
Occurrence and Antimicrobial Resistance of Arcobacter spp. Recovered from Aquatic Environments

 ,
,  and
and
Abstract
:1. Introduction
2. Results
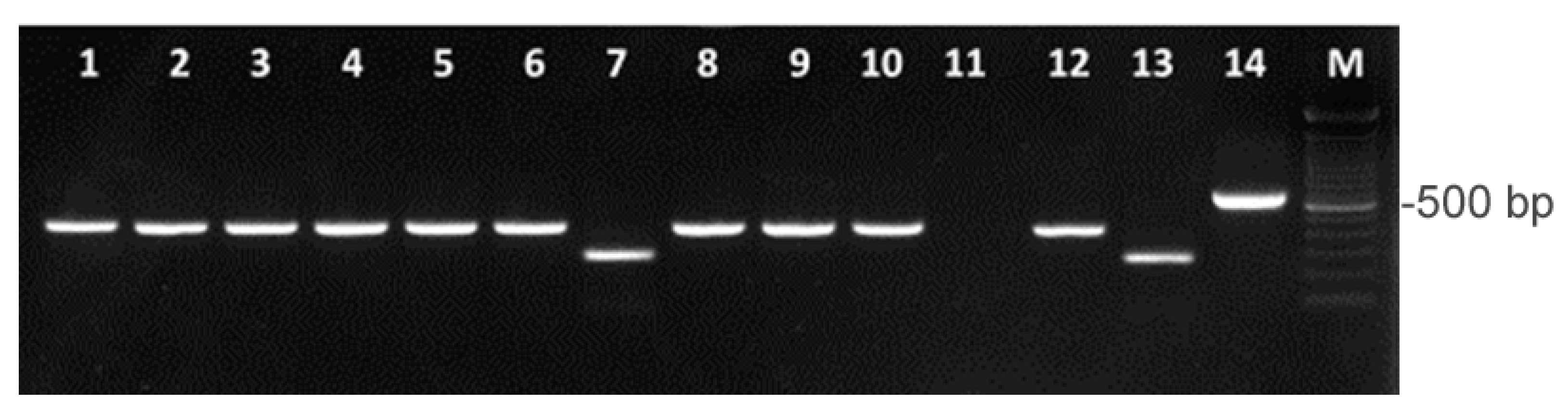
2.1. Isolation and Identification of Arcobacter Species
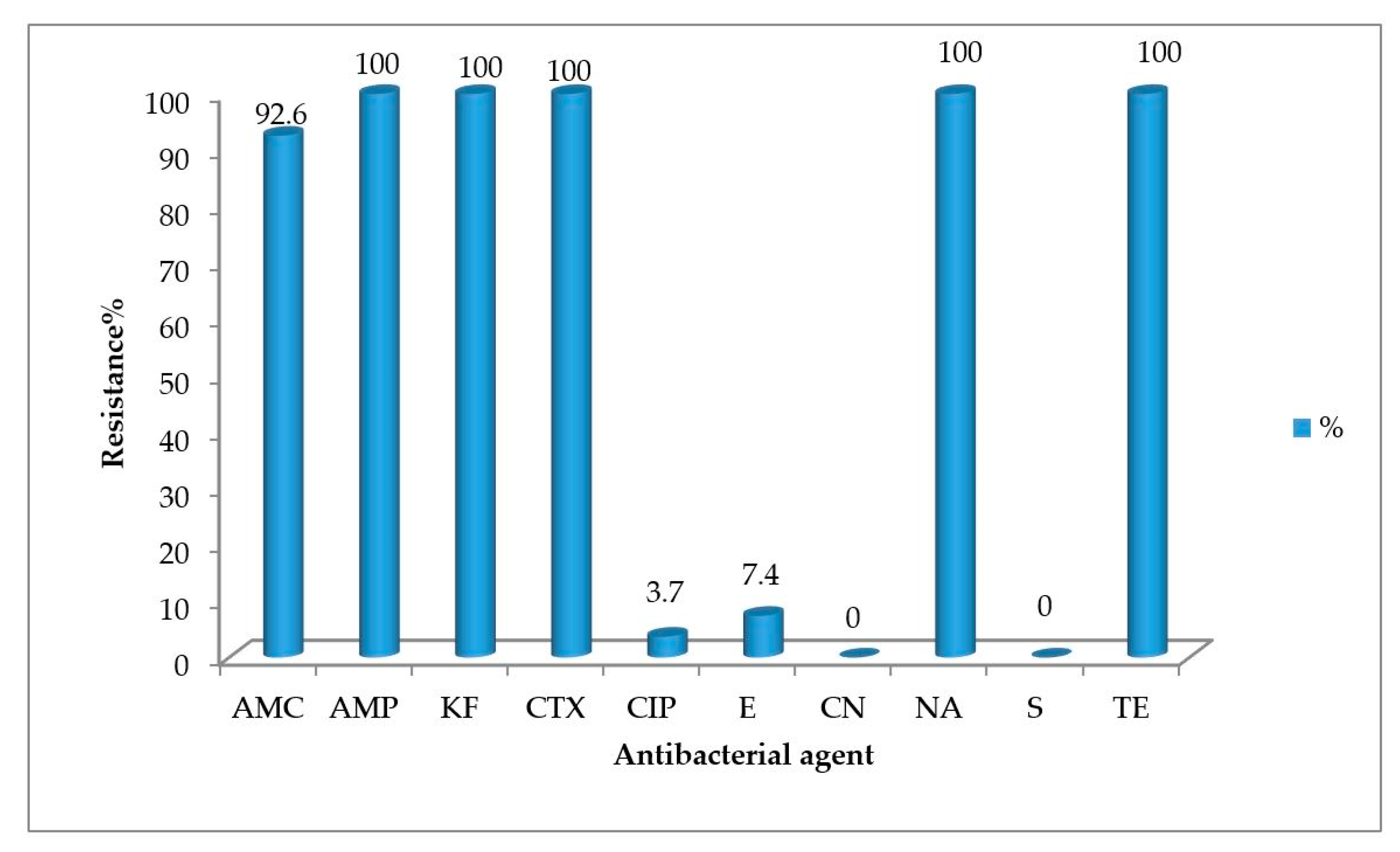
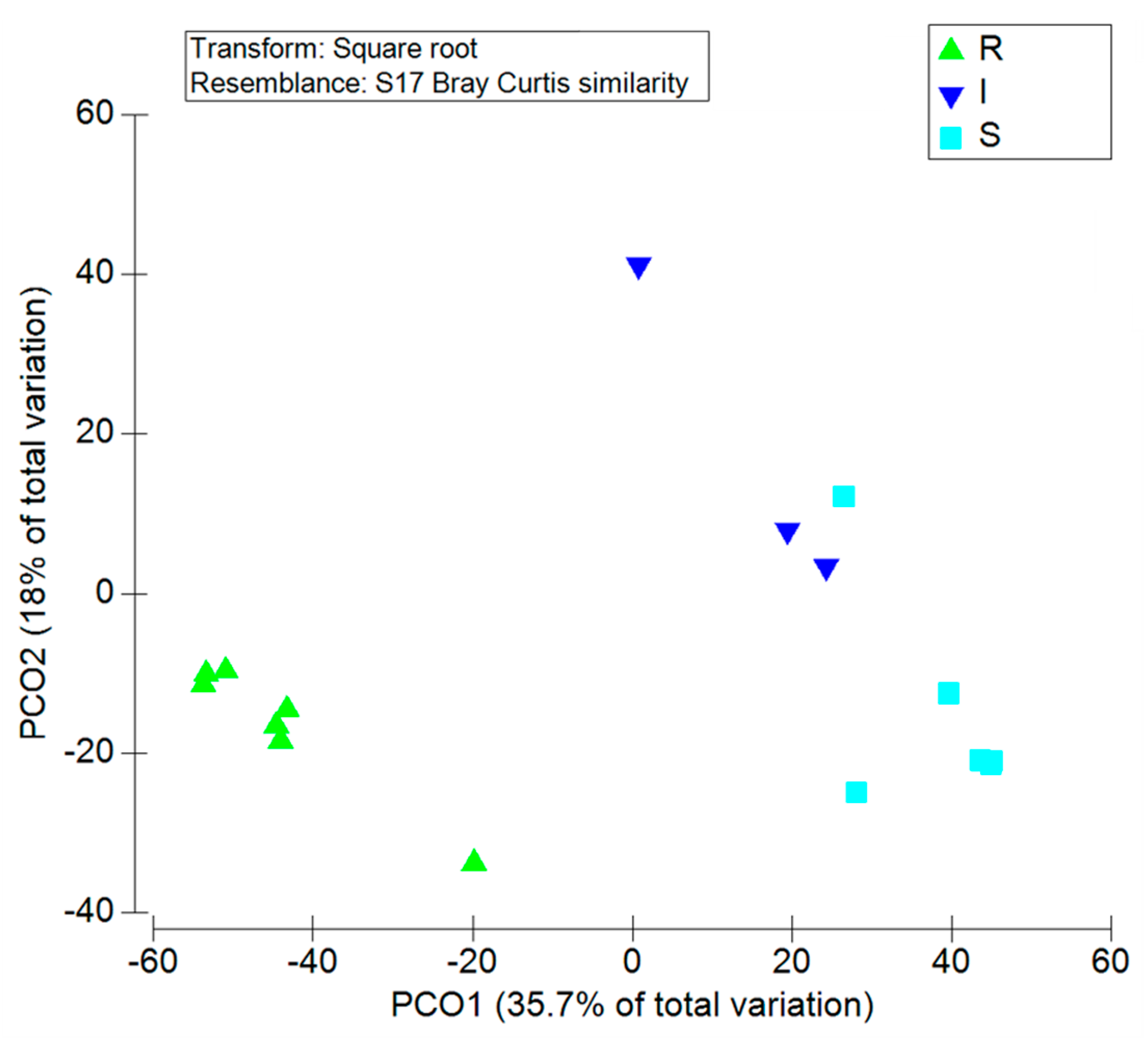
2.2. Antimicrobial Susceptibility Testing
2.3. Analysis of the Quinolone and Tetracycline Resistance Genes
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Isolation of Arcobacter
4.3. Identification of Arcobacter Species by Multiplex PCR
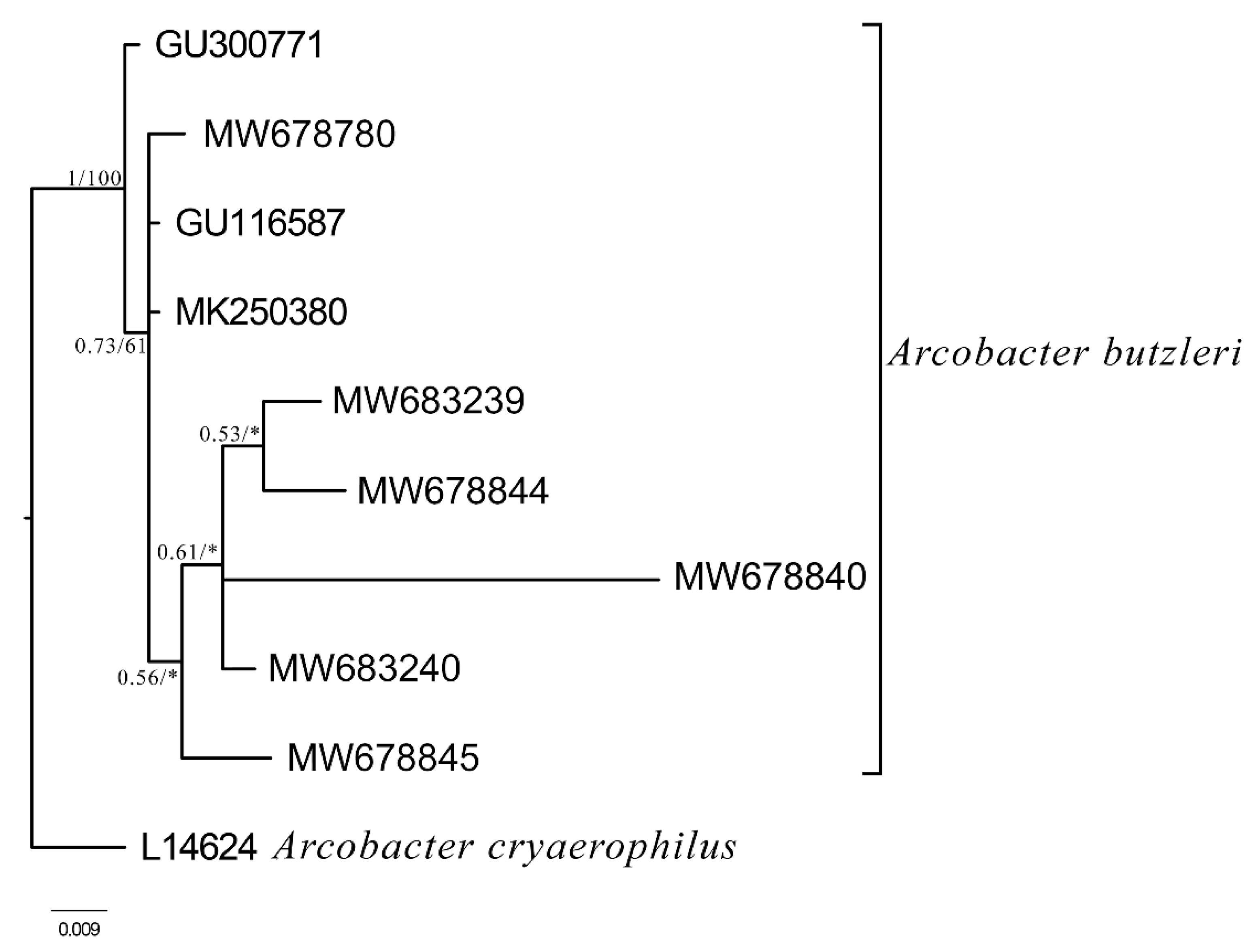
4.4. Identification of Arcobacter Species by Sequence Analysis
4.5. Antimicrobial Susceptibility Testing
4.6. Analysis of the Quinolone and Tetracycline Resistance Genes
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vandamme, P.; De Ley, J. Proposal for a New Family, Campylobacteraceae. Int. J. Syst. Bacteriol. 1991, 41, 451–455. [Google Scholar] [CrossRef]
- Kjeldgaard, J.; Jørgensen, K.; Ingmer, H. Growth and Survival at Chiller Temperatures of Arcobacter butzleri. Int. J. Food Microbiol. 2009, 131, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Collado, L.; Kasimir, G.; Perez, U.; Bosch, A.; Pinto, R.; Saucedo, G.; Huguet, J.M.; Figueras, M.J. Occurrence and Diversity of Arcobacter Spp. along the Llobregat River Catchment, at Sewage Effluents and in a Drinking Water Treatment Plant. Water Res. 2010, 44, 3696–3702. [Google Scholar] [CrossRef]
- Webb, A.L.; Taboada, E.N.; Selinger, L.B.; Boras, V.F.; Inglis, G.D. Prevalence and Diversity of Waterborne Arcobacter butzleri in Southwestern Alberta, Canada. Can. J. Microbiol. 2017, 63, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Maruyama, S.; Kabeya, H.; Boonmar, S.; Nimsuphan, B.; Nagai, A.; Kozawa, K.; Nakajima, T.; Mikami, T.; Kimura, H. Isolation and Phylogenetic Analysis of Arcobacter Spp. in Ground Chicken Meat and Environmental Water in Japan and Thailand. Microbiol. Immunol. 2004, 48, 527–533. [Google Scholar] [CrossRef]
- Musmanno, R.A.; Russi, M.; Lior, H.; Figura, N. In Vitro Virulence Factors of Arcobacter butzleri Strains Isolated from Superficial Water Samples. New Microbiol. 1997, 20, 63–68. [Google Scholar]
- Rice, E.W.; Rodgers, M.R.; Wesley, I.V.; Johnson, C.H.; Tanner, S.A. Isolation of Arcobacter butzleri from Ground Water. Lett. Appl. Microbiol. 1999, 28, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Fong, T.-T.; Mansfield, L.S.; Wilson, D.L.; Schwab, D.J.; Molloy, S.L.; Rose, J.B. Massive Microbiological Groundwater Contamination Associated with a Waterborne Outbreak in Lake Erie, South Bass Island, Ohio. Environ. Health Perspect 2007, 115, 856–864. [Google Scholar] [CrossRef] [Green Version]
- Collado, L.; Inza, I.; Guarro, J.; Figueras, M.J. Presence of Arcobacter Spp. in Environmental Waters Correlates with High Levels of Fecal Pollution. Environ. Microbiol. 2008, 10, 1635–1640. [Google Scholar] [CrossRef]
- Merga, J.Y.; Royden, A.; Pandey, A.K.; Williams, N.J. Arcobacter Spp. Isolated from Untreated Domestic Effluent. Lett. Appl. Microbiol. 2014, 59, 122–126. [Google Scholar] [CrossRef]
- Talay, F.; Molva, C.; Atabay, H.I. Isolation and Identification of Arcobacter Species from Environmental and Drinking Water Samples. Folia Microbiol. 2016, 61, 479–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, J.; Lior, H.; Feuerpfeil, I. Isolation of Arcobacter butzleri from a Drinking Water Reservoir in Eastern Germany. Zent. Hyg. Umw. 1993, 193, 557–562. [Google Scholar]
- Fera, M.T.; Maugeri, T.L.; Gugliandolo, C.; Beninati, C.; Giannone, M.; La Camera, E.; Carbone, M. Detection of Arcobacter Spp. in the Coastal Environment of the Mediterranean Sea. Appl. Environ. Microbiol. 2004, 70, 1271–1276. [Google Scholar] [CrossRef] [Green Version]
- Ghane, F.G.M.M. Isolation of Arcobacter butzleri from Caspian Sea’s Water. J. Appl. Environ. Microbiol. 2014, 2, 61–64. [Google Scholar] [CrossRef]
- Noto, A.M.D.; Sciortino, S.; Cardamone, C.; Ciravolo, C.; Napoli, C.; Alio, V.; Arculeo, P.; Oliveri, G.; Costa, A. Detection of Arcobacter spp. in Food Products Collected from Sicilia Region: A Preliminary Study. Ital. J. Food Saf. 2018, 7. [Google Scholar] [CrossRef]
- Collado, L.; Guarro, J.; Figueras, M.J. Prevalence of Arcobacter in Meat and Shellfish. J. Food Prot. 2009, 72, 1102–1106. [Google Scholar] [CrossRef] [PubMed]
- Mottola, A.; Bonerba, E.; Bozzo, G.; Marchetti, P.; Celano, G.V.; Colao, V.; Terio, V.; Tantillo, G.; Figueras, M.J.; Di Pinto, A. Occurrence of Emerging Food-Borne Pathogenic Arcobacter spp. Isolated from Pre-Cut (Ready-to-Eat) Vegetables. Int. J. Food Microbiol. 2016, 236, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Parisi, A.; Capozzi, L.; Bianco, A.; Caruso, M.; Latorre, L.; Costa, A.; Giannico, A.; Ridolfi, D.; Bulzacchelli, C.; Santagada, G. Identification of Virulence and Antibiotic Resistance Factors in Arcobacter butzleri Isolated from Bovine Milk by Whole Genome Sequencing. Ital. J. Food Saf. 2019, 8, 7840. [Google Scholar] [CrossRef] [Green Version]
- Collado, L.; Figueras, M.J. Taxonomy, Epidemiology, and Clinical Relevance of the Genus Arcobacter. Clin. Microbiol. Rev. 2011, 24, 174–192. [Google Scholar] [CrossRef] [Green Version]
- Figueras, M.J.; Levican, A.; Pujol, I.; Ballester, F.; Rabada Quilez, M.J.; Gomez-Bertomeu, F. A Severe Case of Persistent Diarrhoea Associated with Arcobacter cryaerophilus but Attributed to Campylobacter Sp. and a Review of the Clinical Incidence of Arcobacter Spp. New Microbes New Infect. 2014, 2, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Skovgaard, N. Microorganisms in Foods 7: Microbiological Testing in Food Safety Management—By International Commission for the Microbiological Specifications of Foods (ICMSF). Kluwer Academic/Plenum Publishers, New York, Xiii+367 Pages, Hardbound, ISBN 0-306-47262-7, Euro 144.50/ US$ 125.00/GP@$ 88.50. Http://Www.Wkap.Nl. Int. J. Food Microbiol. 2003, 2–3, 291–292. [Google Scholar] [CrossRef]
- Lerner, J.; Brumberger, V.; Preac-Mursic, V. Severe Diarrhea Associated With Arcobacter butzleri. Eur. J. Clin. Microbiol. Infect. Dis. 1994, 13, 660–662. [Google Scholar] [CrossRef]
- Vandenberg, O.; Dediste, A.; Houf, K.; Ibekwem, S.; Souayah, H.; Cadranel, S.; Douat, N.; Zissis, G.; Butzler, J.-P.; Vandamme, P. Arcobacter Species in Humans. Emerg. Infect. Dis. 2004, 10, 1863–1867. [Google Scholar] [CrossRef] [PubMed]
- Arguello, E.; Otto, C.C.; Mead, P.; Babady, N.E. Bacteremia Caused by Arcobacter butzleri in an Immunocompromised Host. J. Clin. Microbiol. 2015, 53, 1448–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandamme, P.; Pugina, P.; Benzi, G.; Etterijck, R.V.; Vlaes, L.; Kersters, K.; Butzler, J.P.; Lior, H.; Lauwers, S. Outbreak of Recurrent Abdominal Cramps Associated with Arcobacter butzleri in an Italian School. J. Clin. Microbiol. 1992, 30, 2335–2337. [Google Scholar] [CrossRef] [Green Version]
- Chieffi, D.; Fanelli, F.; Fusco, V. Arcobacter butzleri: Up-to-Date Taxonomy, Ecology, and Pathogenicity of an Emerging Pathogen. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2071–2109. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Veuthey, B.; Jara, R.; Santander, K.; Mella, A.; Ruiz, S.; Collado, L. Antimicrobial Resistance and Virulence Genes Profiles of Arcobacter butzleri Strains Isolated from Back Yard Chickens and Retail Poultry Meat in Chile. Lett. Appl. Microbiol. 2020, 72, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.; Queiroz, J.A.; Oleastro, M.; Domingues, F.C. Insights in the Pathogenesis and Resistance of Arcobacter: A Review. Crit. Rev. Microbiol. 2016, 42, 364–383. [Google Scholar] [CrossRef] [PubMed]
- Abdelbaqi, K.; Ménard, A.; Prouzet-Mauleon, V.; Bringaud, F.; Lehours, P.; Mégraud, F. Nucleotide Sequence of the GyrA Gene of Arcobacter Species and Characterization of Human Ciprofloxacin-Resistant Clinical Isolates. FEMS Immunol. Med. Microbiol. 2007, 49, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Abeele, A.-M.; Vogelaers, D.; Vanlaere, E.; Houf, K. Antimicrobial Susceptibility Testing of Arcobacter butzleri and Arcobacter cryaerophilus Strains Isolated from Belgian Patients. J. Antimicrob. Chemother. 2016, 71, 1241–1244. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, S.; Luís, Â.; Oleastro, M.; Pereira, L.; Domingues, F.C. A Meta-Analytic Perspective on Arcobacter Spp. Antibiotic Resistance. J. Glob. Antimicrob. Resist. 2019, 16, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Vitale, M.; Galluzzo, P.; Buffa, P.G.; Carlino, E.; Spezia, O.; Alduina, R. Comparison of Antibiotic Resistance Profile and Biofilm Production of Staphylococcus aureus Isolates Derived from Human Specimens and Animal-Derived Samples. Antibiotics (Basel) 2019, 8, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sucato, A.; Vecchioni, L.; Savoca, D.; Presentato, A.; Arculeo, M.; Alduina, R. A Comparative Analysis of Aquatic and Polyethylene-Associated Antibiotic-Resistant Microbiota in the Mediterranean Sea. Biology 2021, 10, 200. [Google Scholar] [CrossRef]
- Blasi, M.F.; Migliore, L.; Mattei, D.; Rotini, A.; Thaller, M.C.; Alduina, R. Antibiotic Resistance of Gram-Negative Bacteria from Wild Captured Loggerhead Sea Turtles. Antibiotics (Basel) 2020, 9, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alduina, R.; Gambino, D.; Presentato, A.; Gentile, A.; Sucato, A.; Savoca, D.; Filippello, S.; Visconti, G.; Caracappa, G.; Vicari, D.; et al. Is Caretta Caretta a Carrier of Antibiotic Resistance in the Mediterranean Sea? Antibiotics (Basel) 2020, 9, 116. [Google Scholar] [CrossRef] [Green Version]
- Hsu, T.-T.D.; Lee, J. Global Distribution and Prevalence of Arcobacter in Food and Water. Zoonoses Public Health 2015, 62, 579–589. [Google Scholar] [CrossRef]
- Isidro, J.; Ferreira, S.; Pinto, M.; Domingues, F.; Oleastro, M.; Gomes, J.P.; Borges, V. Virulence and Antibiotic Resistance Plasticity of Arcobacter butzleri: Insights on the Genomic Diversity of an Emerging Human Pathogen. Infect. Genet. Evol. 2020, 80, 104213. [Google Scholar] [CrossRef]
- Lee, C.; Agidi, S.; Marion, J.W.; Lee, J. Arcobacter in Lake Erie Beach Waters: An Emerging Gastrointestinal Pathogen Linked with Human-Associated Fecal Contamination. Appl. Environ. Microbiol. 2012, 78, 5511–5519. [Google Scholar] [CrossRef]
- Çelik, E.; Ünver, A. Isolation and Identification of Arcobacter spp. by Multiplex PCR from Water Sources in Kars Region. Curr. Microbiol. 2015, 71, 546–550. [Google Scholar] [CrossRef]
- Maugeri, T.; Irrera, G.P.; Lentini, V.; Carbone, M.H.; Fera, M.T.; Gugliandolo, C. Detection and Enumeration of Arcobacter spp. in the Coastal Environment of the Straits of Messina (Italy). New Microbiol. 2005, 28, 177–182. [Google Scholar]
- Atabay, H.I.; Wainø, M.; Madsen, M. Detection and Diversity of Various Arcobacter Species in Danish Poultry. Int. J. Food Microbiol. 2006, 109, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Atabay, H.I.; Unver, A.; Sahin, M.; Otlu, S.; Elmali, M.; Yaman, H. Isolation of Various Arcobacter Species from Domestic Geese (Anser Anser). Vet. Microbiol. 2008, 128, 400–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertas, N.; Dogruer, Y.; Gonulalan, Z.; Guner, A.; Ulger, I. Prevalence of Arcobacter Species in Drinking Water, Spring Water, and Raw Milk as Determined by Multiplex PCR. J. Food Prot. 2010, 73, 2099–2102. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.H.; Saleha, A.A.; Zunita, Z.; Cheah, Y.K.; Murugaiyah, M.; Korejo, N.A. Genetic Characterization of Arcobacter Isolates from Various Sources. Vet. Microbiol. 2012, 160, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Moreno, Y.; Alonso, J.L.; Botella, S.; Ferrús, M.A.; Hernández, J. Survival and Injury of Arcobacter after Artificial Inoculation into Drinking Water. Res. Microbiol. 2004, 155, 726–730. [Google Scholar] [CrossRef]
- Ferreira, S.; Fraqueza, M.J.; Queiroz, J.A.; Domingues, F.C.; Oleastro, M. Genetic Diversity, Antibiotic Resistance and Biofilm-Forming Ability of Arcobacter butzleri Isolated from Poultry and Environment from a Portuguese Slaughterhouse. Int. J. Food Microbiol. 2013, 162, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Assanta, M.A.; Roy, D.; Lemay, M.J.; Montpetit, D. Attachment of Arcobacter butzleri, a New Waterborne Pathogen, to Water Distribution Pipe Surfaces. J. Food Prot. 2002, 65, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Fera, M.T.; Maugeri, T.L.; Giannone, M.; Gugliandolo, C.; La Camera, E.; Blandino, G.; Carbone, M. In Vitro Susceptibility of Arcobacter butzleri and Arcobacter cryaerophilus to Different Antimicrobial Agents. Int. J. Antimicrob. Agents 2003, 21, 488–491. [Google Scholar] [CrossRef]
- Šilha, D.; Pejchalová, M.; Šilhová, L. Susceptibility to 18 Drugs and Multidrug Resistance of Arcobacter Isolates from Different Sources within the Czech Republic. J. Glob. Antimicrob. Resist. 2017, 9, 74–77. [Google Scholar] [CrossRef]
- Ramees, T.P.; Dhama, K.; Karthik, K.; Rathore, R.S.; Kumar, A.; Saminathan, M.; Tiwari, R.; Malik, Y.S.; Singh, R.K. Arcobacter: An Emerging Food-Borne Zoonotic Pathogen, Its Public Health Concerns and Advances in Diagnosis and Control—A Comprehensive Review. Vet. Q. 2017, 37, 136–161. [Google Scholar] [CrossRef] [Green Version]
- Zambri, M.; Cloutier, M.; Adam, Z.; Lapen, D.R.; Wilkes, G.; Sunohara, M.; Topp, E.; Talbot, G.; Khan, I.U.H. Novel Virulence, Antibiotic Resistance and Toxin Gene-Specific PCR-Based Assays for Rapid Pathogenicity Assessment of Arcobacter faecis and Arcobacter lanthieri. BMC Microbiol. 2019, 19, 11. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.Y.; Byelashov, O.A.B.A.; Geornaras, I.G.; Goodridge, L.D.G.D.; Nightingale, K.K.N.K.; Belk, K.E.B.E.; Smith, G.C.S.C.; Sofos, J.N.S.N. Presence of Antibiotic-Resistant Commensal Bacteria in Samples from Agricultural, City, and National Park Environments Evaluated by Standard Culture and Real-Time PCR Methods. Can. J. Microbiol. 2010, 56, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Schnappinger, D.; Hillen, W. Tetracyclines: Antibiotic Action, Uptake, and Resistance Mechanisms. Arch. Microbiol. 1996, 165, 359–369. [Google Scholar] [CrossRef]
- Roberts, M.C. Update on Acquired Tetracycline Resistance Genes. FEMS Microbiol. Lett. 2005, 245, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B.; McMurry, L.M.; Barbosa, T.M.; Burdett, V.; Courvalin, P.; Hillen, W.; Roberts, M.C.; Rood, J.I.; Taylor, D.E. Nomenclature for New Tetracycline Resistance Determinants. Antimicrob. Agents Chemother. 1999, 43, 1523–1524. [Google Scholar] [CrossRef] [Green Version]
- Billington, S.J.; Songer, J.G.; Jost, B.H. Widespread Distribution of a Tet W Determinant among Tetracycline-Resistant Isolates of the Animal Pathogen Arcanobacterium pyogenes. Antimicrob. Agents Chemother. 2002, 46, 1281–1287. [Google Scholar] [CrossRef] [Green Version]
- Chopra, I.; Roberts, M. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [Green Version]
- Houf, K.; De Zutter, L.; Van Hoof, J.; Vandamme, P. Occurrence and Distribution of Arcobacter Species in Poultry Processing. J. Food Prot. 2002, 65, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Hausdorf, L.; Neumann, M.; Bergmann, I.; Sobiella, K.; Mundt, K.; Fröhling, A.; Schlüter, O.; Klocke, M. Occurrence and Genetic Diversity of Arcobacter Spp. in a Spinach-Processing Plant and Evaluation of Two Arcobacter-Specific Quantitative PCR Assays. Syst. Appl. Microbiol. 2013, 36, 235–243. [Google Scholar] [CrossRef]
- van Driessche, E.; Houf, K.; van Hoof, J.; De Zutter, L.; Vandamme, P. Isolation of Arcobacter Species from Animal Feces. FEMS Microbiol. Lett. 2003, 229, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Chinivasagam, H.N.; Corney, B.G.; Wright, L.L.; Diallo, I.S.; Blackall, P.J. Detection of Arcobacter Spp. in Piggery Effluent and Effluent-Irrigated Soils in Southeast Queensland. J. Appl. Microbiol. 2007, 103, 418–426. [Google Scholar] [CrossRef]
- Wesley, I.V. Helicobacter and Arcobacter Species: Risks for Foods and Beverages. J. Food Prot. 1996, 59, 1127–1132. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, A.; Eckert, E.M.; D’Urso, S.; Bertoni, R.; Gillan, D.C.; Wattiez, R.; Corno, G. Co-Occurrence of Integrase 1, Antibiotic and Heavy Metal Resistance Genes in Municipal Wastewater Treatment Plants. Water Res. 2016, 94, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Houf, K.; Tutenel, A.; De Zutter, L.; Van Hoof, J.; Vandamme, P. Development of a Multiplex PCR Assay for the Simultaneous Detection and Identification of Arcobacter butzleri, Arcobacter cryaerophilus and Arcobacter skirrowii. FEMS Microbiol. Lett. 2000, 193, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. Partitionfinder: Combined Selection of Partitioning Schemes and Substitution Models for Phylogenetic Analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.; Gorley, R.N. PRIMER v6: User Manual/Tutorial (Plymouth Routines in Multivariate Ecological Research); Primer-e Ltd: Plymouth, MA, USA, 2006. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample (Source) | N. of Samples | Arcobacter spp. (%) | A. butzleri | A. cryaerophilus |
|---|---|---|---|---|
| Rivers | 11 | 9 (81.8) | 9 | |
| Ponds | 8 | 6 (75) | 5 | 1 |
| Streams | 5 | 2 (40) | 2 | |
| Well water | 20 | 2 (10) | 2 | |
| Spring water | 17 | 1 (5.8) | 1 | |
| Drinking water | 12 | 0 | - | |
| Seawater | 21 | 3 (14.2) | 3 | |
| Seaweeds | 6 | 5 (83.3) | 5 | |
| Total | 100 | 28 (28%) | 27 (96.4%) | 1 (3.6%) |
| Sample Number | Area of Origin | Type of Sample | Arcobacter spp. |
|---|---|---|---|
| 1 | Messina | Seawater | ND |
| Seaweed | A. butzleri | ||
| 2 | Palermo | Seawater | ND |
| Seaweed | ND | ||
| 3 | Palermo | Seawater | A. butzleri |
| Seaweed | A. butzleri | ||
| 4 | Messina | Seawater | ND |
| Seaweed | A. butzleri | ||
| 5 | Palermo | Seawater | A. butzleri |
| Seaweed | A. butzleri | ||
| 6 | Messina | Seawater | A. butzleri |
| Seaweed | A. butzleri |
| Antibiotics | Isolates from | Total (n = 28) | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rivers (n = 9) | Streams (n = 2) | Ponds (n = 6) | Well Waters (n = 2) | Spring Waer (n = 1) | Seawater (n = 3) | Seaweed (n = 5) | |||||||||||||||||||||
| AB (n = 5) | AC (n = 1) | ||||||||||||||||||||||||||
| R | I | S | R | I | S | R | I | S | R | I | S | R | I | S | R | I | S | R | I | S | R | I | S | R | I | S | |
| Amoxicillin-clavulanic acid (AMC) | 7 | 0 | 2 | 2 | 0 | 0 | 5 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 5 | 0 | 0 | 26 | 0 | 2 |
| Ampicillin (AMP) | 9 | 0 | 0 | 2 | 0 | 0 | 5 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 5 | 0 | 0 | 28 | 0 | 0 |
| Cefalotin (KF) | 9 | 0 | 0 | 2 | 0 | 0 | 5 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 5 | 0 | 0 | 28 | 0 | 0 |
| Cefotaxime (CTX) | 9 | 0 | 0 | 2 | 0 | 0 | 5 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 5 | 0 | 0 | 28 | 0 | 0 |
| Ciprofloxacin (CIP) | 1 | 0 | 8 | 0 | 0 | 2 | 0 | 0 | 5 | 1 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 5 | 2 | 0 | 26 |
| Erythromycin (E) | 0 | 2 | 7 | 0 | 0 | 2 | 0 | 0 | 5 | 1 | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 1 | 0 | 2 | 1 | 0 | 4 | 3 | 3 | 22 |
| Gentamycin (CN) | 0 | 0 | 9 | 0 | 0 | 2 | 0 | 0 | 5 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 5 | 0 | 0 | 28 |
| Nalidixic acid (NA) | 9 | 0 | 0 | 2 | 0 | 0 | 5 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 5 | 0 | 0 | 28 | 0 | 0 |
| Streptomycin (S) | 0 | 0 | 9 | 0 | 0 | 2 | 0 | 0 | 5 | 1 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 5 | 1 | 0 | 27 |
| Tetracycline (TE) | 9 | 0 | 0 | 2 | 0 | 0 | 5 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 | 5 | 0 | 0 | 28 | 0 | 0 |
| Sample | Specie | tet Resistance Genes |
|---|---|---|
| Seawater | A. butzleri | tetW−, tetO+, tetA− |
| River | A. butzleri | tetW+, tetO+, tetA− |
| Pond with aquatic animals | A. butzleri | tetW+, tetO−, tetA− |
| River | A. butzleri | tetW+, tetO+, tetA− |
| River | A. butzleri | tetW+, tetO+, tetA− |
| Seawater | A. butzleri | tetW+, tetO+, tetA− |
| River | A. butzleri | tetW+, tetO−, tetA− |
| Pond with turtles | A. butzleri | tetW+, tetO−, tetA− |
| Pond with aquatic animals | A. cryaerophilus | tetW+, tetO−, tetA− |
| Target Name | Primer Sequence (5′-3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|
| 16s rDNA | cggtgaatacgttcycgg | 142 | [63] |
| gghtaccttgttacgactt | |||
| tetA | gctacatcctgcttgccttc | 210 | [64] |
| catagatcgccgtgaagagg | |||
| tetW | acatcattgatactccaggtcacg | 120 | [51] |
| tttcactttgtggttgaacccctc | |||
| tetO | ggaggggttcaaccacaaag | 88 | [51] |
| ctatgtaaataaaatggatag | |||
| gyrA | tggattaaagccagttcatagaag | 344 | [29] |
| tcatmgwatcatcataatttggwac | |||
| qnrS | gacgtgctaacttgcgtgat | 118 | [59] |
| tggcattgttggaaacttg | |||
| butz | cctggacttgacatagtaagaatga | 401 | [64] |
| arco | cgtattcaccgtagcatagc | ||
| skir | ggcgatttactggaacaca | 641 | [64] |
| arco | cgtattcaccgtagcatagc | ||
| cry1 | tgctggagcggatagaagta | 257 | [64] |
| cry2 | aacaacctacgtccttcgac |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sciortino, S.; Arculeo, P.; Alio, V.; Cardamone, C.; Nicastro, L.; Arculeo, M.; Alduina, R.; Costa, A. Occurrence and Antimicrobial Resistance of Arcobacter spp. Recovered from Aquatic Environments. Antibiotics 2021, 10, 288. https://doi.org/10.3390/antibiotics10030288
Sciortino S, Arculeo P, Alio V, Cardamone C, Nicastro L, Arculeo M, Alduina R, Costa A. Occurrence and Antimicrobial Resistance of Arcobacter spp. Recovered from Aquatic Environments. Antibiotics. 2021; 10(3):288. https://doi.org/10.3390/antibiotics10030288
Chicago/Turabian StyleSciortino, Sonia, Pietro Arculeo, Vincenzina Alio, Cinzia Cardamone, Luisa Nicastro, Marco Arculeo, Rosa Alduina, and Antonella Costa. 2021. "Occurrence and Antimicrobial Resistance of Arcobacter spp. Recovered from Aquatic Environments" Antibiotics 10, no. 3: 288. https://doi.org/10.3390/antibiotics10030288
APA StyleSciortino, S., Arculeo, P., Alio, V., Cardamone, C., Nicastro, L., Arculeo, M., Alduina, R., & Costa, A. (2021). Occurrence and Antimicrobial Resistance of Arcobacter spp. Recovered from Aquatic Environments. Antibiotics, 10(3), 288. https://doi.org/10.3390/antibiotics10030288