of Polymyxin Resistance among Carbapenemase-Producing Klebsiella pneumoniae in Colombia. Antibiotics. 2021; 10(3):284. https://doi.org/10.3390/antibiotics10030284
Chicago/Turabian StyleDe La Cadena, Elsa, María Fernanda Mojica, Juan Carlos García-Betancur, Tobías Manuel Appel, Jessica Porras, Christian José Pallares, Juan Sebastián Solano-Gutiérrez, Laura J. Rojas, and María Virginia Villegas. 2021. "Molecular Analysis of Polymyxin Resistance among Carbapenemase-Producing Klebsiella pneumoniae in Colombia" Antibiotics 10, no. 3: 284. https://doi.org/10.3390/antibiotics10030284
APA StyleDe La Cadena, E., Mojica, M. F., García-Betancur, J. C., Appel, T. M., Porras, J., Pallares, C. J., Solano-Gutiérrez, J. S., Rojas, L. J., & Villegas, M. V. (2021). Molecular Analysis of Polymyxin Resistance among Carbapenemase-Producing Klebsiella pneumoniae in Colombia. Antibiotics, 10(3), 284. https://doi.org/10.3390/antibiotics10030284