
In Vitro Antibacterial, Anti-Adhesive and Anti-Biofilm Activities of Krameria lappacea (Dombey) Burdet & B.B. Simpson Root Extract against Methicillin-Resistant Staphylococcus aureus Strains

 ,
,  ,
,  , ,
, ,  , , ,
, , , 
Abstract
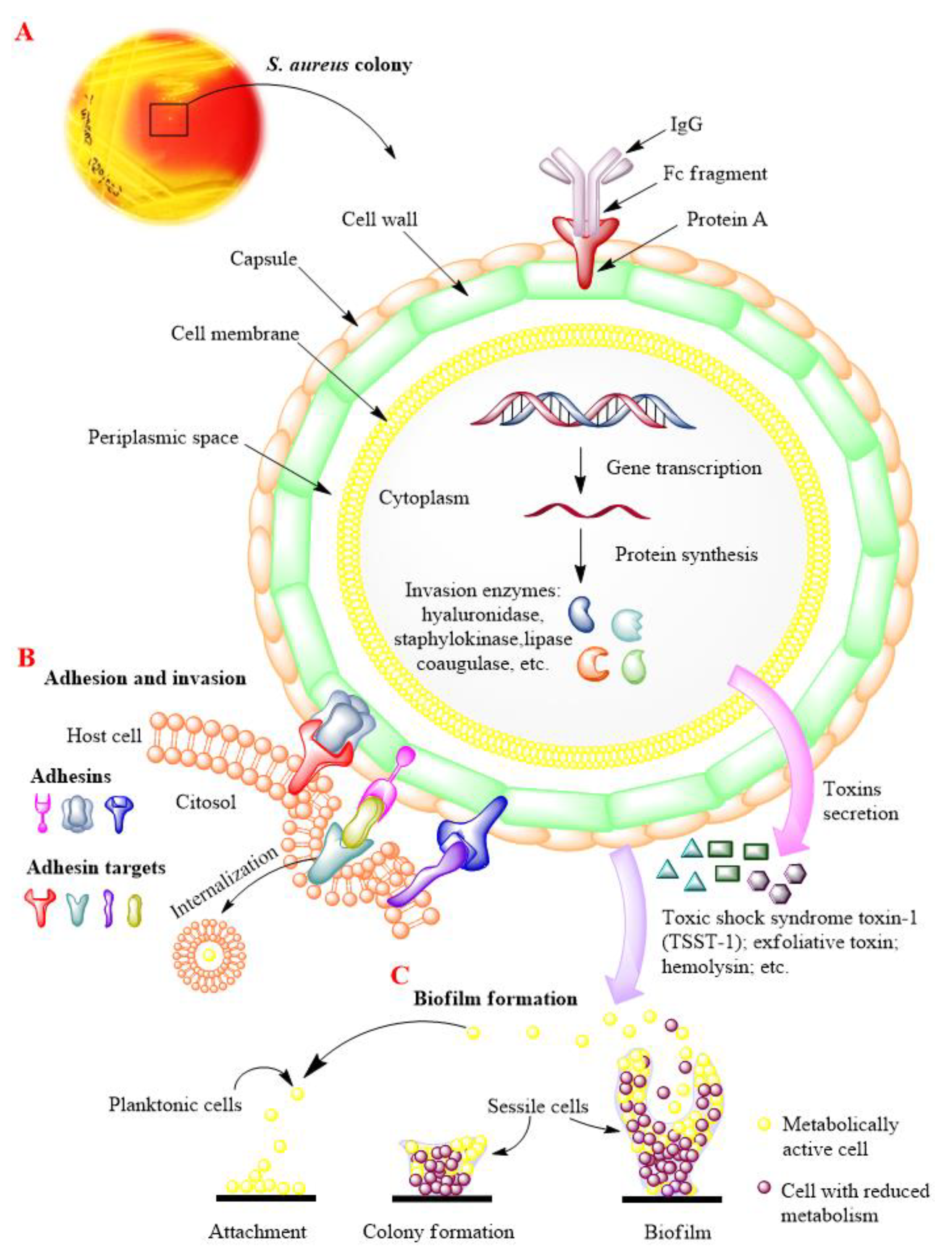
:1. Introduction
2. Results
2.1. Susceptibility of S. aureus Strains to Cefoxitin and Oxacillin
2.2. Detection of Macrolide-Lincosamide-Streptogramin B Phenotypes
2.3. Detection of Penicillin-Binding Protein 2a (PBP2a)
2.4. Antibacterial Activity
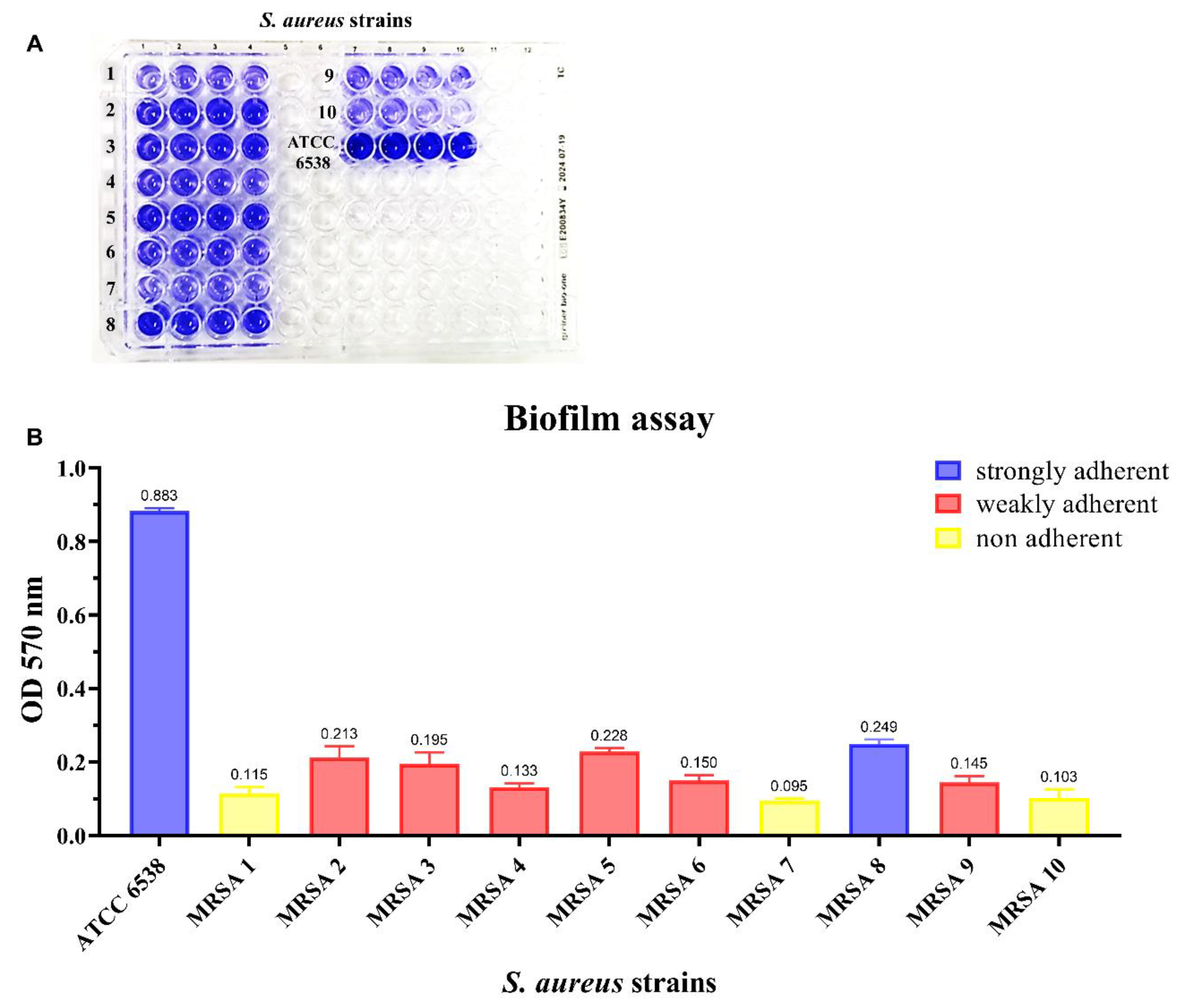
2.5. Biofilm Quantification Assay
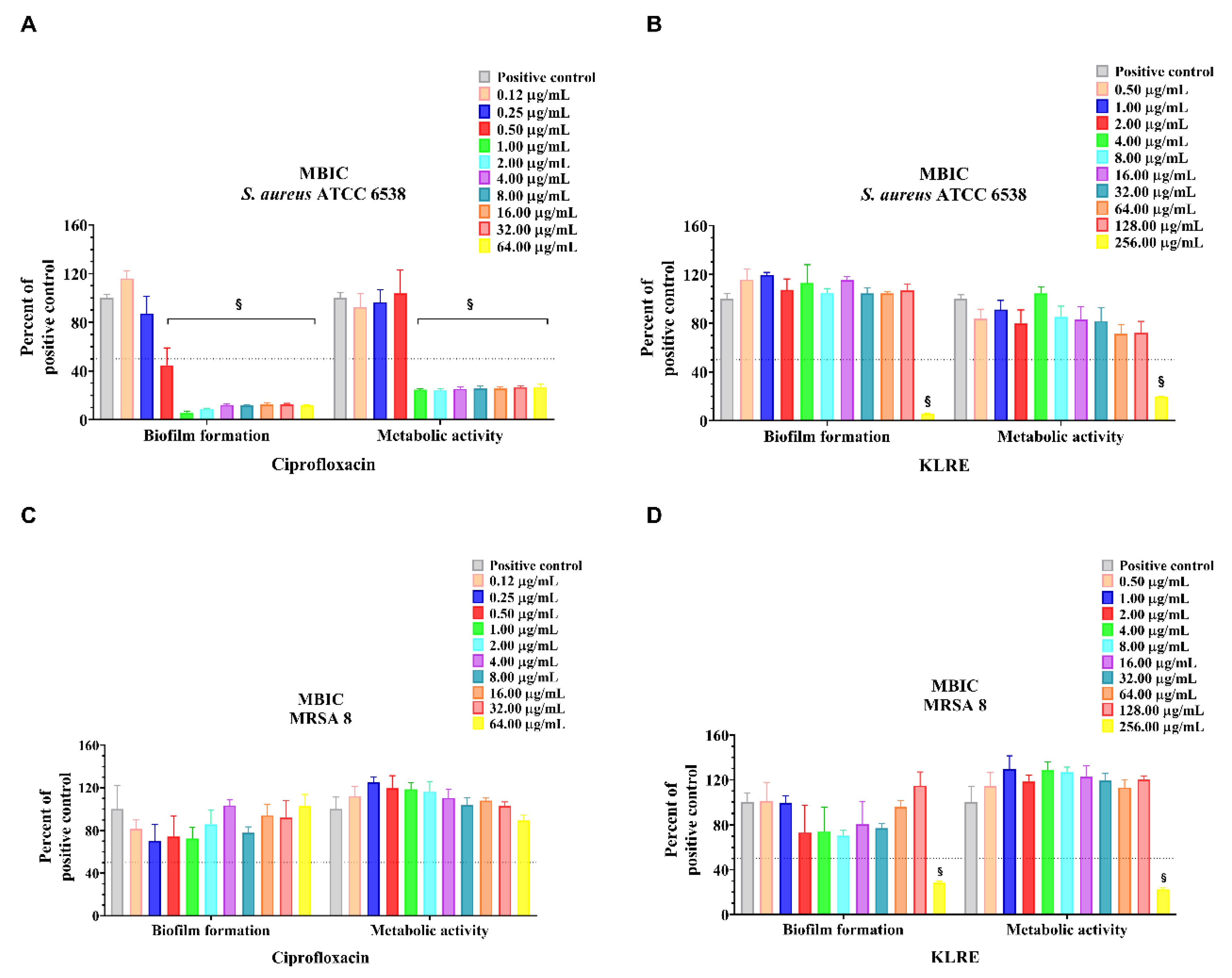
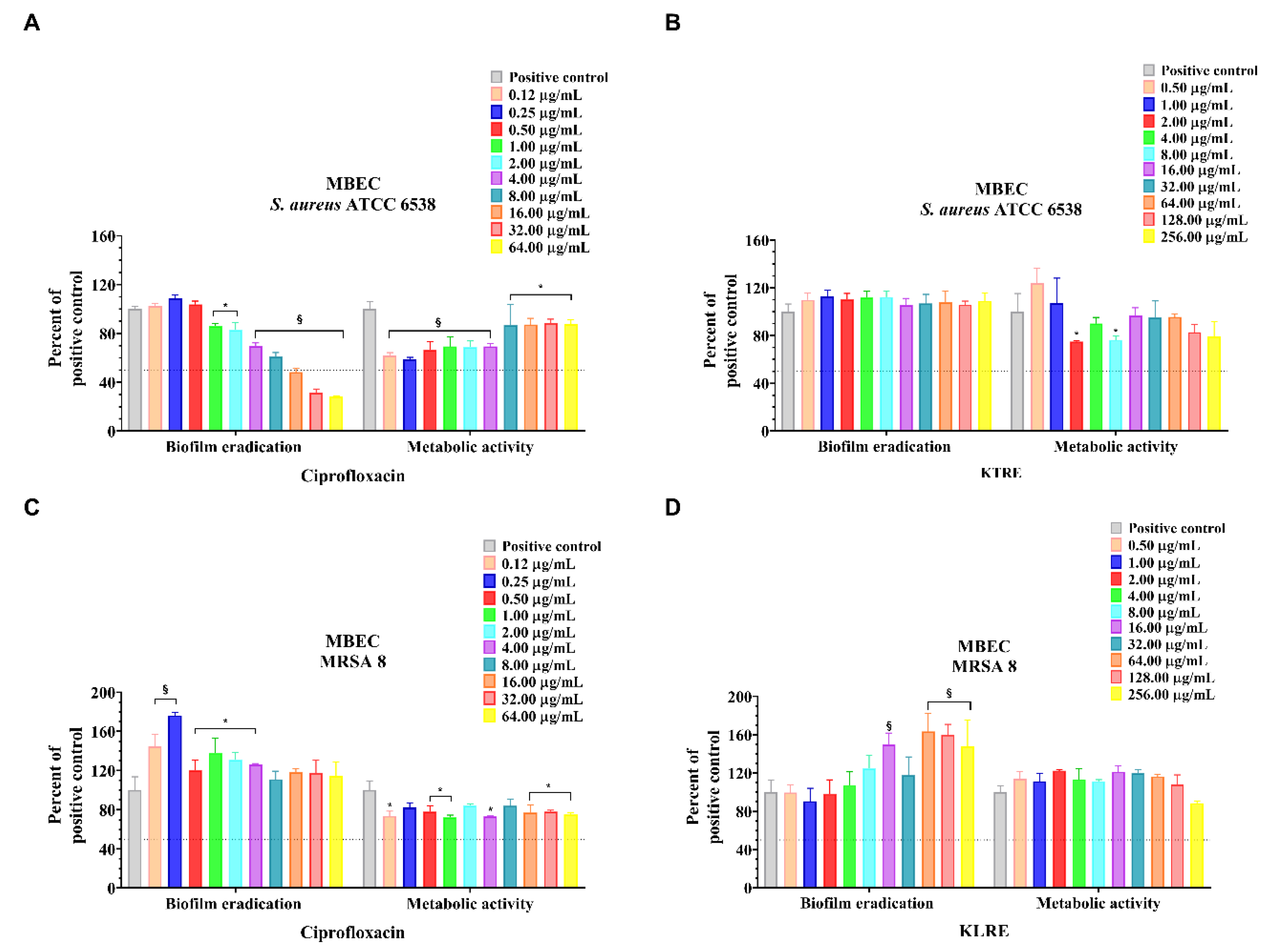
2.6. MBIC, MBEC and MTT Reduction Assay
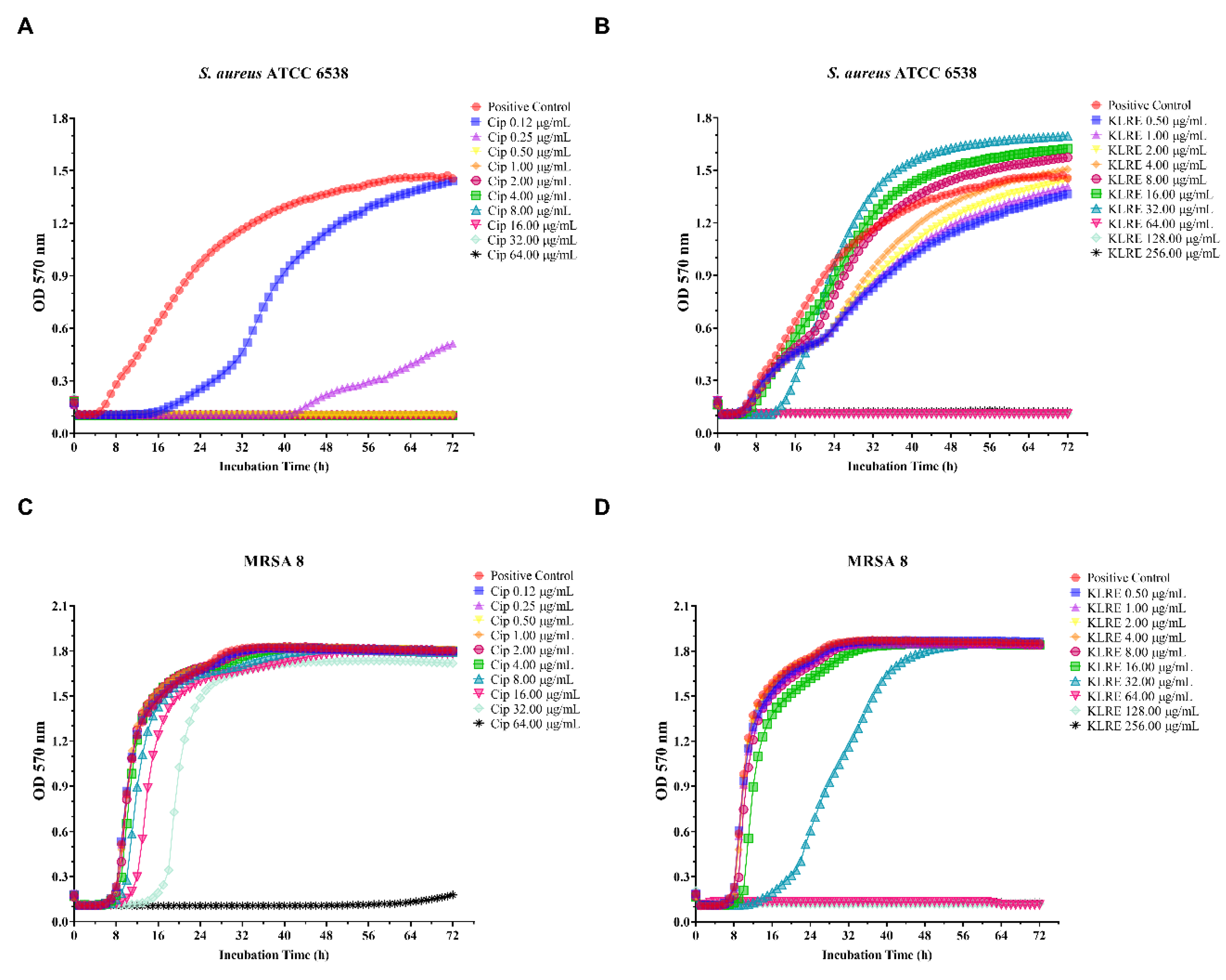
2.7. Effect of KLRE and Ciprofloxacin on Standard and Clinical S. aureus Strains: Growth Curves
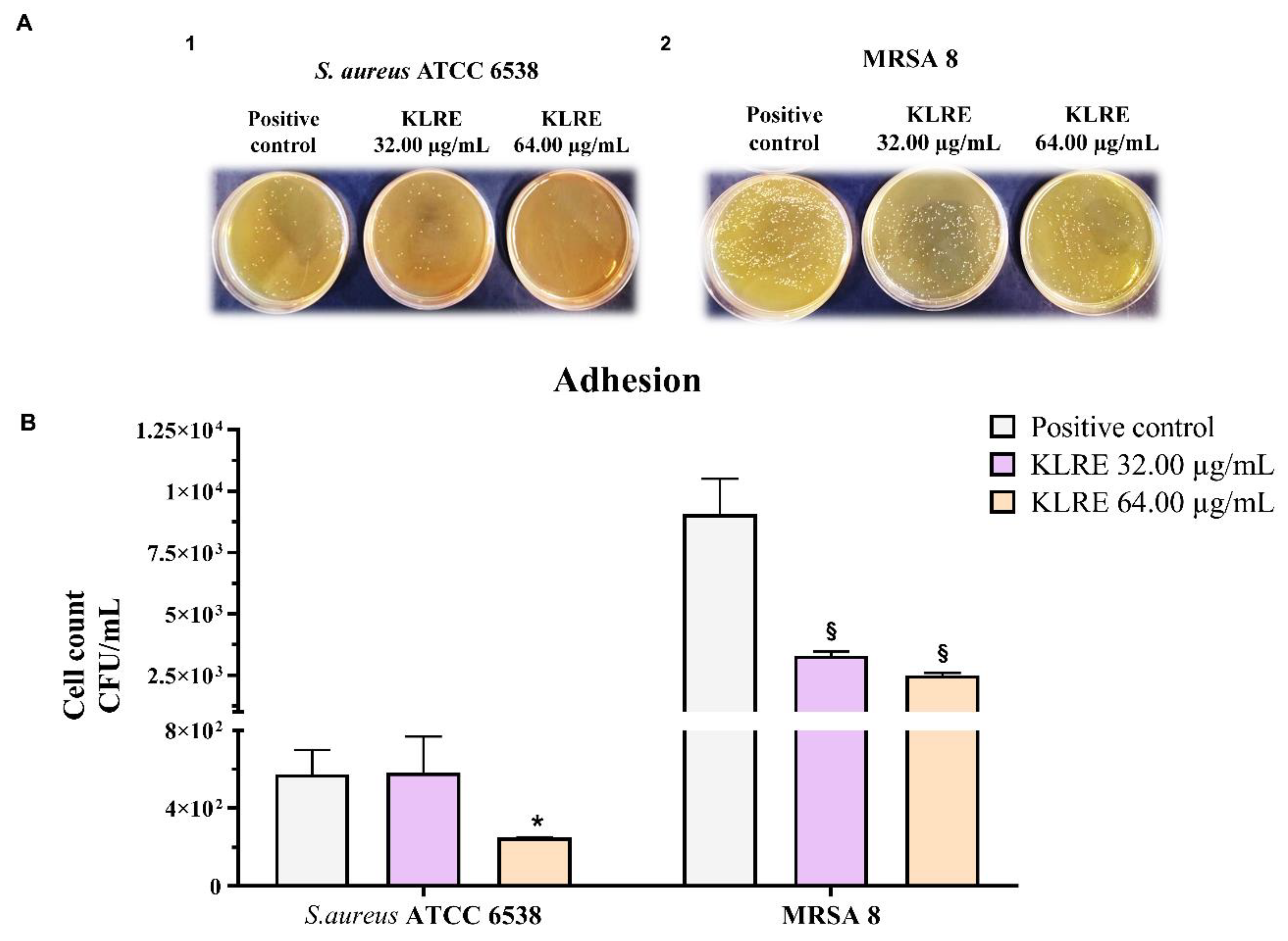
2.8. Effect of KLRE on the Adhesion of S. aureus to Human Lung A549 Cell Line
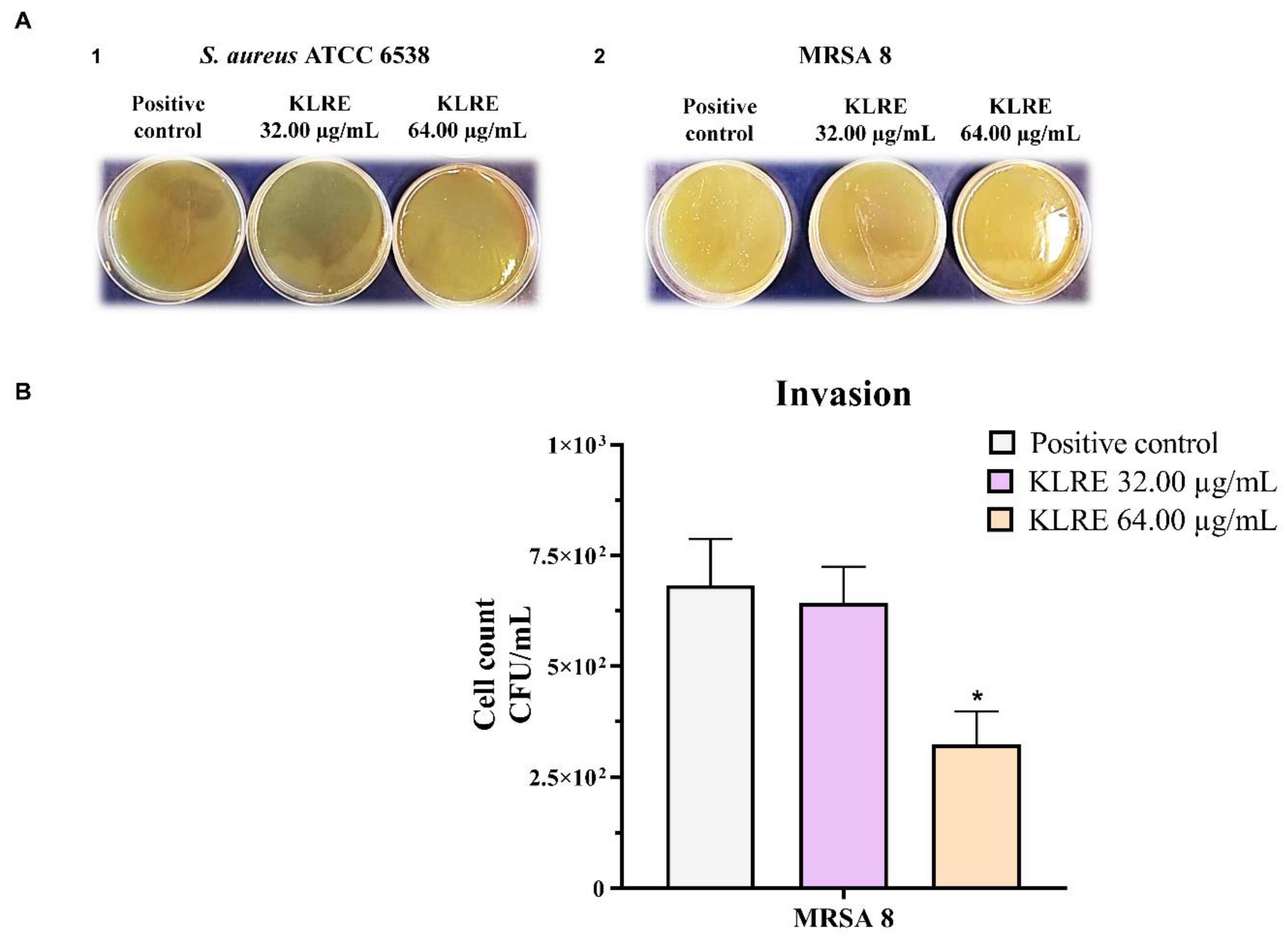
2.9. Effect of KLRE on the Invasion of S. aureus to Human Lung A549 Cell Line
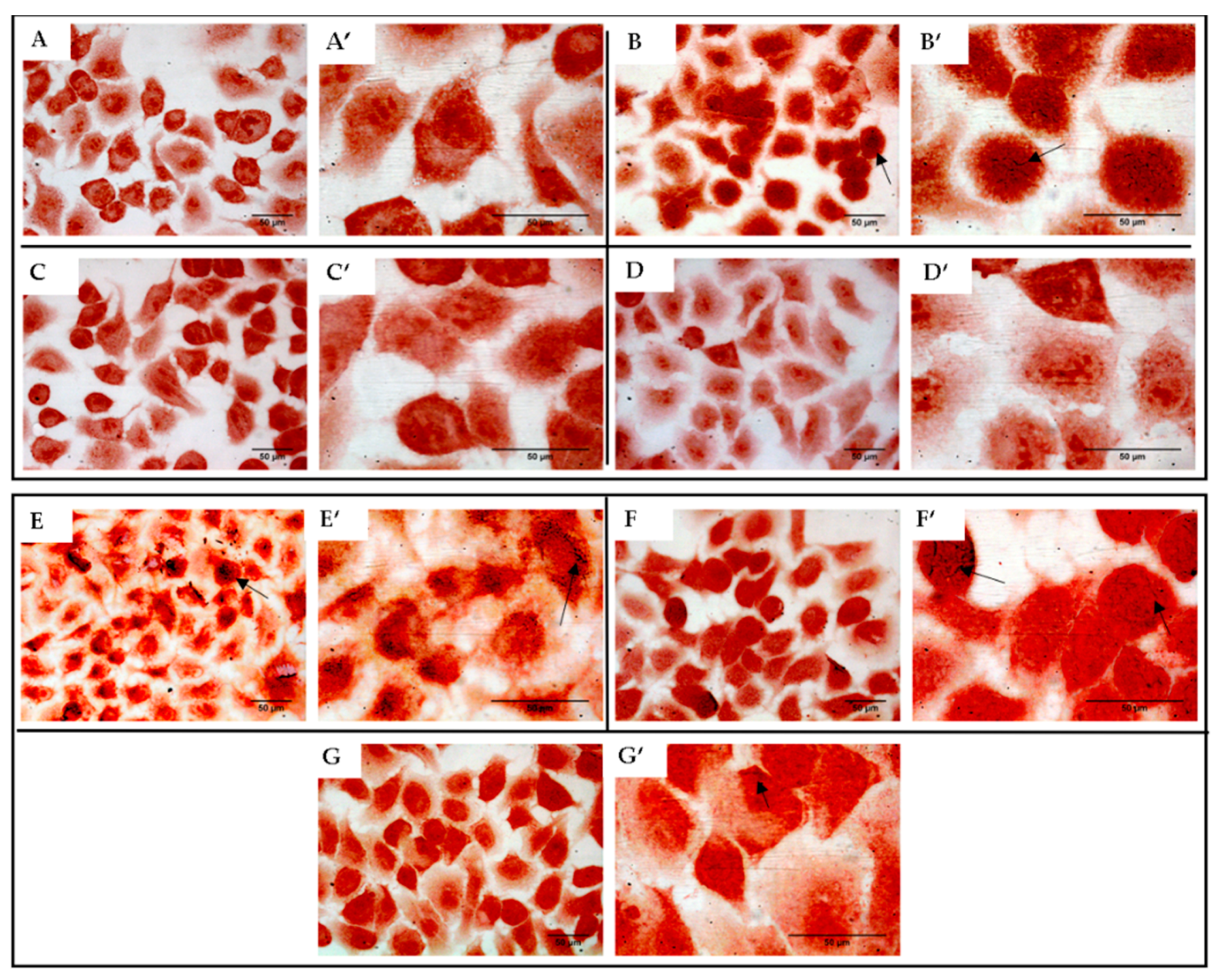
2.10. Effect of KLRE on the Adhesion of S. aureus to Human Lung A549 Cell Line: Gram Staining
2.11. Detection of Flavonoids and Proanthocyanidins by UHPLC-HRMS
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material and Preparation of the Extract
4.3. Bacterial Strains and Human Cells
4.4. Determination of S. aureus Strains Susceptibility to Cefoxitin and Oxacillin by Disc-Diffusion Test
4.5. Detection of Macrolide-Lincosamide-Streptogramin B Phenotypes
4.6. Detection of Penicillin-Binding Protein 2a (PBP2a)
4.7. Antibacterial Activity of Krameria Lappacea Root Extract (KLRE)
4.8. Biofilm Quantification Assay
4.9. Minimum Biofilm Inhibitory Concentration Assay
4.10. Minimum Biofilm Eradication Concentration Assay
4.11. MTT Reduction Assay
4.12. Growth Curve
4.13. Cell Culture
4.14. Bacterial Invasion and Adhesion Assay
4.15. Gram Staining
4.16. Flavonoid Extraction and UHPLC-HRMS Analysis
4.17. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gould, D.; Chamberlaine, A. Staphylococcus aureus: A review of the literature. J. Clin. Nurs. 1995, 4, 5–12. [Google Scholar] [CrossRef]
- Sivaraman, K.; Venkataraman, N.; Cole, A.M. Staphylococcus aureus nasal carriage and its contributing factors. Future Microbiol. 2009, 4, 999–1008. [Google Scholar] [CrossRef]
- Yang, E.S.; Tan, J.; Eells, S.; Rieg, G.; Tagudar, G.; Miller, L.G. Body site colonization in patients with community-associated methicillin-resistant Staphylococcus aureus and other types of S. aureus skin infections. Clin. Microbiol. Infect. 2010, 16, 425–431. [Google Scholar] [CrossRef]
- Foster, T.J. Surface Proteins of Staphylococcus aureus. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Hook, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef]
- Leonard, A.C.; Petrie, L.E.; Cox, G. Bacterial Anti-adhesives: Inhibition of staphylococcus aureus nasal colonization. ACS Infect. Dis. 2019, 5, 1668–1681. [Google Scholar] [CrossRef]
- Visansirikul, S.; Kolodziej, S.A.; Demchenko, A.V. Staphylococcus aureus capsular polysaccharides: A structural and synthetic perspective. Org. Biomol. Chem. 2020, 18, 783–798. [Google Scholar] [CrossRef]
- O’Riordan, K.; Lee, J.C. Staphylococcus aureus capsular polysaccharides. Clin. Microbiol. Rev. 2004, 17, 218–234. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Liu, W.; Arora, S.; Ganesh, V.; Ko, Y.P.; Hook, M. The complex fibrinogen interactions of the staphylococcus aureus coagulases. Front. Cell Infect. Microbiol. 2019, 9, 106. [Google Scholar] [CrossRef]
- Hu, C.; Xiong, N.; Zhang, Y.; Rayner, S.; Chen, S. Functional characterization of lipase in the pathogenesis of Staphylococcus aureus. Biochem. Biophys. Res. Commun. 2012, 419, 617–620. [Google Scholar] [CrossRef]
- Ibberson, C.B.; Jones, C.L.; Singh, S.; Wise, M.C.; Hart, M.E.; Zurawski, D.V.; Horswill, A.R. Staphylococcus aureus hyaluronidase is a CodY-regulated virulence factor. Infect. Immun. 2014, 82, 4253–4264. [Google Scholar] [CrossRef]
- Bokarewa, M.I.; Jin, T.; Tarkowski, A. Staphylococcus aureus: Staphylokinase. Int. J. Biochem. Cell Biol. 2006, 38, 504–509. [Google Scholar] [CrossRef]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus biofilms: Recent developments in biofilm dispersal. Front. Cell Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef]
- Liu, J.; Chen, D.; Peters, B.M.; Li, L.; Li, B.; Xu, Z.; Shirliff, M.E. Staphylococcal chromosomal cassettes mec (SCCmec): A mobile genetic element in methicillin-resistant Staphylococcus aureus. Microb. Pathog. 2016, 101, 56–67. [Google Scholar] [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-resistant staphylococcus aureus: Molecular characterization, evolution, and epidemiology. Clin. Microbiol. Rev. 2018, 31. [Google Scholar] [CrossRef]
- ElFeky, D.S.; Awad, A.R.; Elshobaky, M.A.; Elawady, B.A. Effect of Ceftaroline, vancomycin, gentamicin, macrolides, and ciprofloxacin against methicillin-resistant staphylococcus aureus isolates: An in vitro study. Surg. Infect. (Larchmt) 2020, 21, 150–157. [Google Scholar] [CrossRef]
- Jernigan, J.A.; Hatfield, K.M.; Wolford, H.; Nelson, R.E.; Olubajo, B.; Reddy, S.C.; McCarthy, N.; Paul, P.; McDonald, L.C.; Kallen, A.; et al. Multidrug-resistant bacterial infections in U.S. hospitalized patients, 2012–2017. N. Engl. J. Med. 2020, 382, 1309–1319. [Google Scholar] [CrossRef]
- De la Calle, C.; Morata, L.; Cobos-Trigueros, N.; Martinez, J.A.; Cardozo, C.; Mensa, J.; Soriano, A. Staphylococcus aureus bacteremic pneumonia. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 497–502. [Google Scholar] [CrossRef]
- Defres, S.; Marwick, C.; Nathwani, D. MRSA as a cause of lung infection including airway infection, community-acquired pneumonia and hospital-acquired pneumonia. Eur. Respir. J. 2009, 34, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Genovese, C.; Acquaviva, R.; Ronsisvalle, S.; Tempera, G.; Antonio Malfa, G.; D’Angeli, F.; Ragusa, S.; Nicolosi, D. In vitro evaluation of biological activities of orobanche crenata forssk. leaves extract. Nat. Prod. Res. 2020, 34, 3234–3238. [Google Scholar] [CrossRef]
- Acquaviva, R.; D’Angeli, F.; Malfa, G.A.; Ronsisvalle, S.; Garozzo, A.; Stivala, A.; Ragusa, S.; Nicolosi, D.; Salmeri, M.; Genovese, C. Antibacterial and anti-biofilm activities of walnut pellicle extract (Juglans regia L.) against coagulase-negative staphylococci. Nat. Prod. Res. 2019, 1–6. [Google Scholar] [CrossRef]
- Simpson, B.B. Krameriaceae (Flora Neotropica); New York Botanical Garden Press: Bronx, NY, USA, 1989; Volume 49, pp. 1–108. [Google Scholar]
- Simpson, B.B. The past and present uses of rhatany (Krameria, Krameriaceae). Econ. Bot. 1991, 45, 397–409. [Google Scholar] [CrossRef]
- Carini, M.; Aldini, G.; Orioli, M.; Facino, R.M. Antioxidant and photoprotective activity of a lipophilic extract containing neolignans from Krameria triandra roots. Planta Med. 2002, 68, 193–197. [Google Scholar] [CrossRef]
- Baumgartner, L.; Sosa, S.; Atanasov, A.G.; Bodensieck, A.; Fakhrudin, N.; Bauer, J.; Favero, G.D.; Ponti, C.; Heiss, E.H.; Schwaiger, S.; et al. Lignan derivatives from Krameria lappacea roots inhibit acute inflammation in vivo and pro-inflammatory mediators in vitro. J. Nat. Prod. 2011, 74, 1779–1786. [Google Scholar] [CrossRef]
- Facino, R.M.; Carini, M.; Aldini, G.; DeAngelis, L. A rapid screening by liquid chromatography mass spectrometry and fast-atom bombardment tandem mass spectrometry of phenolic constituents with radical scavenging activity, from Krameria triandra roots. Rapid Commun. Mass Spectrom. 1997, 11, 1303–1308. [Google Scholar] [CrossRef]
- Scholz, E.; Rimpler, H. Proanthocyanidins from Krameria triandra root. Planta Med. 1989, 55, 379–384. [Google Scholar] [CrossRef]
- Ortiz, S.; Lecso-Bornet, M.; Bonnal, C.; Houze, S.; Michel, S.; Grougnet, R.; Boutefnouchet, S. Bioguided identification of triterpenoids and neolignans as bioactive compounds from anti-infectious medicinal plants of the Taira Atacama’s updates community (Calama, Chile). J. Ethnopharmacol. 2019, 231, 217–229. [Google Scholar] [CrossRef]
- CLSI 2020. Performance Standards for Antimicrobial Susceptibility Testing; 30th Informational Supplement, CLSI Document M100-S30; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Manandhar, S.; Singh, A.; Varma, A.; Pandey, S.; Shrivastava, N. Biofilm producing clinical staphylococcus aureus isolates augmented prevalence of antibiotic resistant cases in tertiary care hospitals of nepal. Front. Microbiol. 2018, 9, 2749. [Google Scholar] [CrossRef]
- Abbas, H.A.; Elsherbini, A.M.; Shaldam, M.A. Glyceryl trinitrate blocks staphyloxanthin and biofilm formation in Staphylococcus aureus. Afr. Health Sci. 2019, 19, 1376–1384. [Google Scholar] [CrossRef]
- Lanks, C.W.; Musani, A.I.; Hsia, D.W. Community-acquired Pneumonia and Hospital-acquired Pneumonia. Med. Clin. N. Am. 2019, 103, 487–501. [Google Scholar] [CrossRef]
- Webb, B.J.; Dascomb, K.; Stenehjem, E.; Dean, N. Predicting risk of drug-resistant organisms in pneumonia: Moving beyond the HCAP model. Respir. Med. 2015, 109, 1–10. [Google Scholar] [CrossRef]
- Kollef, M.H.; Shorr, A.; Tabak, Y.P.; Gupta, V.; Liu, L.Z.; Johannes, R.S. Epidemiology and outcomes of health-care-associated pneumonia-Results from a large US database of culture-positive pneumonia. Chest 2005, 128, 3854–3862. [Google Scholar] [CrossRef]
- Barbier, F.; Andremont, A.; Wolff, M.; Bouadma, L. Hospital-acquired pneumonia and ventilator-associated pneumonia: Recent advances in epidemiology and management. Curr. Opin. Pulm. Med. 2013, 19, 216–228. [Google Scholar] [CrossRef]
- Karakuzu, Z.; Iscimen, R.; Akalin, H.; Girgin, N.K.; Kahveci, F.; Sinirtas, M. Prognostic risk factors in ventilator-associated pneumonia. Med. Sci. Monitor. 2018, 24, 1321–1328. [Google Scholar] [CrossRef]
- Nguile-Makao, M.; Zahar, J.R.; Francais, A.; Tabah, A.; Garrouste-Orgeas, M.; Allaouchiche, B.; Goldgran-Toledano, D.; Azoulay, E.; Adrie, C.; Jamali, S.; et al. Attributable mortality of ventilator-associated pneumonia: Respective impact of main characteristics at ICU admission and VAP onset using conditional logistic regression and multi-state models. Intensive Care Med. 2010, 36, 781–789. [Google Scholar] [CrossRef]
- Somers, E.C.; Eschenauer, G.A.; Troost, J.P.; Golob, J.L.; Gandhi, T.N.; Wang, L.; Zhou, N.; Petty, L.A.; Baang, J.H.; Dillman, N.O.; et al. Tocilizumab for treatment of mechanically ventilated patients with COVID-19. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Sharifipour, E.; Shams, S.; Esmkhani, M.; Khodadadi, J.; Fotouhi-Ardakani, R.; Koohpaei, A.; Doosti, Z.; Ej Golzari, S. Evaluation of bacterial co-infections of the respiratory tract in COVID-19 patients admitted to ICU. BMC Infect. Dis. 2020, 20, 646. [Google Scholar] [CrossRef]
- Saraiva, A.M.; Castro, R.H.A.; Cordeiro, R.P.; Sobrinho, T.J.S.P.; Castro, V.T.N.A.; Amorim, E.L.C.; Xavier, H.S.; Pisciottano, M.N.C. In vitro evaluation of antioxidant, antimicrobial and toxicity properties of extracts of Schinopsis brasiliensis Engl. (Anacardiaceae). Afr. J. Pharm. Pharmacol. 2011, 5, 1724–1731. [Google Scholar] [CrossRef]
- Wu, H.; Moser, C.; Wang, H.Z.; Hoiby, N.; Song, Z.J. Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 2015, 7, 1–7. [Google Scholar] [CrossRef]
- Nguyen, T.K.; Argudin, M.A.; Deplano, A.; Nhung, P.H.; Nguyen, H.A.; Tulkens, P.M.; Dodemont, M.; Van Bambeke, F. Antibiotic resistance, biofilm formation, and intracellular survival as possible determinants of persistent or recurrent infections bystaphylococcus aureusin a vietnamese tertiary hospital: Focus on bacterial response to moxifloxacin. Microb. Drug Resist. 2020, 26, 537–544. [Google Scholar] [CrossRef]
- Lu, C.; Liu, H.; Shangguan, W.; Chen, S.; Zhong, Q. Antibiofilm activities of the cinnamon extract against Vibrio parahaemolyticus and Escherichia coli. Arch. Microbiol. 2021, 203, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Miari, M.; Rasheed, S.S.; Ahmad, N.H.; Itani, D.; Abou Fayad, A.; Matar, G.M. Natural products and polysorbates: Potential Inhibitors of biofilm formation in Pseudomonas aeruginosa. J. Infect. Dev. Countr. 2020, 14, 580. [Google Scholar] [CrossRef]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef]
- Wolfmeier, H.; Pletzer, D.; Mansour, S.C.; Hancock, R.E.W. New perspectives in biofilm eradication. ACS Infect. Dis. 2018, 4, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Sinha, B.; Francois, P.P.; Nusse, O.; Foti, M.; Hartford, O.M.; Vaudaux, P.; Foster, T.J.; Lew, D.P.; Herrmann, M.; Krause, K.H. Fibronectin-binding protein acts as Staphylococcus aureus invasin via fibronectin bridging to integrin alpha(5)beta(1). Cell Microbiol. 1999, 1, 101–117. [Google Scholar] [CrossRef]
- de Moura, P.H.B.; de Sousa, A.A.; Porzel, A.; Wessjohann, L.A.; Leal, I.C.R.; Martins, R.C.C. Characterization of antibacterial proanthocyanidins of dalbergia monetaria, an amazonian medicinal plant, by UHPLC-HRMS/MS. Planta Med. 2020, 86, 858–866. [Google Scholar] [CrossRef]
- Craft, K.M.; Nguyen, J.M.; Berg, L.J.; Townsend, S.D. Methicillin-resistant Staphylococcus aureus (MRSA): Antibiotic-resistance and the biofilm phenotype. Medchemcomm 2019, 10, 1231–1241. [Google Scholar] [CrossRef]
- Nicolosi, D.; Tempera, G.; Genovese, C.; Furneri, P.M. Anti-adhesion activity of A2-type proanthocyanidins (a cranberry major component) on uropathogenic E. coli and P. mirabilis Strains. Antibiotics (Basel) 2014, 3, 143–154. [Google Scholar] [CrossRef]
- Ahmed, G.F.; Elkhatib, W.F.; Noreddin, A.M. Inhibition of Pseudomonas aeruginosa PAO1 adhesion to and invasion of A549 lung epithelial cells by natural extracts. J. Infect. Public Health 2014, 7, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Hui, J.; Choy, J.; Suwandaratne, S.P.; Shervill, J.; Gan, B.S.; Howard, J.C.; Reid, G. Antimicrobial activity of Vaccinium macrocarpon (cranberry) produced proanthocya-nidin (PAC) on the growth and adhesion properties of Staphylococcus aureus. J. Food Sci. Nutr. New Ser. 2004, 9, 29–33. [Google Scholar]
- Howell, A.B.; Reed, J.D.; Krueger, C.G.; Winterbottom, R.; Cunningham, D.G.; Leahy, M. A-type cranberry proanthocyanidins and uropathogenic bacterial anti-adhesion activity. Phytochemistry 2005, 66, 2281–2291. [Google Scholar] [CrossRef]
- Genovese, C.; D’Angeli, F.; Di Salvatore, V.; Tempera, G.; Nicolosi, D. Streptococcus agalactiae in pregnant women: Serotype and antimicrobial susceptibility patterns over five years in Eastern Sicily (Italy). Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 2387–2396. [Google Scholar] [CrossRef]
- Mogana, R.; Adhikari, A.; Tzar, M.N.; Ramliza, R.; Wiart, C. Antibacterial activities of the extracts, fractions and isolated compounds from Canarium patentinervium Miq. against bacterial clinical isolates. BMC Complement. Med. Ther. 2020, 20, 55. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Stiefel, P.; Rosenberg, U.; Schneider, J.; Mauerhofer, S.; Maniura-Weber, K.; Ren, Q. Is biofilm removal properly assessed? Comparison of different quantification methods in a 96-well plate system. Appl. Microbiol. Biotechnol. 2016, 100, 4135–4145. [Google Scholar] [CrossRef]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: A quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef]
- Heydari, S.; Eftekhar, F. Biofilm Formation and beta-Lactamase Production in Burn Isolates of Pseudomonas aeruginosa. Jundishapur. J. Microbiol. 2015, 8, e15514. [Google Scholar] [CrossRef]
- Alvarez-Escobar, M.; Freitas, S.C.; Hansford, D.; Monteiro, F.J.; Pelaez-Vargas, A. Soft lithography and minimally human invasive technique for rapid screening of oral biofilm formation on new microfabricated dental material surfaces. Int. J. Dent. 2018, 2018, 4219625. [Google Scholar] [CrossRef]
- Cue, D.; Dombek, P.E.; Lam, H.; Cleary, P.P. Streptococcus pyogenes serotype M1 encodes multiple pathways for entry into human epithelial cells. Infect. Immun. 1998, 66, 4593–4601. [Google Scholar] [CrossRef]
- Gephart, P.; Murray, R.G.E.; Costilow, R.N.; Nester, E.W.; Wood, W.A.; Krieg, N.R.; Phillips, G.B. Manual of Methods for General Bacteriology; Gram., C., Ed.; ASM Press: Washington, DC, USA, 1884. [Google Scholar]
- Gram, C. Ueber die isolirte Färbung der Schizomyceten in Schnitt-und Trockenpräparaten. Fortschr. Med. 1981, 2, 185–189. [Google Scholar]
- Falcone, E.; Ahmed, I.M.M.; Oliveri, V.; Bellia, F.; Vileno, B.; El Khoury, Y.; Hellwig, P.; Faller, P.; Vecchio, G. Acrolein and Copper as Competitive Effectors of alpha-Synuclein. Chem. Eur. J. 2020, 26, 1871–1879. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
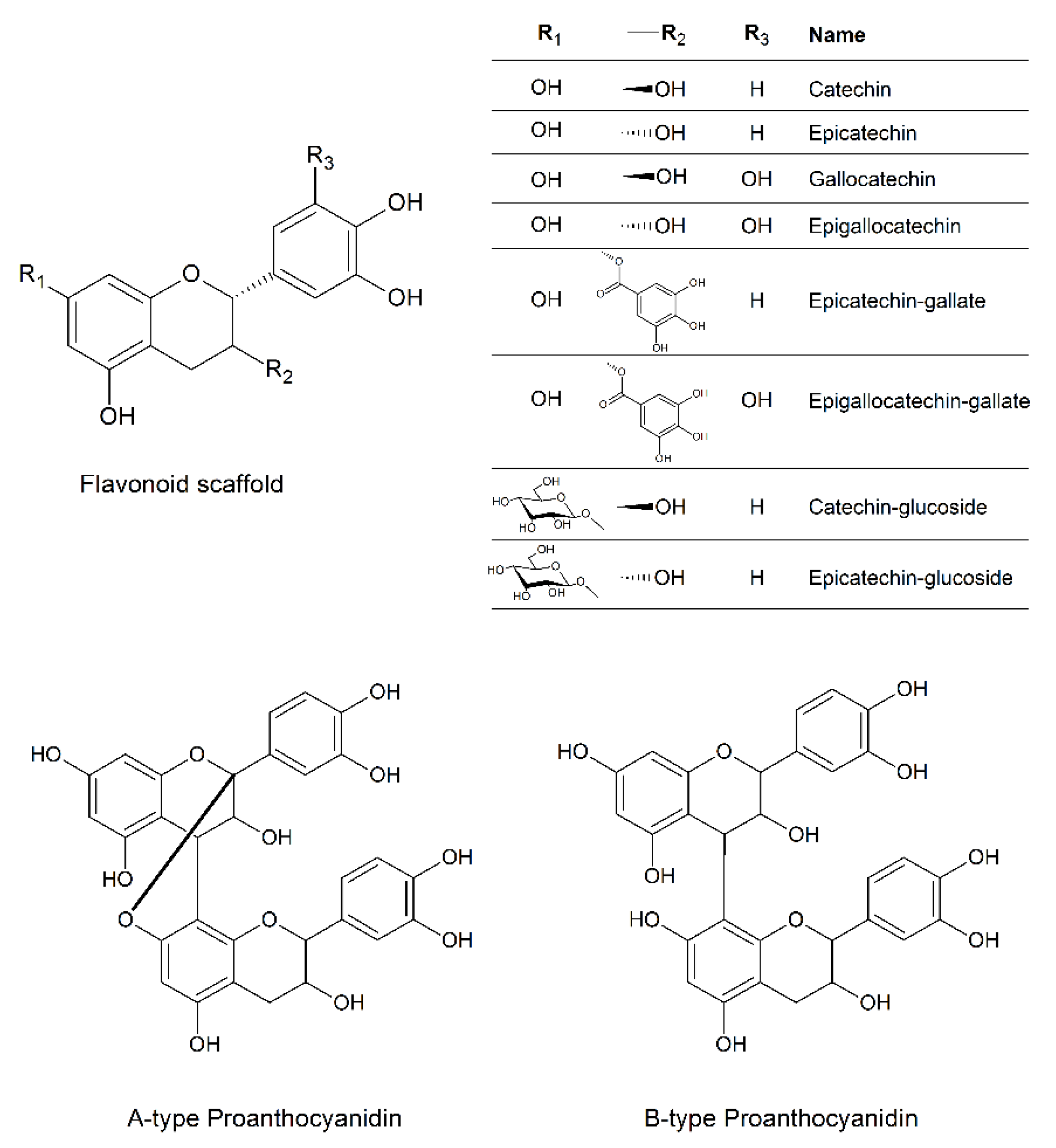{kind=link}
{kind=link}
| KLRE Range [256.00-0.50 µg/mL] | Ciprofloxacin Range [64.00-0.12 µg/mL] | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Bacterial strains a | MIC b | MBC c | MBC/MIC d | EFF. e | MIC | MBC | MBC/MIC | EFF. | I.C. f |
| S. aureus ATCC 6538 | 64.00 | 128.00 | 2 | BC | 0.25 | 0.25 | 1 | BC | S |
| MRSA1 | 64.00 | 128.00 | 2 | BC | 32.00 | 32.00 | 1 | BC | R |
| MRSA 2 | 64.00 | 256.00 | 4 | BC | 64.00 | 64.00 | 1 | BC | R |
| MRSA 3 | 64.00 | 256.00 | 4 | BC | 64.00 | 64.00 | 1 | BC | R |
| MRSA 4 | 64.00 | 128.00 | 2 | BC | 32.00 | 32.00 | 1 | BC | R |
| MRSA 5 | 64.00 | 128.00 | 2 | BC | 32.00 | 32.00 | 1 | BC | R |
| MRSA 6 | 64.00 | 256.00 | 4 | BC | 32.00 | 32.00 | 1 | BC | R |
| MRSA 7 | 64.00 | 128.00 | 2 | BC | 64.00 | 64.00 | 1 | BC | R |
| MRSA 8 | 64.00 | 128.00 | 2 | BC | 64.00 | 64.00 | 1 | BC | R |
| MRSA 9 | 64.00 | 256.00 | 4 | BC | 32.00 | 32.00 | 1 | BC | R |
| MRSA 10 | 64.00 | 128.00 | 2 | BC | 16.00 | 16.00 | 1 | BC | R |
| KLRE Range [256.00–0.50 µg/mL] | Ciprofloxacin Range [64.00–0.12 µg/mL] | |||||||
|---|---|---|---|---|---|---|---|---|
| Bacterial Strains | MBIC50 a | ID50 MBIC b | MBEC50 c | ID50 MBEC d | MBIC50 | ID50 MBIC | MBEC50 | ID50 MBEC |
| S. aureus ATCC 6538 | 256.00 | 256.00 | >256.00 | >256.00 | 1.00 | 1.00 | 16.00 | >64.00 |
| MRSA 8 e | 256.00 | 256.00 | >256.00 | >256.00 | >64.00 | >64.00 | >64.00 | >64.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genovese, C.; D’Angeli, F.; Bellia, F.; Distefano, A.; Spampinato, M.; Attanasio, F.; Nicolosi, D.; Di Salvatore, V.; Tempera, G.; Lo Furno, D.; et al. In Vitro Antibacterial, Anti-Adhesive and Anti-Biofilm Activities of Krameria lappacea (Dombey) Burdet & B.B. Simpson Root Extract against Methicillin-Resistant Staphylococcus aureus Strains. Antibiotics 2021, 10, 428. https://doi.org/10.3390/antibiotics10040428
Genovese C, D’Angeli F, Bellia F, Distefano A, Spampinato M, Attanasio F, Nicolosi D, Di Salvatore V, Tempera G, Lo Furno D, et al. In Vitro Antibacterial, Anti-Adhesive and Anti-Biofilm Activities of Krameria lappacea (Dombey) Burdet & B.B. Simpson Root Extract against Methicillin-Resistant Staphylococcus aureus Strains. Antibiotics. 2021; 10(4):428. https://doi.org/10.3390/antibiotics10040428
Chicago/Turabian StyleGenovese, Carlo, Floriana D’Angeli, Francesco Bellia, Alfio Distefano, Mariarita Spampinato, Francesco Attanasio, Daria Nicolosi, Valentina Di Salvatore, Gianna Tempera, Debora Lo Furno, and et al. 2021. "In Vitro Antibacterial, Anti-Adhesive and Anti-Biofilm Activities of Krameria lappacea (Dombey) Burdet & B.B. Simpson Root Extract against Methicillin-Resistant Staphylococcus aureus Strains" Antibiotics 10, no. 4: 428. https://doi.org/10.3390/antibiotics10040428
APA StyleGenovese, C., D’Angeli, F., Bellia, F., Distefano, A., Spampinato, M., Attanasio, F., Nicolosi, D., Di Salvatore, V., Tempera, G., Lo Furno, D., Mannino, G., Milardo, F., & Li Volti, G. (2021). In Vitro Antibacterial, Anti-Adhesive and Anti-Biofilm Activities of Krameria lappacea (Dombey) Burdet & B.B. Simpson Root Extract against Methicillin-Resistant Staphylococcus aureus Strains. Antibiotics, 10(4), 428. https://doi.org/10.3390/antibiotics10040428