
Antimicrobial Resistance Profile and ExPEC Virulence Potential in Commensal Escherichia coli of Multiple Sources

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
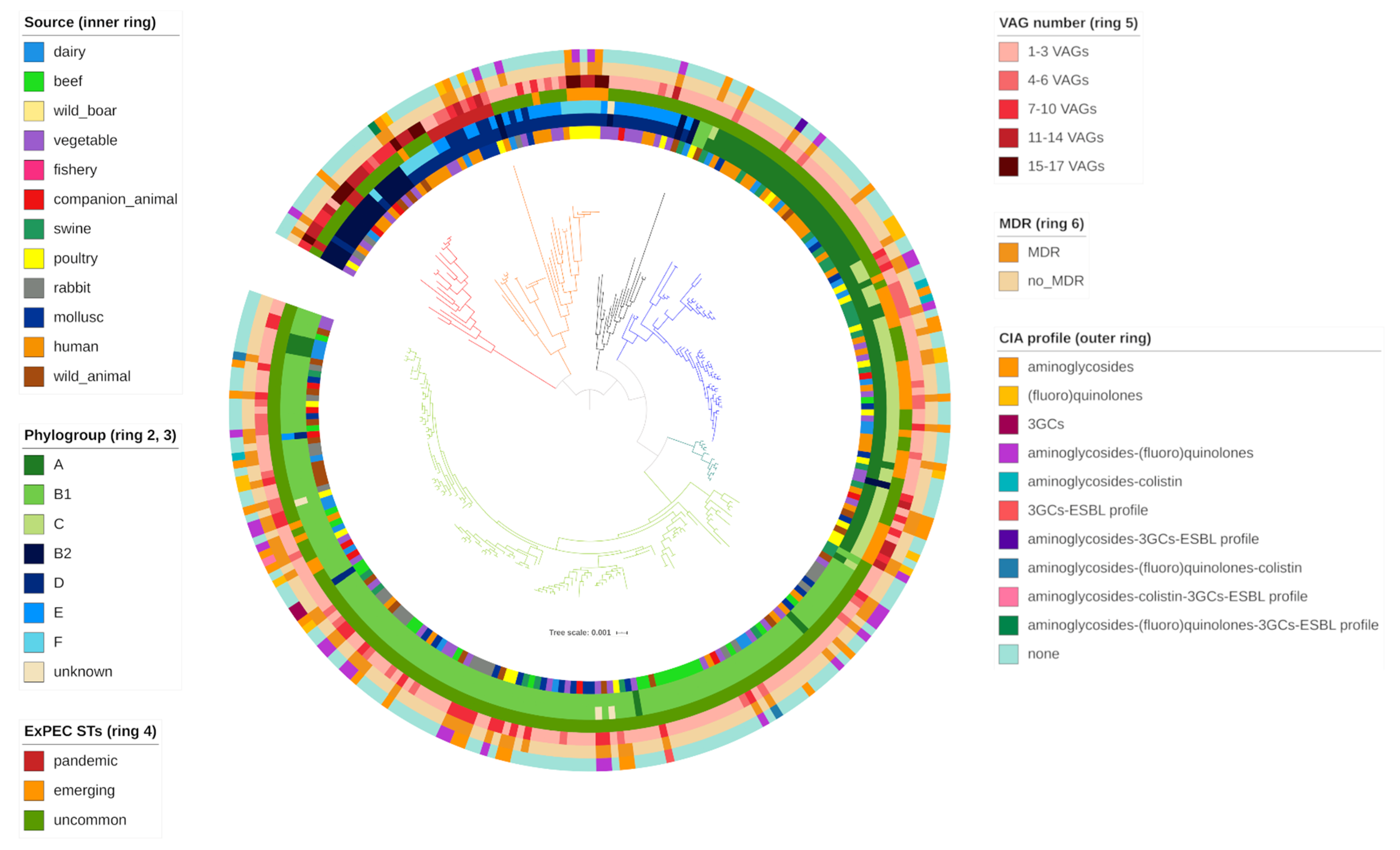
2. Results
2.1. Occurrence of E. coli Strains in Animal, Food and Human Samples
2.2. Antimicrobial Resistance Phenotypes
2.3. Phylogroups
2.4. Expec Virulence Potential
2.5. Statistical Results
3. Discussion
3.1. Tetracycline, Sulfonamide, Streptomycin and Ampicillin Resistance Is the Most Common among the Overall Collection
3.2. The Potential Role of Aquaculture, Vegetable and Wildlife as AMR Source Sentinels
3.3. Phenotypic Q Resistance Is the Most Common Among HP-Cias
3.4. Poultry and Rabbit Are the Sources Mainly Associated to AMR
3.5. Phenotypic Pattern Is Generally Concordant with Genotypic AMR Profile
3.6. Important Phylogroup Variability Occurred among the Different Sources
3.7. Clermont Quadruplex PCR Is a Valid Alternative to In Silico Phylotyping Technique
3.8. Commensal E. coli Conceal Potential Multiresistant Pandemic and Emerging Expec Pathogens
3.9. E. coli Indicator Are Expec VAG Reservoir
3.10. Pandemic and Emerging Expec STs Mainly Belonged to Poultry Source
4. Materials and Methods
4.1. Samples Collection
4.2. Genetic Features of the Collection
4.3. Bacterial Isolation and Molecular Identification
4.4. Antimicrobial Resistance
4.5. Phylogrouping
4.6. Expec Virulence Potential
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. Genet. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Poolman, J.T.; Wacker, M. Extraintestinal Pathogenic Escherichia coli, a Common Human Pathogen: Challenges for Vaccine Development and Progress in the Field. J. Infect. Dis. 2016, 213, 6–13. [Google Scholar] [CrossRef] [Green Version]
- McNulty, C.A.M.; Lecky, D.M.; Xu-McCrae, L.; Nakiboneka-Ssenabulya, D.; Chung, K.-T.; Nichols, T.; Thomas, H.L.; Thomas, M.; Alvarez-Buylla, A.; Turner, K.; et al. CTX-M ESBL-producing Enterobacteriaceae: Estimated prevalence in adults in England in 2014. J. Antimicrob. Chemother. 2018, 73, 1368–1388. [Google Scholar] [CrossRef]
- Rodríguez-Baño, J.; López-Cerero, L.; Navarro, M.D.; De Alba, P.D.; Pascual, A. Faecal carriage of extended-spectrum β-lactamase-producing Escherichia coli: Prevalence, risk factors and molecular epidemiology. J. Antimicrob. Chemother. 2008, 62, 1142–1149. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and Simple Determination of the Escherichia coli Phylogenetic Group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- WHO. Critically Important Antimicrobials for Human Medicine, 6th Revision; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- ECDC. Surveillance Report. Surveillance of Antimicrobial Resistance in Europe 2017; ECDC: Solna, Sweden, 2018. [Google Scholar]
- Johnson, J.R.; Russo, T.A. Extraintestinal pathogenic Escherichia coli: “The other bad E. coli”. J. Lab. Clin. Med. 2002, 139, 155–162. [Google Scholar] [CrossRef]
- Johnson, J.R.; Kuskowski, M.A.; O’Bryan, T.T.; Maslow, J.N. Epidemiological Correlates of Virulence Genotype and Phylogenetic Background among Escherichia coli Blood Isolates from Adults with Diverse-Source Bacteremia. J. Infect. Dis. 2002, 185, 1439–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhler, C.-D.; Dobrindt, U. What defines extraintestinal pathogenic Escherichia coli? Int. J. Med. Microbiol. 2011, 301, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Olier, M.; Hoede, C.; Diancourt, L.; Brisse, S.; Keroudean, M.; Glodt, J.; Picard, B.; Oswald, E.; Denamur, E. Animal and human pathogenic Escherichia coli strains share common genetic backgrounds. Infect. Genet. Evol. 2011, 11, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Owens, K.; Gajewski, A.; Clabots, C. Escherichia coli Colonization Patterns among Human Household Members and Pets, with Attention to Acute Urinary Tract Infection. J. Infect. Dis. 2008, 197, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The population genetics of commensal Escherichia coli. Nat. Rev. Microbiol. Genet. 2010, 8, 207–217. [Google Scholar] [CrossRef]
- Jang, J.; Hur, H.-G.; Sadowsky, M.; Byappanahalli, M.; Yan, T.; Ishii, S. Environmental Escherichia coli: Ecology and public health implications—A review. J. Appl. Microbiol. 2017, 123, 570–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J. 2019, 17, e05598. [Google Scholar]
- Fuhrmeister, A.S.; Jones, R.N. The Importance of Antimicrobial Resistance Monitoring Worldwide and the Origins of SENTRY An-timicrobial Surveillance Program. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Tacconelli, E.; Sifakis, F.; Harbarth, S.; Schrijver, R.; van Mourik, M.; Voss, A.; Sharland, M.; Rajendran, N.B.; Rodríguez-Baño, J.; Bielicki, J.; et al. Surveillance for control of antimicrobial resistance. Lancet Infect. Dis. 2018, 18, e99–e106. [Google Scholar] [CrossRef] [Green Version]
- ECDC/EFSA/EMA. Second joint report on the integrated analysis of the consumption of antimicrobial agents and occur-rence of antimicrobial resistance in bacteria from humans and food-producing animals. EFSA J. 2017, 15, 4872. [Google Scholar]
- Seiffert, S.N.; Tinguely, R.; Lupo, A.; Neuwirth, C.; Perreten, V.; Endimiani, A. High prevalence of extend-ed-spectrum-cephalosporin-resistant Enterobacteriaceae in poultry meat in Switzerland: Emergence of CMY-2-and VEB-6-possessing Proteus mirabilis. Antimicrob. Agents Chemother. 2013, 57, 6406–6408. [Google Scholar] [CrossRef] [Green Version]
- Blaak, H.; De Kruijf, P.; Hamidjaja, R.A.; Van Hoek, A.H.; de Roda Husman, A.M.; Schets, F.M. Prevalence and characteristics of ESBL-producing E. coli in Dutch recreational waters influenced by wastewater treatment plants. Vet. Microbiol. 2014, 171, 448–459. [Google Scholar] [CrossRef]
- Dahms, C.; Hübner, N.-O.; Kossow, A.; Mellmann, A.; Dittmann, K.; Kramer, A. Occurrence of ESBL-Producing Escherichia coli in Livestock and Farm Workers in Mecklenburg-Western Pomerania, Germany. PLoS ONE 2015, 10, e0143326. [Google Scholar] [CrossRef]
- Economou, V.; Gousia, P. Agriculture and food animals as a source of antimicrobial-resistant bacteria. Infect. Drug Resist. 2015, 8, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, S. Understanding the contribution of environmental factors in the spread of antimicrobial resistance. Environ. Health Prev. Med. 2015, 20, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; De Schaetzen, M.-A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial Resistance in the Food Chain: A Review. Int. J. Environ. Res. Public Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anaissie, E.J.; Penzak, S.R.; Dignani, M.C. The hospital water supply as a source of nosocomial infections: A plea for action. Arch. Intern. Med. 2002, 162, 1483–1492. [Google Scholar] [CrossRef] [Green Version]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-spectrum beta-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: A global perspective. Clin. Microbiol Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, D.; Wang, J.; Fanning, S.; McMahon, B.J. Antimicrobial Resistance in Wildlife: Implications for Public Health. Zoonoses Public Health 2015, 62, 534–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Schaik, W. The human gut resistome. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madec, J.-Y.; Haenni, M. Antimicrobial resistance plasmid reservoir in food and food-producing animals. Plasmid 2018, 99, 72–81. [Google Scholar] [CrossRef]
- Von Wintersdorff, C.J.H.; Penders, J.; Van Niekerk, J.M.; Mills, N.D.; Majumder, S.; Van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of Antimicrobial Resistance in Microbial Ecosystems through Horizontal Gene Transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [Green Version]
- Guenther, S.; Ewers, C.; Wieler, L.H. Extended-Spectrum Beta-Lactamases Producing E. coli in Wildlife, yet Another Form of Environmental Pollution? Front. Microbiol. 2011, 2, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massella, E.; Reid, C.J.; Cummins, M.L.; Anantanawat, K.; Zingali, T.; Serraino, A.; Piva, S.; Giacometti, F.; Djordjevic, S.P. Snapshot Study of Whole Genome Sequences of Escherichia coli from Healthy Companion Animals, Livestock, Wildlife, Humans and Food in Italy. Antibiotics 2020, 9, 782. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.; Mather, A.E.; Sánchez-Busó, L.; Page, A.J.; Parkhill, J.; Keane, J.A.; Harris, S.R. ARIBA: Rapid antimicrobial re-sistance genotyping directly from sequencing reads. Microb. Genom. 2017, 3, e000131. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Li, G.; Wilking, H.; Kiessling, S.; Alt, K.; Antáo, E.M.; Laturnus, C.; Diehl, I.; Glodde, S.; Homeier, T.; et al. Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related are they? Int. J. Med. Microbiol. 2007, 297, 163–176. [Google Scholar] [CrossRef]
- Dierikx, C.; van der Goot, J.; Fabri, T.; van Essen-Zandbergen, A.; Smith, H.; Mevius, D. Extended-spectrum-β-lactamase-and AmpC-β-lactamase-producing Escherichia coli in Dutch broilers and broiler farmers. J. Antimicrob. Chemother. 2013, 68, 60–67. [Google Scholar] [CrossRef]
- Pletinckx, L.J.; Verhegghe, M.; Crombe, F.; Dewulf, J.; De Bleecker, Y.; Rasschaert, G.; Butaye, P.; Goddeeris, B.M.; De Man, I. Evidence of possible methicillin-resistant Staphylococcus aureus ST398 spread between pigs and other animals and people re-siding on the same farm. Prev. Vet. Med. 2013, 109, 293–303. [Google Scholar] [CrossRef]
- Van Hoek, A.H.; Veenman, C.; Van Overbeek, W.M.; Lynch, G.; De Roda Husman, A.M.; Blaak, H. Prevalence and charac-terization of ESBL- and AmpC-producing Enterobacteriaceae on retail vegetables. Int. J. Food Microbiol. 2015, 204, 1–8. [Google Scholar] [CrossRef]
- Davis, G.S.; Waits, K.; Nordstrom, L.; Weaver, B.; Aziz, M.; Gauld, L.; Grande, H.; Bigler, R.; Horwinski, J.; Porter, S.B.; et al. Intermingled Klebsiella pneumoniae Populations Between Retail Meats and Human Urinary Tract Infections. Clin. Infect. Dis. 2015, 61, 892–899. [Google Scholar] [CrossRef] [Green Version]
- Florez-Cuadrado, D.; Moreno, M.A.; Ugarte-Ruiz, M.; Dominguez, L. Antimicrobial Resistance in the Food Chain in the Eu-ropean Union. Adv. Food Nutr. Res. 2018, 86, 115–136. [Google Scholar]
- Hudson, J.A.; Frewer, L.J.; Jones, G.; Brereton, P.A.; Whittingham, M.J.; Stewart, G. The agri-food chain and antimicrobial resistance: A review. Trends Food Sci. Technol. 2017, 69, 131–147. [Google Scholar] [CrossRef] [Green Version]
- Manges, A. Escherichia coli and urinary tract infections: The role of poultry-meat. Clin. Microbiol. Infect. 2016, 22, 122–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bywater, R.; Silley, P.; Simjee, S. Antimicrobial breakpoints—Definitions and conflicting requirements. Vet. Microbiol. 2006, 118, 158–159. [Google Scholar] [CrossRef] [PubMed]
- Simjee, S.; Silley, P.; Werling, H.O.; Bywater, R. Potential confusion regarding the term ’resistance’ in epidemiological surveys. J. Antimicrob. Chemother. 2007, 61, 228–229. [Google Scholar] [CrossRef] [PubMed]
- EMA. European Surveillance of Veterinary Antimicrobial Consumption (ESVAC). 2019. Available online: https://www.ema.europa.eu/en/veterinary-regulatory/overview/antimicrobial-resistance/european-surveillance-veterinary-antimicrobial-consumption-esvac (accessed on 2 October 2019).
- De Verdier, K.; Nyman, A.; Greko, C.; Bengtsson, B. Antimicrobial resistance and virulence factors in Escherichia coli from swedish dairy calves. Acta Vet. Scand. 2012, 54, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dotto, G.; Giacomelli, M.; Grilli, G.; Ferrazzi, V.; Carattoli, A.; Fortini, D.; Piccirillo, A. High Prevalence of oqxAB in Escherichia coli Isolates from Domestic and Wild Lagomorphs in Italy. Microb. Drug Resist. 2014, 20, 118–123. [Google Scholar] [CrossRef]
- EFSA. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2016. EFSA J. 2018, 16, 5182. [Google Scholar]
- EFSA. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2014. EFSA J. 2016, 14, 4380. [Google Scholar]
- EFSA. The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2017/2018. EFSA J. 2020, 18, 6007. [Google Scholar]
- Skockova, A.; Bogdanovicoa, K.; Kolackova, I.; Karpiskova, R. Antimicrobial-resistant and extended-spectrum be-ta-lactamase-producing Escherichia coli in raw cow’s milk. J. Food Prot. 2015, 78, 72–77. [Google Scholar] [CrossRef]
- EFSA. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2015. EFSA J. 2017, 15, 4694. [Google Scholar]
- Bailey, J.K.; Pinyon, J.L.; Anantham, S.; Hall, R.M. Commensal Escherichia coli of healthy humans: A reservoir for antibi-otic-resistance determinants. J. Med. Microbiol. 2010, 59, 1331–1339. [Google Scholar] [CrossRef]
- Carvalho, A.; Barbosa, A.; Arais, L.; Ribeiro, P.; Carneiro, V.; Cerqueira, A. Resistance patterns, ESBL genes, and genetic re-latedness of Escherichia coli from dogs and owners. Braz. J. Microbiol. 2016, 47, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Costa, D.; Poeta, P.; Sáenz, Y.; Coelho, A.C.; Matos, M.; Vinué, L.; Rodrigues, J.; Torres, C. Prevalence of antimicrobial resistance and resistance genes in faecal Escherichia coli isolates recovered from healthy pets. Vet. Microbiol. 2008, 127, 97–105. [Google Scholar] [CrossRef]
- Grevskott, D.H.; Svanevik, C.S.; Sunde, M.; Wester, A.L.; Lunestad, B.T. Marine Bivalve Mollusks As Possible Indicators of Multidrug-Resistant Escherichia coli and Other Species of the Enterobacteriaceae Family. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Holvoet, K.; Sampers, I.; Callens, B.; Dewulf, J.; Uyttendaele, M. Moderate Prevalence of Antimicrobial Resistance in Escherichia coli Isolates from Lettuce, Irrigation Water, and Soil. Appl. Environ. Microbiol. 2013, 79, 6677–6683. [Google Scholar] [CrossRef] [Green Version]
- Nobili, G.; Franconieri, I.; Basanisi, M.; La Bella, G.; Tozzoli, R.; Caprioli, A.; La Salandra, G. Short communication: Isolation of Shiga toxin-producing Escherichia coli in raw milk and mozzarella cheese in southern Italy. J. Dairy Sci. 2016, 99, 7877–7880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skurnik, D.; Bonnet, D.; Bernède-Bauduin, C.; Michel, R.; Guette, C.; Becker, J.M.; Balaire, C.; Chau, F.; Mohler, J.; Jarlier, V. Characteristics of human intestinal Escherichia coli with changing environments. Environ. Microbiol. 2008, 10, 2132–2137. [Google Scholar] [CrossRef] [PubMed]
- ECDC. Antimicrobial Consumption. Annual Epidemiological Report for 2017; ECDC: Solna, Sweden, 2018. [Google Scholar]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef] [Green Version]
- Pezzella, C.; Ricci, A.; Digiannatale, E.; Luzzi, I.; Carattoli, A. Tetracycline and Streptomycin Resistance Genes, Transposons, and Plasmids in Salmonella enterica Isolates from Animals in Italy. Antimicrob. Agents Chemother. 2004, 48, 903–908. [Google Scholar] [CrossRef] [Green Version]
- Reid, C.J.; Chowdhury, P.R.; Djordjevic, S.P. Tn6026 and Tn6029 are found in complex resistance regions mobilised by diverse plasmids and chromosomal islands in multiple antibiotic resistant Enterobacteriaceae. Plasmid 2015, 80, 127–137. [Google Scholar] [CrossRef]
- Silver, L.L. Challenges of Antibacterial Discovery. Clin. Microbiol. Rev. 2011, 24, 71–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FVE. Antimicrobial Use in Food-Producing Animals. Replies to EFSA/EMA Questions on the Use of Antimicrobials in Food-Producing Animals in EU and Possible Measures to Reduce Antimicrobial Use. 2016. Available online: https://www.ema.europa.eu/en/documents/report/annex-replies-efsa/ema-questions-use-antimicrobials-food-producing-animals-eu-possible-measures-reduce-antimicrobial_en.pdf (accessed on 30 June 2020).
- Federica, G.; Pezzi, A.; Galletti, G.; Tamba, M.; Merialdi, G.; Piva, S.; Serraino, A.; Rubini, S. Antimicrobial resistance patterns in Salmonella enterica subsp. enterica and Escherichia coli isolated from bivalve molluscs and marine environment. Food Control. 2021, 121, 107590. [Google Scholar]
- Walczak, J.J.; Xu, S. Manure as a Source of Antibiotic-Resistant Escherichia coli and Enterococci: A Case Study of a Wisconsin, USA Family Dairy Farm. Water Air Soil Pollut. 2011, 219, 579–589. [Google Scholar] [CrossRef]
- Alves, M.S.; Pereira, A.; Araújo, S.M.; Castro, B.B.; Correia, A.C.M.; Henriques, I. Seawater is a reservoir of multi-resistant Escherichia coli, including strains hosting plasmid-mediated quinolones resistance and extended-spectrum beta-lactamases genes. Front. Microbiol. 2014, 5, 426. [Google Scholar] [CrossRef]
- Baliere, C.; Rince, A.; Blanco, J.; Dahbi, G.; Harel, J.; Vogeleer, P.; Giard, J.C.; Mariani-Kurkdjian, P.; Gourmelon, M. Prevalence and Characterization of Shiga Toxin-Producing and Enteropathogenic Escherichia coli in Shellfish-Harvesting Areas and Their Watersheds. Front. Microbiol. 2015, 6, 1356. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.L. Environmental pollution by antibiotics and by antibiotic resistance determinants. Environ. Pollut. 2009, 157, 2893–2902. [Google Scholar] [CrossRef] [PubMed]
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Microbiol. Genet. 2010, 8, 251–259. [Google Scholar] [CrossRef]
- Skurnik, D.; Ruimy, R.; Andremont, A.; Amorin, C.; Rouquet, P.; Picard, B.; Denamur, E. Effect of human vicinity on antimi-crobial resistance and integrons in animal faecal Escherichia coli. J. Antimicrob Chemother. 2006, 57, 1215–1219. [Google Scholar] [CrossRef]
- Sousa, M.; Torres, C.; Barros, J.; Somalo, S.; Igrejas, G.; Poeta, P. Gilthead Seabream (Sparus aurata) as Carriers of SHV-12 and TEM-52 Extended-Spectrum Beta-Lactamases-Containing Escherichia coli Isolates. Foodborne Pathog. Dis. 2011, 8, 1139–1141. [Google Scholar] [CrossRef]
- Campos, J.; Mourao, J.; Pestana, N.; Peixe, L.; Novais, C.; Antunes, P. Microbiological quality of ready-to-eat salads: An un-derestimated vehicle of bacteria and clinically relevant antibiotic resistance genes. Int J. Food Microbiol. 2013, 166, 464–470. [Google Scholar] [CrossRef]
- Literak, I.; Dolejska, M.; Radimersky, T.; Klimes, J.; Friedman, M.; Aarestrup, F.M.; Hasman, H.; Cizek, A. Antimicrobial-resistant faecal Escherichia coli in wild mammals in central Europe: Multiresistant Escherichia coli producing extended-spectrum beta-lactamases in wild boars. J. Appl. Microbiol. 2010, 108, 1702–1711. [Google Scholar] [CrossRef]
- Giacopello, C.; Foti, M.; Mascetti, A.; Grosso, F.; Ricciardi, D.; Fisichella, V.; Piccolo, F.L. Antimicrobial resistance patterns of Enterobacteriaceae in European wild bird species admitted in a wildlife rescue centre. Vet. Ital. 2016, 52, 139–144. [Google Scholar]
- De Jong, A.; Stephan, B.; Silley, P. Fluoroquinolone resistance of Escherichia coli and Salmonella from healthy livestock and poultry in the EU. J. Appl. Microbiol. 2011, 112, 239–245. [Google Scholar] [CrossRef]
- Hricova, K.; Roderova, M.; Pudova, V.; Hanulik, V.; Halova, D.; Julinkova, P.; Dolejska, M.; Papousek, I.; Bardon, J. Quino-lone-resistant Escherichia coli in Poultry Farming. Cent. Eur. J. Public Health 2017, 25, 163–167. [Google Scholar] [CrossRef]
- Literak, I.; Reitschmied, T.; Bujnakova, D.; Dolejska, M.; Cizek, A.; Bardon, J.; Pokludova, L.; Alexa, P.; Halova, D.; Jamborova, I. Broilers as a source of quinolone-resistant and extraintestinal pathogenic Escherichia coli in the Czech Republic. Microb. Drug Resist. 2013, 19, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.; Poeta, P.; Sáenz, Y.; Vinué, L.; Coelho, A.C.; Matos, M.; Rojo-Bezares, B.; Rodrigues, J.; Torres, C. Mechanisms of Antibiotic Resistance in Escherichia coli Isolates Recovered from Wild Animals. Microb. Drug Resist. 2008, 14, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Guenther, S.; Grobbel, M.; Lübke-Becker, A.; Goedecke, A.; Friedrich, N.D.; Wieler, L.H.; Ewers, C. Antimicrobial resistance profiles of Escherichia coli from common European wild bird species. Veter. Microbiol. 2010, 144, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Logue, C.M.; Johnson, J.R.; Kuskowski, M.A.; Sherwood, J.S.; Barnes, H.J.; Debroy, C.; Wannemuehler, Y.M.; Obata-Yasuoka, M.; Spanjaard, L.; et al. Associations between multidrug resistance, plasmid content, and virulence potential among extraintestinal pathogenic and commensal Escherichia coli from humans and poultry. Foodborne Pathog. Dis. 2012, 9, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Tadesse, D.A.; Zhao, S.; Tong, E.; Ayers, S.; Singh, A.; Bartholomew, M.J.; Mcdermott, P.F. Antimicrobial drug resistance in Escherichia coli from humans and food animals, United States, 1950–2002. J. Emerg. Infect. Dis. 2012, 18, 741. [Google Scholar] [CrossRef]
- Robinson, T.P.; Thornton, P.K.; Franceschini, G.; Kruska, R.L.; Chiozza, F.; Notenbaert, A.M.O.; Cecchi, G.; Herrero, M.; Epprecht, M.; Fritz, S.; et al. Global Livestock Production Systems; FAO: Rome, Italy; ILRI: Nairobi, Kenya, 2011. [Google Scholar]
- Jensen, V.; Jacobsen, E.; Bager, F. Veterinary antimicrobial-usage statistics based on standardized measures of dosage. Prev. Vet. Med. 2004, 64, 201–215. [Google Scholar] [CrossRef]
- Chantziaras, I.; Boyen, F.; Callens, B.; Dewulf, J. Correlation between veterinary antimicrobial use and antimicrobial resistance in food-producing animals: A report on seven countries. J. Antimicrob. Chemother. 2014, 69, 827–834. [Google Scholar] [CrossRef] [Green Version]
- Blanco, J.E.; Blanco, M.; Blanco, J.; Rioja, L.; Ducha, J. Serotypes, toxins and antibiotic resistance of Escherichia coli strains isolated from diarrhoeic and healthy rabbits in Spain. Vet. Microbiol. 1994, 38, 193–201. [Google Scholar] [CrossRef]
- Freitas-Silva, J.; Inácio, Â.S.; Mourão, J.; Antunes, P.; Mendes, Â.; De Carvalho, A.P.; Vasconcelos, V.; Peixe, L.; Da Costa, P.M. Occurrence of mcr-1 in Escherichia coli from rabbits of intensive farming. Veter. Microbiol. 2018, 227, 78–81. [Google Scholar] [CrossRef] [Green Version]
- Medina, A.; De La Fuente, R.; Badiola, I.; De Rozas, A.P.; Orden, J.A.; Ruiz-Santa Quiteria, J.A. Antimicrobial Resistance in Commensal Escherichia coli Isolates from Rabbits. Rev. Cient. 2015, 25, 38–42. [Google Scholar]
- Agnoletti, F.; Brunetta, R.; Bano, L.; Drigo, I.; Mazzolini, E. Longitudinal study on antimicrobial consumption and resistance in rabbit farming. Int. J. Antimicrob. Agents. 2018, 51, 197–205. [Google Scholar] [CrossRef]
- Johnson, J.R.; Kuskowski, M.A.; Menard, M.; Gajewski, A.; Xercavins, M.; Garau, J. Similarity between Human and Chicken Escherichia coli Isolates in Relation to Ciprofloxacin Resistance Status. J. Infect. Dis. 2006, 194, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Kuskowski, M.A.; Smith, K.; O’bryan, T.T.; Tatini, S. Antimicrobial-resistant and extraintestinal pathogenic Escherichia coli in retail foods. J. Infect. Dis. 2005, 191, 1040–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Porter, S.B.; Johnston, B.; Thuras, P.; Clock, S.; Crupain, M.; Rangan, U. Extraintestinal Pathogenic and Anti-microbial-Resistant Escherichia coli, Including Sequence Type 131 (ST131), from Retail Chicken Breasts in the United States in 2013. Appl. Environ. Microbiol. 2017, 83, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Sannes, M.R.; Croy, C.; Johnston, B.; Clabots, C.; Kuskowski, M.A.; Bender, J.; Smith, K.E.; Winokur, P.L.; Be-longia, E.A. Antimicrobial drug-resistant Escherichia coli from humans and poultry products, Minnesota and Wisconsin, 2002–2004. Emerg. Infect. Dis. 2007, 13, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.L.; Wootton, L.; Day, M.R.; Threlfall, E.J. Plasmid-mediated quinolone resistance determinant qnrS1 found in Salmonella enterica strains isolated in the UK. J. Antimicrob. Chemother. 2007, 59, 1071–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, D.C.; Jacoby, G.A. Mechanisms of drug resistance: Quinolone resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 12–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sáenz, Y.; Zarazaga, M.; Briñas, L.; Ruiz-Larrea, F.; Torres, C. Mutations in gyrA and parC genes in nalidixic acid-resistant Escherichia coli strains from food products, humans and animals. J. Antimicrob. Chemother. 2003, 51, 1001–1005. [Google Scholar] [CrossRef]
- Paltansing, S.; Kraakman, M.E.M.; Ras, J.M.C.; Wessels, E.; Bernards, A.T. Characterization of fluoroquinolone and cephalosporin resistance mechanisms in Enterobacteriaceae isolated in a Dutch teaching hospital reveals the presence of an Escherichia coli ST131 clone with a specific mutation in parE. J. Antimicrob. Chemother. 2012, 68, 40–45. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, S.; Kehrenberg, C.; Doublet, B.; Cloeckaert, A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 2004, 28, 519–542. [Google Scholar] [CrossRef] [Green Version]
- Poole, K. Efflux-Mediated Antimicrobial Resistance. J. Antimicrob. Chemother. 2005, 56, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Piddock, L.J.V. Multidrug-resistance efflux pumps? Not just for resistance. Nat. Rev. Microbiol. Genet. 2006, 4, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quesada, A.; Porrero, M.C.; Téllez, S.; Palomo, G.; García, M.; Domínguez, L. Polymorphism of genes encoding PmrAB in colistin-resistant strains of Escherichia coli and Salmonella enterica isolated from poultry and swine. J. Antimicrob. Chemother. 2015, 70, 71–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafique, M.; Potter, R.F.; Ferreiro, A.; Wallace, M.A.; Rahim, A.; Malik, A.A.; Siddique, N.; Abbas, M.A.; D’Souza, A.W.; Burnham, C.-A.D.; et al. Genomic Characterization of Antibiotic Resistant Escherichia coli Isolated From Domestic Chickens in Pakistan. Front. Microbiol. 2020, 10, 3052. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S. The Salmonella PmrAB regulon: Lipopolysaccharide modifications, antimicrobial peptide resistance and more. Trends Microbiol. 2008, 16, 284–290. [Google Scholar] [CrossRef]
- Kempf, I.; Fleury, M.A.; Drider, D.; Bruneau, M.; Sanders, P.; Chauvin, C.; Madec, J.Y.; Jouy, E. What do we know about re-sistance to colistin in Enterobacteriaceae in avian and pig production in Europe? Int. J. Antimicrob. Agents 2013, 42, 379–383. [Google Scholar] [CrossRef]
- Carlos, C.; Pires, M.M.; Stoppe, N.C.; Hachich, E.M.; Sato, M.I.; Gomes, T.A.T.; Amaral, L.A.; Ottoboni, L.M.M. Escherichia coli phylogenetic group determination and its application in the identification of the major animal source of fecal contamination. BMC Microbiol. 2010, 10, 161. [Google Scholar] [CrossRef] [Green Version]
- Escobar-Páramo, P.; Le Menac’H, A.; Le Gall, T.; Amorin, C.; Gouriou, S.; Picard, B.; Skurnik, D.; Denamur, E. Identification of forces shaping the commensal Escherichia coli genetic structure by comparing animal and human isolates. Environ. Microbiol. 2006, 8, 1975–1984. [Google Scholar] [CrossRef]
- Smati, M.; Clermont, O.; Bleibtreu, A.; Fourreau, F.; David, A.; Daubié, A.; Hignard, C.; Loison, O.; Picard, B.; Denamur, E. Quantitative analysis of commensal Escherichia coli populations reveals host-specific enterotypes at the intra-species level. Microbiologyopen 2015, 4, 604–615. [Google Scholar] [CrossRef]
- Alonso, C.A.; González-Barrio, D.; Ruiz-Fons, F.; Ruiz-Ripa, L.; Torres, C. High frequency of B2 phylogroup among non-clonally related fecal Escherichia coli isolates from wild boars, including the lineage ST131. FEMS Microbiol. Ecol. 2017, 93, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, D.M.; Cowling, A. The distribution and genetic structure of Escherichia coli in Australian vertebrates: Host and ge-ographic effects. Microbiology 2003, 149, 3575–3586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoppe, N.D.C.; Silva, J.S.; Carlos, C.; Sato, M.I.Z.; Saraiva, A.M.; Ottoboni, L.M.M.; Torres, T.T. Worldwide Phylogenetic Group Patterns of Escherichia coli from Commensal Human and Wastewater Treatment Plant Isolates. Front. Microbiol. 2017, 8, 2512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsen, L.; Kurbasic, A.; Skjøt-Rasmussen, L.; Ejrnæs, K.; Porsbo, L.J.; Pedersen, K.; Jensen, L.B.; Emborg, H.-D.; Agersø, Y.; Olsen, K.E.; et al. Escherichia coli Isolates from Broiler Chicken Meat, Broiler Chickens, Pork, and Pigs Share Phylogroups and Antimicrobial Resistance with Community-Dwelling Humans and Patients with Urinary Tract Infection. Foodborne Pathog. Dis. 2010, 7, 537–547. [Google Scholar] [CrossRef]
- Ferreira, J.C.; Filho, R.A.C.P.; Kuaye, A.P.Y.; Andrade, L.N.; Chang, Y.-F.; Darini, A.L.C. Virulence potential of commensal multidrug resistant Escherichia coli isolated from poultry in Brazil. Infect. Genet. Evol. 2018, 65, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Smati, M.; Clermont, O.; Le Gal, F.; Schichmanoff, O.; Jauréguy, F.; Eddi, A.; Denamur, E.; Picard, B.; for the Coliville Group. Real-Time PCR for Quantitative Analysis of Human Commensal Escherichia coli Populations Reveals a High Frequency of Subdominant Phylogroups. Appl. Environ. Microbiol. 2013, 79, 5005–5012. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, V.M.; Pinchbeck, G.L.; Nuttall, T.; McEwan, N.; Dawson, S.; Williams, N.J. Antimicrobial resistance risk factors and characterisation of faecal E. coli isolated from healthy Labrador retrievers in the United Kingdom. Prev. Vet. Med. 2015, 119, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Lescat, M.; Clermont, O.; Woerther, P.L.; Glodt, J.; Dion, S.; Skurnik, D.; Djossou, F.; Dupont, C.; Perroz, G.; Picard, B.; et al. Commensal Escherichia coli strains in Guiana reveal a high genetic diversity with host-dependant population structure. Environ. Microbiol. Rep. 2013, 5, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Bok, E.; Mazurek, J.; Stosik, M.; Wojciech, M.; Baldy-Chudzik, K. Prevalence of virulence determinants and antimicrobial re-sistance among commensal Escherichia coli derived from dairy and beef cattle. Int. J. Environ. Res. Public Health 2015, 12, 970–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamelin, K.; Bruant, G.; El-Shaarawi, A.; Hill, S.; Edge, T.A.; Fairbrother, J.; Harel, J.; Maynard, C.; Masson, L.; Brousseau, R. Occurrence of Virulence and Antimicrobial Resistance Genes in Escherichia coli Isolates from Different Aquatic Ecosystems within the St. Clair River and Detroit River Areas. Appl. Environ. Microbiol. 2006, 73, 477–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard, B.; Garcia, J.S.; Gouriou, S.; Duriez, P.; Brahimi, N.; Bingen, E.; Elion, J.; Denamur, E. The link between phylogeny and virulence in Escherichia coli extraintestinal infection. Infect. Immun. 1999, 67, 546–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Kuskowski, M. Clonal origin, virulence factors, and virulence. Infect. Immun. 2000, 68, 424–425. [Google Scholar] [CrossRef] [Green Version]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Schierack, P.; Walk, N.; Ewers, C.; Wilking, H.; Steinrück, H.; Filter, M.; Wieler, L.H. ExPEC-typical virulence-associated genes correlate with successful colonization by intestinal E. coli in a small piglet group. Environ Microbiol. 2008, 10, 1742–1751. [Google Scholar] [CrossRef]
- Los, F.C.O.; Randis, T.M.; Aroian, R.V.; Ratner, A.J. Role of Pore-Forming Toxins in Bacterial Infectious Diseases. Microbiol. Mol. Biol. Rev. 2013, 77, 173–207. [Google Scholar] [CrossRef] [Green Version]
- Sedeek, D.M.; Rady, M.M.; Fedawy, H.S.; Rabie, N.S. Molecular Epidemiology and Sequencing of Avian Pathogenic Escherichia coli APEC in Egypt. Adv. Anim. Vet. Sci. 2020, 8, 499–505. [Google Scholar] [CrossRef]
- Nicolas-Chanoine, M.-H.; Bertrand, X.; Madec, J.-Y. Escherichia coli ST131, an Intriguing Clonal Group. Clin. Microbiol. Rev. 2014, 27, 543–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, L.W. Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin. Microbiol. Infect. 2014, 20, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Dodson, K.W.; Pinkner, J.S.; Rose, T.; Magnusson, G.; Hultgren, S.J.; Waksman, G. Structural Basis of the Interaction of the Pyelonephritic E. coli Adhesin to Its Human Kidney Receptor. Cell 2001, 105, 733–743. [Google Scholar] [CrossRef] [Green Version]
- Lund, B.; Lindberg, F.; Marklund, B.I.; Normark, S. The PapG protein is the α-d-galactopyranosyl-(1→4)-β-d-galactopyranose-binding adhesin of uropathogenic Escherichia coli. Proc. Natl. Acad. Sci. USA 1987, 84, 5898–5902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, M.; Mobley, H. Role of P-fimbrial-mediated adherence in pyelonephritis and persistence of uropathogenic Escherichia coli (UPEC) in the mammalian kidney. Kidney Int. 2007, 72, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katouli, M. Population structure of gut Escherichia coli and its role in development of extra-intestinal infections. Iran. J. Microbiol. 2010, 2, 59–72. [Google Scholar]
- Restieri, C.; Garriss, G.; Locas, M.-C.; Dozois, C.M. Autotransporter-Encoding Sequences Are Phylogenetically Distributed among Escherichia coli Clinical Isolates and Reference Strains. Appl. Environ. Microbiol. 2007, 73, 1553–1562. [Google Scholar] [CrossRef] [Green Version]
- Rijavec, M.; Müller-Premru, M.; Zakotnik, B.; Žgur-Bertok, D. Virulence factors and biofilm production among Escherichia coli strains causing bacteraemia of urinary tract origin. J. Med. Microbiol. 2008, 57, 1329–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, S.; Nakano, M.; Terai, A.; Yuri, K.; Nakata, K.; Nair, G.B.; Kurazono, H.; Ogawa, O. The presence of the virulence island containing the usp gene in uropathogenic Escherichia coli is associated with urinary tract infection in an experimental mouse model. J. Urol. 2001, 165, 1347–1351. [Google Scholar] [CrossRef]
- Johnson, J.R.; Manges, A.R.; O’Bryan, T.T.; Riley, L.W. A disseminated multidrug-resistant clonal group of uropathogenic Escherichia coli in pyelonephritis. Lancet 2002, 359, 2249–2251. [Google Scholar] [CrossRef]
- Villa, L.; García-Fernández, A.; Fortini, D.; Carattoli, A. Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J. Antimicrob. Chemother. 2010, 65, 2518–2529. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Nolan, L.K. Pathogenomics of the virulence plasmids of Escherichia coli. Microbiol. Mol. Biol. Rev. 2009, 73, 750–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, G.J.; Mendonça, N. Association between antimicrobial resistance and virulence in Escherichia coli. Virulence 2012, 3, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura, A.; Oliveira, C.; Henriques, I.; Smalla, K.; Correia, A. Broad diversity of conjugative plasmids in integron-carrying bacteria from wastewater environments. FEMS Microbiol. Lett. 2012, 330, 157–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitout, J.D.D. Extraintestinal pathogenic Escherichia coli: An update on antimicrobial resistance, laboratory diagnosis and treatment. Expert Rev. Anti Infect. Ther. 2012, 10, 1165–1176. [Google Scholar] [CrossRef]
- Bauchart, P.; Germon, P.; Bree, A.; Oswald, E.; Hacker, J.; Dobrindt, U. Pathogenomic comparison of human extraintestinal and avian pathogenic Escherichia coli—Search for factors involved in host specificity or zoonotic potential. Microb. Pathog. 2010, 49, 105–115. [Google Scholar] [CrossRef]
- Li, G.; Cai, W.; Hussein, A.; Wannemuehler, Y.M.; Logue, C.M.; Nolan, L.K. Proteome response of an extraintestinal pathogenic Escherichia coli strain with zoonotic potential to human and chicken sera. J. Proteom. 2012, 75, 4853–4862. [Google Scholar] [CrossRef]
- Vincent, C.; Boerlin, P.; Daignault, D.; Dozois, C.M.; Dutil, L.; Galanakis, C.; Reid-Smith, R.J.; Tellier, P.-P.; Tellis, P.A.; Ziebell, K.; et al. Food Reservoir for Escherichia coli Causing Urinary Tract Infections. Emerg. Infect. Dis. 2010, 16, 88–95. [Google Scholar] [CrossRef]
- Bergeron, C.R.; Prussing, C.; Boerlin, P.; Daignault, D.; Dutil, L.; Reid-Smith, R.J.; Zhanel, G.G.; Manges, A.R. Chicken as res-ervoir for extraintestinal pathogenic Escherichia coli in humans, Canada. Emerg. Infect. Dis. 2012, 18, 415–421. [Google Scholar] [CrossRef]
- Mor-Mur, M.; Yuste, J. Emerging Bacterial Pathogens in Meat and Poultry: An Overview. Food Bioprocess Technol. 2010, 3, 24–35. [Google Scholar] [CrossRef]
- Lyhs, U.; Ikonen, I.; Pohjanvirta, T.; Raninen, K.; Perko-Mäkelä, P.; Pelkonen, S. Extraintestinal pathogenic Escherichia coli in poultry meat products on the Finnish retail market. Acta Vet. Scand. 2012, 54, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, S.M.; Subbiah, M.; Jeffrey, L.; Ullman, J.L.; Frear, C.; Call, D.R. Evaluation of 27 different biochars for potential sequestration of antibiotic residues in food animal production environments. J. Environ. Chem. Eng. 2015, 3, 162–169. [Google Scholar] [CrossRef]
- Horakova, K.; Mlejnkova, H.; Mlejnek, P. Specific detection of Escherichia coli isolated from water samples using polymerase chain reaction targeting four genes: Cytochrome bd complex, lactose permease, β-d-glucuronidase, and β-d-galactosidase. J. Appl. Microbiol. 2008, 105, 970–976. [Google Scholar] [CrossRef]
- CLSI. M100: Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- EUCAST. EUCAST Guideline for the Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance, v2.0.; EUCAST: Växjö, Sweden, 2017. [Google Scholar]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 9.0; EUCAST: Växjö, Sweden, 2019. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Informational Supplement: M31-S1, No. 17; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2004; Volume 24. [Google Scholar]
- CLSI. M100: Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.; Giske, C.; Harbarth, S.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Gordon, D.M.; Brisse, S.; Walk, S.T.; Denamur, E. Characterization of the cryptic Escherichia lineages: Rapid identification and prevalence. Environ. Microbiol. 2011, 13, 2468–2477. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (ExPEC) Lineages. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef] [PubMed]
- Lindstedt, B.-A.; Finton, M.D.; Porcellato, D.; Brandal, L.T. High frequency of hybrid Escherichia coli strains with combined Intestinal Pathogenic Escherichia coli (IPEC) and Extraintestinal Pathogenic Escherichia coli (ExPEC) virulence factors isolated from human faecal samples. BMC Infect. Dis. 2018, 18, 544. [Google Scholar] [CrossRef] [PubMed]
- Kudinha, T.; Kong, F.; Johnson, J.R.; Andrew, S.D.; Anderson, P.; Gilbert, G.L. Multiplex PCR-based reverse line blot assay for simultaneous detection of 22 virulence genes in uropathogenic Escherichia coli. Appl Environ. Microbiol. 2012, 78, 1198–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóth, I.; Nougayrède, J.P.; Dobrindt, U.; Ledger, T.N.; Boury, M.; Morabito, S.; Fujiwara, T.; Sugai, M.; Hacker, J.; Oswald, E. Cytolethal distending toxin type I and type IV genes are framed with lambdoid prophage genes in extraintestinal pathogenic Escherichia coli. Infect Immun. 2009, 77, 492–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostblom, A.; Adlerberth, I.; Wold, A.E.; Nowrouzian, F.L. Pathogenicity island markers, virulence determinants malX and usp, and the capacity of Escherichia coli to persist in infants’ commensal microbiotas. Appl. Environ. Microbiol. 2011, 77, 2303–2308. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; Urbana, University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Source | Number of Samples | Number of Isolated E. coli | E. coli Occurrence (%) |
|---|---|---|---|
| Beef | 33 | 21 | 63.64 |
| Wild Boar | 31 | 22 | 70.97 |
| Vegetable | 164 | 24 | 14.63 |
| Fishery | 94 | 24 | 25.53 |
| Companion Animal | 12 | 12 | 100.00 |
| Swine | 6 | 6 | 100.00 |
| Poultry | 33 | 25 | 75.76 |
| Rabbit | 14 | 10 | 71.43 |
| Human | 27 | 25 | 92.59 |
| HP-CIA | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sources | n | R ≥ 1 | MDR | CN | S | C | ST | SXT | TE | NA | ENR | EFT | CAZ | COL | ESBL |
| Dairy | 25 | 6 (24%) | 6 (24%) | 1 (4%) | 6 (24%) | 3 (12%) | 6 (24%) | 6 (24%) | 6 (24%) | 3 (12%) | 3 (12%) | 2 (8%) | 2 (8%) | 1 (4%) | 2 (8%) |
| Beef | 24 | 7 (29.17%) | 4 (16.66%) | 2 (8.33%) | 5 (20.83%) | 2 (8.33%) | 4 (16.67%) | 4 (16.67%) | 3 (12.5%) | 1 (4.17%) | 1 (4.17%) | 2 (8.33%) | 2 (8.33%) | 0 (0%) | 0 (0%) |
| Wild boar | 22 | 1 (4.55%) | 1 (4.55%) | 0 (0%) | 1 (4.55%) | 0 (0%) | 1 (4.55%) | 1 (4.55%) | 1 (4.55%) | 1 (4.55%) | 1 (4.55%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Vegetable | 24 | 2 (8.33%) | 2 (8.33%) | 0 (0%) | 2 (8.33%) | 0 (0%) | 1 (4.17%) | 1 (4.17%) | 1 (4.55%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Fishery | 24 | 5 (20.83%) | 3 (12.5%) | 1 (4.17%) | 3 (12.5%) | 0 (0%) | 2 (8.33%) | 3 (12.5%) | 3 (12.5%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Companion animal | 12 | 4 (33.33%) | 3 (25%) | 0 (0%) | 1 (8.33%) | 1 (8.33%) | 3 (25%) | 3 (25%) | 4 (33.33%) | 1 (8.33%) | 1 (8.33%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Swine | 25 | 16 (64%) | 6 (24%) | 0 (0%) | 8 (32%) | 2 (8%) | 5 (20%) | 5 (20%) | 10 (40%) | 3 (12%) | 3 (12%) | 0 (0%) | 0 (0%) | 2 (8%) | 0 (0%) |
| Poultry | 25 | 23 (92%) | 16 (64%) | 1 (4%) | 13 (52%) | 8 (32%) | 14 (56%) | 15 (60%) | 13 (52%) | 13 (52%) | 11 (44%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Rabbit | 23 | 23 (100%) | 22 (95.65%) | 6 (26.09%) | 22 (95.65%) | 3 (13.04%) | 22 (95.65%) | 22 (95.65%) | 22 (95.65%) | 15 (65.22%) | 13 (56.52%) | 0 (0%) | 0 (0%) | 3 (13.04%) | 0 (0%) |
| Mollusc | 25 | 4 (16%) | 3 (12%) | 0 (0%) | 2 (8%) | 0 (0%) | 1 (4%) | 1 (4%) | 2 (8%) | 1 (4%) | 1 (4%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Human | 25 | 9 (36%) | 7 (28%) | 0 (0%) | 5 (20%) | 3 (12%) | 6 (24%) | 6 (24%) | 6 (24%) | 6 (24%) | 6 (24%) | 1 (4%) | 1 (4%) | 0 (0%) | 1 (4%) |
| Wild animal | 25 | 7 (28%) | 6 (24%) | 0 (0%) | 3 (12%) | 1 (4%) | 6 (24%) | 6 (24%) | 5 (20%) | 4 (16%) | 1 (4%) | 1 (4%) | 1 (4%) | 0 (0%) | 1 (4%) |
| Total | 279 | 107 (38.35%) | 79 (28.32%) | 11 (3.94%) | 71 (25.45%) | 23 (8.24%) | 71 (25.45) | 73 (26.16%) | 76 (27.24%) | 48 (17.20%) | 44 (15.77%) | 6 (2.15%) | 6 (2.15%) | 6 (2.15%) | 4 (1.43%) |
| Phenotypic Resistance | n | Genetic Determinants |
|---|---|---|
| Beta-lactams | ||
| amipicillin | 60 | blaSHV73, blaTEM-1b (2) |
| blaTEM-1a (4) | ||
| blaTEM-1b (38) | ||
| blaTEM-1c (8) | ||
| blaTEM-1d (4) | ||
| blaTEM-214 (1) | ||
| blaTEM-220 (3) | ||
| ampicillin, ceftiofur, ceftazidime | 6 | ampC# (2) |
| blaCMY-2, blaCTX-M55, blaTEM-1b (1) (1) | ||
| blaCTX-M1 (1) | ||
| blaCTX-M1, blaTEM-1b (1) | ||
| blaCTX-M15, blaTEM-1b (1) | ||
| Chloramphenicol | 23 | catA1 (8) |
| catA1,cmlA1 (2) | ||
| catA2 (1) | ||
| cmlA1 (11) | ||
| mdfA, acrAB-TolC (1) | ||
| Aminoglycosides | ||
| gentamicin | 1 | aac(3)_IIa (1) |
| streptomycin | 61 | aadA1 (16) |
| aadA2 (1) | ||
| aadA2b (1) | ||
| strA (2) | ||
| strB(1) | ||
| aadA1, aadA2 (2) | ||
| strA, strB (20) | ||
| aadA1, aadA2b (6) | ||
| strA, strB, aadA5 (2) | ||
| aadA1, aadA2b, strB (1) | ||
| aadA1, strA, strB (7) | ||
| aadA1, aadA2b, strA, strB (2) | ||
| gentamicin, streptomycin | 10 | aac(3)_IIa, aadA1, aadA2b, strA, strB (1) |
| aac(3)_IIa, aadA1, strA, strB (1) | ||
| ant2_Ia, aadA1 (1) | ||
| aac(3)-IId, aadA5, strA, strB (2) | ||
| aac(3)_IIa, aac3_IV, aadA1, aadA2b, strA, strB (1) | ||
| aac(3)_IV, strA, strB (1) | ||
| aac(3)_IV, aadA1, strA, strB (2) | ||
| aac)(3)_IIa, strA, strB (1) | ||
| Sulphonamides | ||
| sulfisoxazole, sulfametoxazole/trimethoprim | 71 | sul1 (2) |
| sul1, dfrA1 (13) | ||
| sul1, dfrA12 (1) | ||
| sul2 (17) | ||
| sul2, dfrA1 (3) | ||
| sul2, dfrA14 (6) | ||
| sul2, dfrA17 (1) | ||
| sul1, sul2 (1) | ||
| sul1, sul2, dfrA1 (7) | ||
| sul1, sul2, dfrA17 (3) | ||
| sul1, sul2, dfrA7 (1) | ||
| sul1, sul2, dfrA1, dfrA14 (1) | ||
| sul1, sul2, dfrA1, dfrA7 (1) | ||
| sul3 (2) | ||
| sul2, sul3, dfrA14 (2) | ||
| sul3, dfrA1 (3) | ||
| sul3, dfrA12 (2) | ||
| sul3, dfrA14 (1) | ||
| sul1, sul2, sul3, dfrA1 (2) | ||
| sul1, sul3, dfrA1 (2) | ||
| Sulfamethoxazole/trimethoprim | 2 | dfrA1 (1) |
| dfrA12 (1) | ||
| Tetracycline | 76 | tetA (54) |
| tetB (19) | ||
| tetA, tetM (1) | ||
| tetA, tetB (2) | ||
| Colistin | 4 | mcr1 (4) |
| 1 * | pmrB# (1) | |
| ESBL profile | 4 | blaCTX-M1 (2) |
| blaCTX-M15 (1) | ||
| blaCTX-M55 (1) | ||
| (Fluoro)quinolones | ||
| nalidixic acid | 4 | gyrA# (D87G) (3) |
| gyrA# (A84P), parC # (S57T) (1) | ||
| nalidixic acid, enrofloxacin | 45 | gyrA# (S83L) (21) |
| gyrA# (S83L-D87N), parC # (S80I) (13) | ||
| gyrA# (S83L-D87N), parC # (S80I-E84G) (2) | ||
| gyrA# (S83L-D87N), parC # (S80I-E84A) (1) | ||
| gyrA# (S83L-D87N), parC # (S80I-E84G-A56T) (1) | ||
| gyrA# (S83L-D87N), parC # (S80I), pare # (S458A) (3) | ||
| gyrA# (S83L-D87N), parC # (S80I), pare # (L416F) (1) | ||
| gyrA# (S83L-D87Y), parC # (S80I) (1) | ||
| gyrA# (S83L-D87Y), parC # (S80I), parE# (S458A) (1) | ||
| qnrB19, pare# (I355T) (1) | ||
| pare# (I355T) (5) | ||
| 4 * | parC# (A56T)(1) | |
| qnrS1 (3) | ||
| Source | n | A | B1 | B2 | C | D | E | F | unknown | H′ | D |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Dairy | 25 | 8 (32%) | 9 (36%) | 2 (8%) | 4 (16%) | 1 (4%) | 1 (4%) | 0 (0%) | 0 (0%) | 1.485 | 0.763 |
| Beef | 24 | 0 (0%) | 22 (91.67%) | 0 (0%) | 1 (4.17%) | 1 (4.17%) | 0 (0%) | 0 (0%) | 0 (0%) | 0.345 | 0.163 |
| Wild boar | 22 | 0(0%) | 3 (13.64%) | 6 (27.27%) | 0(0%) | 0 (0%) | 11 (50%) | 0 (0%) | 2 (9.09%) | 0.973 | 0.68 |
| Vegetable | 24 | 1(4.17%) | 14 (58.33%) | 1 (4.17%) | 6 (25%) | 0 (0%) | 1 (4.17%) | 0 (0%) | 1 (4.17%) | 1.191 | 0.616 |
| Fishery | 24 | 10 (41.67%) | 4 (16.67%) | 2 (8.33%) | 5 (20.83%) | 0 (0%) | 3 (12.50%) | 0 (0%) | 0 (0%) | 1.457 | 0.764 |
| Companion animal | 12 | 0 (0%) | 6 (50%) | 2 (16.67%) | 2 (16.67%) | 0 (0%) | 2 (16.67%) | 0 (0%) | 0 (0%) | 1.089 | 0.644 |
| Swine | 25 | 8 (32%) | 8 (32%) | 0 (0%) | 8 (32%) | 0 (0%) | 1 (4%) | 0 (0%) | 0 (0%) | 1.223 | 0.720 |
| Poultry | 25 | 2 (8%) | 6 (24%) | 1 (4%) | 8 (32%) | 0 (0%) | 2 (8%) | 5 (20%) | 1 (4%) | 1.691 | 0.817 |
| Rabbit | 23 | 0 (0%) | 20 (86.96%) | 1 (4.35%) | 0 (0%) | 2 (8.70%) | 0 (0%) | 0 (0%) | 0 (0%) | 0.470 | 0.245 |
| Mollusc | 25 | 2 (8%) | 15 (60%) | 0 (0%) | 4 (16%) | 3 (12%) | 1 (4%) | 0 (0%) | 0 (0%) | 1.185 | 0.617 |
| Human | 25 | 6 (24%) | 2 (8%) | 2 (8%) | 3 (12%) | 2 (8%) | 3 (12%) | 7 (28%) | 0 (0%) | 1.814 | 0.850 |
| Wild animal | 25 | 1 (4%) | 18 (72%) | 3 (12%) | 2 (8%) | 0 (0%) | 0 (0%) | 1 (4%) | 0 (0%) | 0.951 | 0.477 |
| Total | 279 | 38 (13.62%) | 127 (45.52%) | 20 (7.17%) | 43 (15.41%) | 9 (3.23%) | 25 (8.96%) | 13 (4.66%) | 4 (1.43%) |
| Lineages | n | Functional Category Profile |
|---|---|---|
| Pandemic ExPEC | 13 | adhesin, iron acquisiton system (2) |
| adhesin, iron acquisiton system, protectin (8) | ||
| adhesin, toxin, iron acquisition system, protectin, invasin (3) | ||
| Emerging ExPEC | 37 | adhesin (2) |
| iron acquisition system (1) | ||
| adhesin, protectin (11) | ||
| adhesin, iron acquisition system, protectin (18) | ||
| adhesin, iron acquisition system, toxin, protectin (5) | ||
| Uncommon ExPEC | 229 | adhesin (68) |
| protectin (4) | ||
| adhesin, invasin (1) | ||
| adhesin, toxin (3) | ||
| adhesin, protectin (68) | ||
| adhesin, iron acquisition system (9) | ||
| protectin, iron acquisition system (1) | ||
| adhesin, toxin, protectin (2) | ||
| adhesin, iron acquisition system, protectin (50) | ||
| adhesin, invasin, iron acquisition system, toxin (1) | ||
| adhesin, invasin, iron acquisition system, protectin (2) | ||
| adhesin, toxin, iron acquisition, system, protectin (12) | ||
| adhesin, toxin, iron acquisition system, protectin, invasin (8) | ||
| Functional Category | Virulence Factor | Gene | Pathotype |
|---|---|---|---|
| Adhesin | M-agglutinin subunit | bmaE | UPEC |
| type 1 fimbrial adhesin | fimH | UPEC, NMEC, SEPEC, APEC | |
| iron-regulated-gene-homologue adhesin | iha | UPEC | |
| pilus associated with pyelonephritis | papC | UPEC, SEPEC, APEC | |
| pilus associated with pyelonephritis | papG_II; papG_III | UPEC, SEPEC, APEC | |
| S fimbrial adhesin | sfaS | UPEC, NMEC | |
| bifunctional enterobactin receptor/adhesin | iha | UPEC, NMEC | |
| temperature sensitive hemagglutinin | tsh | UPEC, NMEC, SEPEC, APEC | |
| Inasin | invasion of brain endothelium | ibeA | NMEC, SEPEC, APEC |
| genetic island associated with newborn meningitis | gimB | NMEC | |
| Iron acquisiton system | iron-responsive element | ireA | UPEC |
| catecholate siderophore receptor | iroN | UPEC, NMEC, SEPEC APEC | |
| ferric yersinia uptake | fyuA | UPEC, NMEC | |
| iron repressible protein | irp2 | NMEC | |
| l-lysine 6 monooxigenase | iucD | UPEC, APEC | |
| ferric aerobactin receptor precursor | iutA | UPEC, NMEC | |
| periplasmic iron binding protein | sitA | UPEC, APEC | |
| Toxin | cytolethal distending toxin B | cdtB | UPEC, NMEC, SEPEC APEC |
| cytotoxic necrotising factor | cnf1 | UPEC, MNEC, SEPEC | |
| haemolysin E | hlyE | UPEC | |
| serine protease autotransporters | pic | UPEC | |
| serine protease autotransporters | sat | UPEC | |
| uropathogenic specific protein | usp | UPEC | |
| vacuolating autotransporter toxin | vat | UPEC, APEC | |
| Protectin | increased serum survival | iss | NMEC, SEPEC, APEC |
| structural component of colicin V operon | cvaC | NMEC, SEPEC, APEC | |
| group II capsule antigens | kpsMT_II | NMEC, SEPEC | |
| outer membrane protein | ompT | UPEC, NMEC | |
| transfer protein | traT | NMEC, SEPEC | |
| K1 capsular polysaccharide | neuC | NMEC, UPEC | |
| Other | pathogenicity-associated island marker | malX | UPEC, NMEC, SEPEC, APEC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massella, E.; Giacometti, F.; Bonilauri, P.; Reid, C.J.; Djordjevic, S.P.; Merialdi, G.; Bacci, C.; Fiorentini, L.; Massi, P.; Bardasi, L.; et al. Antimicrobial Resistance Profile and ExPEC Virulence Potential in Commensal Escherichia coli of Multiple Sources. Antibiotics 2021, 10, 351. https://doi.org/10.3390/antibiotics10040351
Massella E, Giacometti F, Bonilauri P, Reid CJ, Djordjevic SP, Merialdi G, Bacci C, Fiorentini L, Massi P, Bardasi L, et al. Antimicrobial Resistance Profile and ExPEC Virulence Potential in Commensal Escherichia coli of Multiple Sources. Antibiotics. 2021; 10(4):351. https://doi.org/10.3390/antibiotics10040351
Chicago/Turabian StyleMassella, Elisa, Federica Giacometti, Paolo Bonilauri, Cameron J. Reid, Steven P. Djordjevic, Giuseppe Merialdi, Cristina Bacci, Laura Fiorentini, Paola Massi, Lia Bardasi, and et al. 2021. "Antimicrobial Resistance Profile and ExPEC Virulence Potential in Commensal Escherichia coli of Multiple Sources" Antibiotics 10, no. 4: 351. https://doi.org/10.3390/antibiotics10040351
APA StyleMassella, E., Giacometti, F., Bonilauri, P., Reid, C. J., Djordjevic, S. P., Merialdi, G., Bacci, C., Fiorentini, L., Massi, P., Bardasi, L., Rubini, S., Savini, F., Serraino, A., & Piva, S. (2021). Antimicrobial Resistance Profile and ExPEC Virulence Potential in Commensal Escherichia coli of Multiple Sources. Antibiotics, 10(4), 351. https://doi.org/10.3390/antibiotics10040351