
Adsorption of Phenazines Produced by Pseudomonas aeruginosa Using AST-120 Decreases Pyocyanin-Associated Cytotoxicity


Abstract
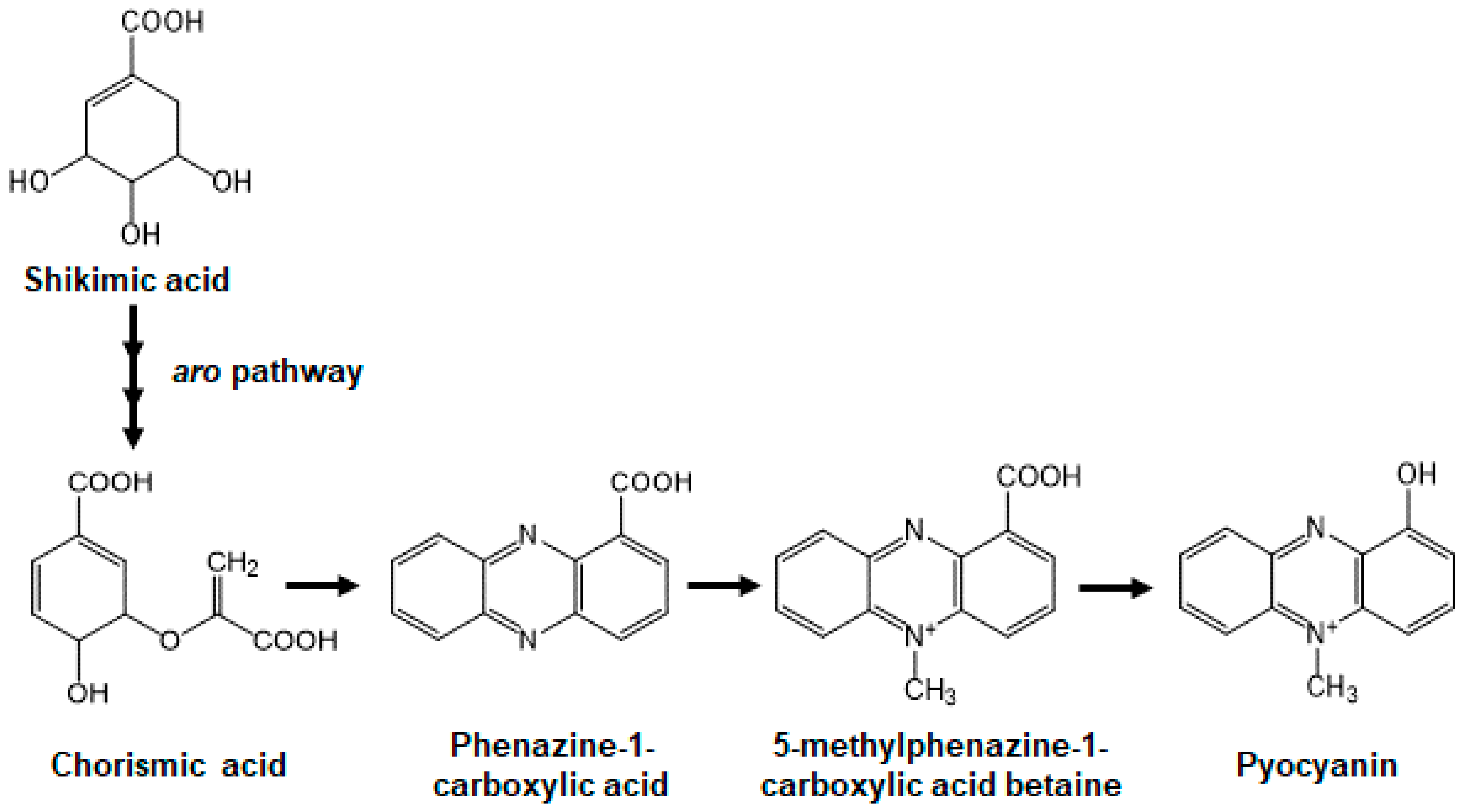
:1. Introduction
2. Results
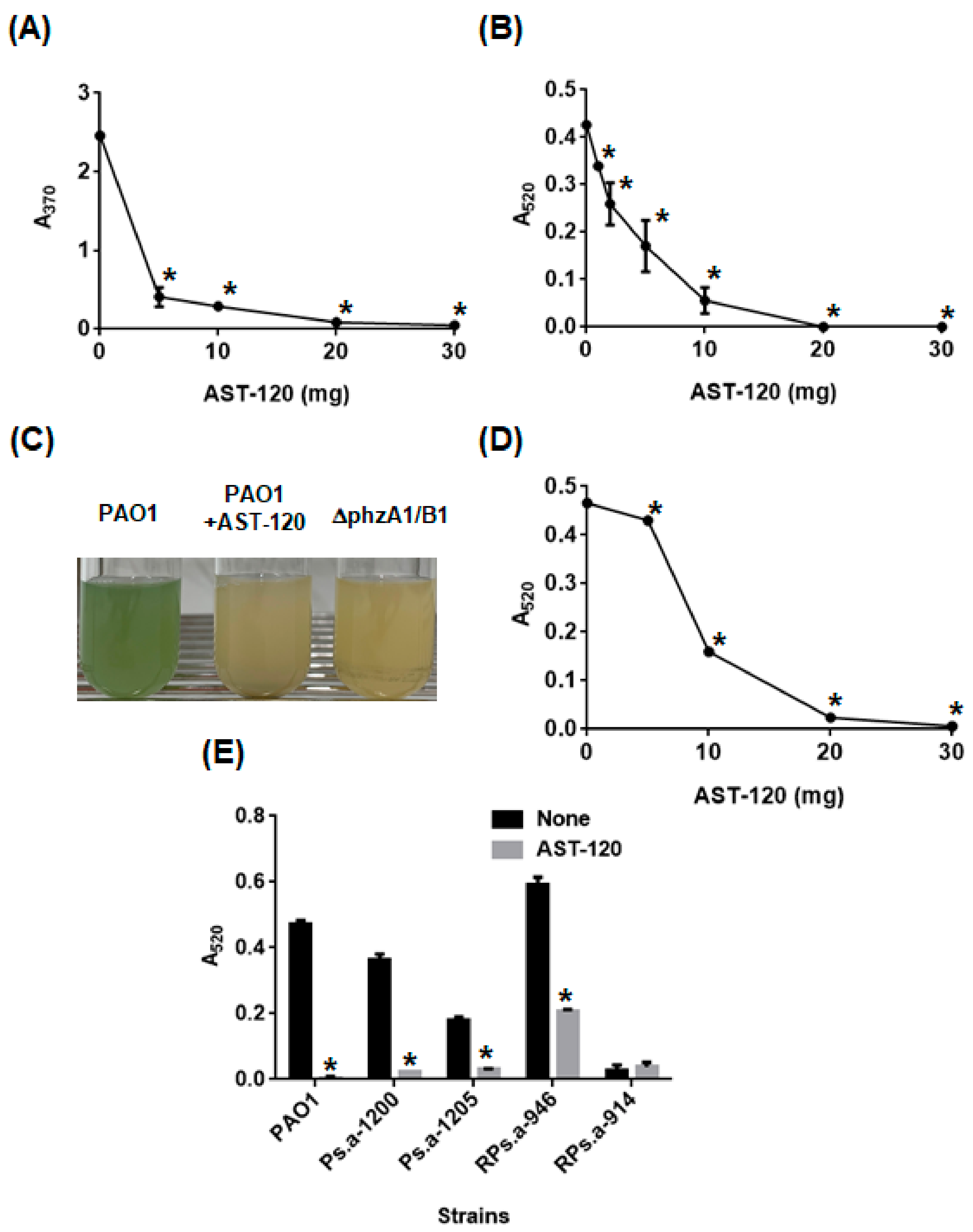
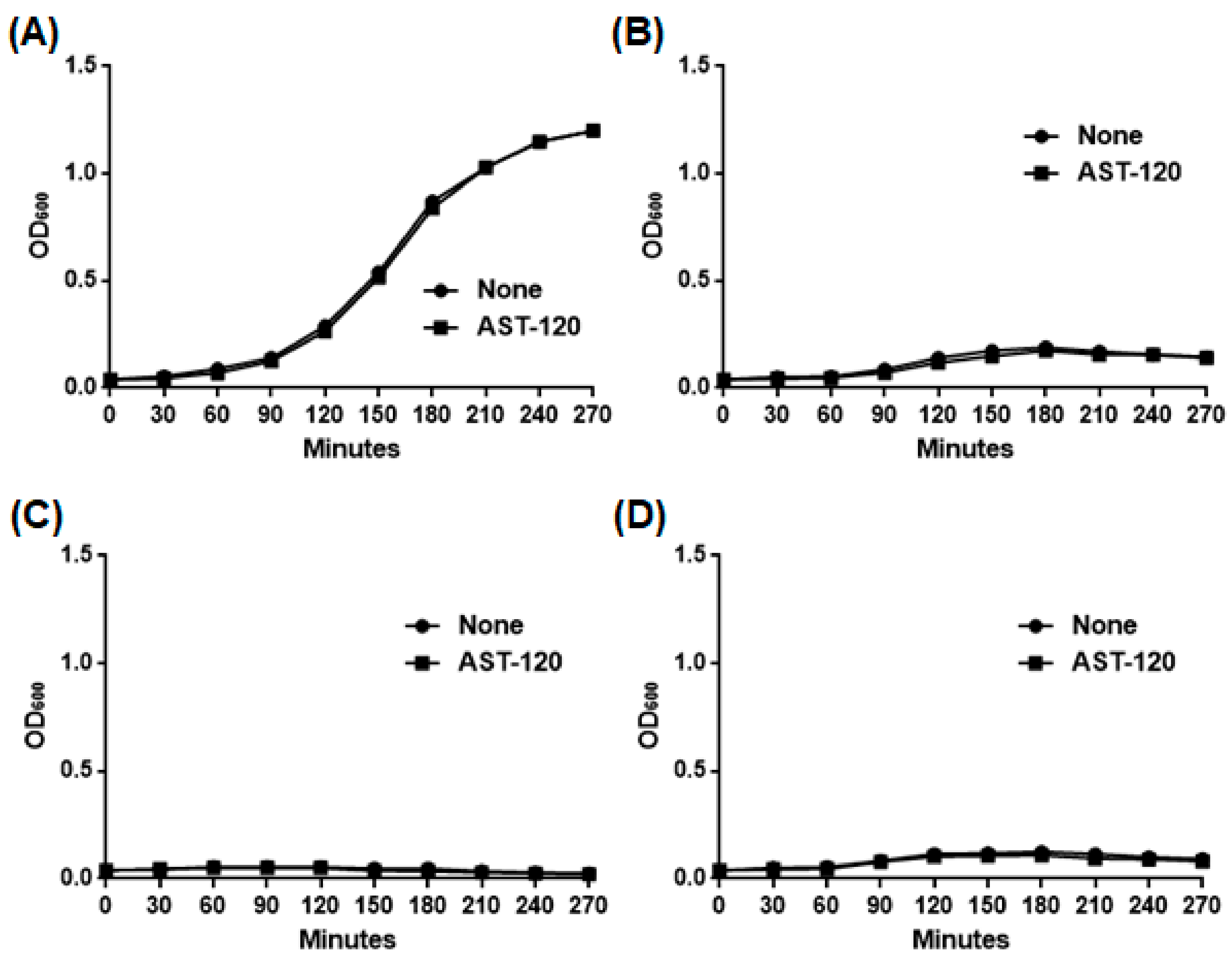
2.1. AST-120 Adsorbs Phenazines Produced by P. aeruginosa and Decreases the Extracellular Pyocyanin Levels
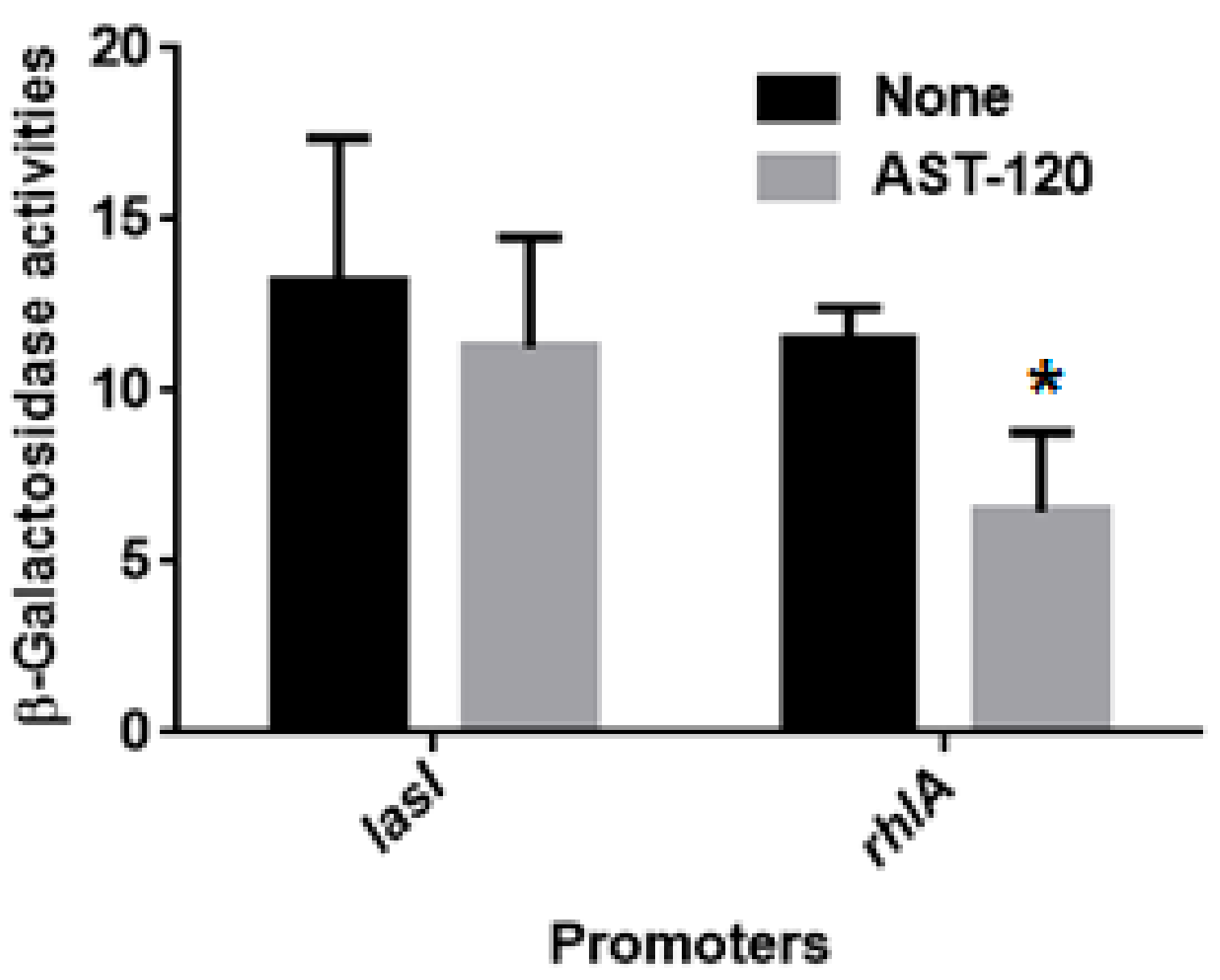
2.2. AST-120 Does Not Alter the Levels of LasI-LasR and RhlI-RhlR-Mediated Quorum Sensing
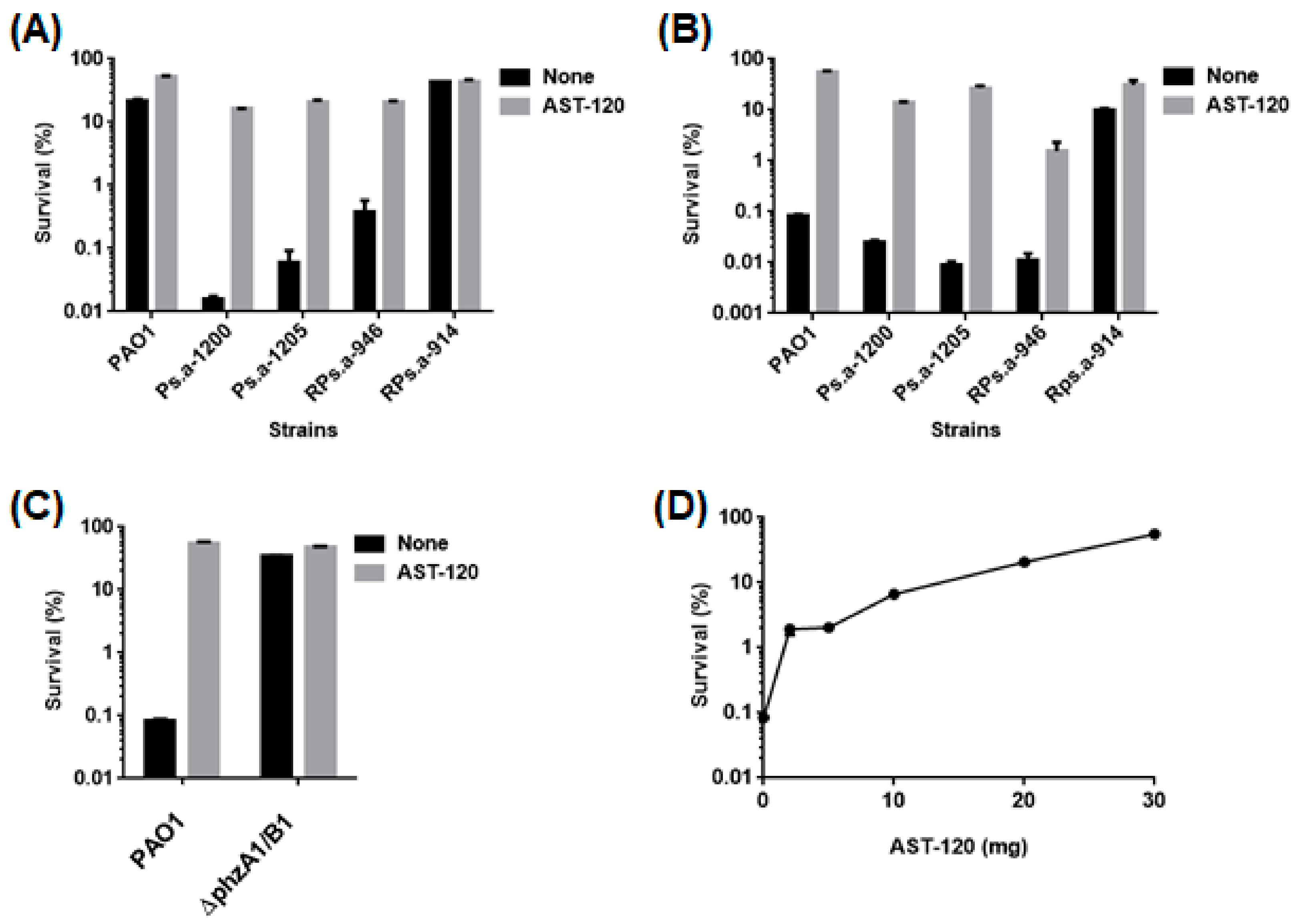
2.3. AST-120 Decreases Pyocyanin-Associated Toxicity of P. aeruginosa to Host Cells
2.4. AST-120 Does Not Impair Antibacterial Activities of Fosfomycin, Colistin and Amikacin
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Host Cells and Culture Conditions
4.2. Mutant Construction
4.3. Pyocyanin Assay
4.4. Reporter Construction and Promoter Assays
4.5. Cytotoxicity Assays
4.6. Minimum Inhibitory Concentration (MIC) Assays
4.7. Statistical Analysis
4.8. Approval for Experiments
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Murray, T.S.; Egan, M.; Kazmierczak, B.I. Pseudomonas aeruginosa chronic colonization in cystic fibrosis patients. Curr. Opin. Pediatr. 2007, 19, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Poletti, V.; Casoni, G.L.; Chilosi, M.; Zompatori, M. Diffuse panbronchiolitis. Eur. Respir. J. 2006, 28, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Turner, K.H.; Everett, J.; Trivedi, U.; Rumbaugh, K.P.; Whiteley, M. Requirements for Pseudomonas aeruginosa Acute Burn and Chronic Surgical Wound Infection. PLoS Genet. 2014, 10, e1004518. [Google Scholar] [CrossRef] [Green Version]
- Mittal, R.; Aggarwal, S.; Sharma, S.; Chhibber, S.; Harjai, K. Urinary tract infections caused by Pseudomonas aeruginosa: A minireview. J. Infect. Public Health 2009, 2, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Alverdy, J.; Holbrook, C.; Rocha, F.; Seiden, L.; Wu, R.L.; Musch, M.; Chang, E.; Ohman, D.; Suh, S. Gut-derived sepsis occurs when the right pathogen with the right virulence genes meets the right host: Evidence for in vivo virulence expression in Pseudomonas aeruginosa. Ann. Surg. 2000, 232, 480–489. [Google Scholar] [CrossRef]
- Poole, K. Pseudomonas aeruginosa: Resistance to the max. Front. Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef] [Green Version]
- Hoiby, N. Recent advances in the treatment of Pseudomonas aeruginosa infections in cystic fibrosis. BMC Med. 2011, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Aloush, V.; Navon-Venezia, S.; Seigman-Igra, Y.; Cabili, S.; Carmeli, Y. Multidrug-Resistant Pseudomonas aeruginosa: Risk Factors and Clinical Impact. Antimicrob. Agents Chemother. 2006, 50, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Obritsch, M.D.; Fish, D.N.; MacLaren, R.; Jung, R. National Surveillance of Antimicrobial Resistance in Pseudomonas aeruginosa Isolates Obtained from Intensive Care Unit Patients from 1993 to 2002. Antimicrob. Agents Chemother. 2004, 48, 4606–4610. [Google Scholar] [CrossRef] [Green Version]
- Mavrodi, D.V.; Bonsall, R.F.; Delaney, S.M.; Soule, M.J.; Phillips, G.; Thomashow, L.S. Functional Analysis of Genes for Biosynthesis of Pyocyanin and Phenazine-1-Carboxamide from Pseudomonas aeruginosa PAO1. J. Bacteriol. 2001, 183, 6454–6465. [Google Scholar] [CrossRef] [Green Version]
- Hall, S.; McDermott, C.; Anoopkumar-Dukie, S.; McFarland, A.J.; Forbes, A.; Perkins, A.V.; Davey, A.K.; Chess-Williams, R.; Kiefel, M.J.; Arora, D.; et al. Cellular Effects of Pyocyanin, a Secreted Virulence Factor of Pseudomonas aeruginosa. Toxins 2016, 8, 236. [Google Scholar] [CrossRef]
- Denning, G.M.; Railsback, M.A.; Rasmussen, G.T.; Cox, C.D.; Britigan, B.E. Pseudomonas pyocyanine alters calcium signaling in human airway epithelial cells. Am. J. Physiol. 1998, 274, L893–L900. [Google Scholar] [CrossRef]
- O’Malley, Y.Q.; Reszka, K.J.; Rasmussen, G.T.; Abdalla, M.Y.; Denning, G.M.; Britigan, B.E. The Pseudomonas secretory product pyocyanin inhibits catalase activity in human lung epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 285, L1077–L1086. [Google Scholar] [CrossRef] [Green Version]
- Ran, H.; Hassett, D.J.; Lau, G.W. Human targets of Pseudomonas aeruginosa pyocyanin. Proc. Natl. Acad. Sci. USA 2003, 100, 14315–14320. [Google Scholar] [CrossRef] [Green Version]
- Mahajan-Miklos, S.; Tan, M.-W.; Rahme, L.G.; Ausubel, F.M. Molecular Mechanisms of Bacterial Virulence Elucidated Using a Pseudomonas aeruginosa—Caenorhabditis elegans Pathogenesis Model. Cell 1999, 96, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Lau, G.W.; Goumnerov, B.C.; Walendziewicz, C.L.; Hewitson, J.; Xiao, W.; Mahajan-Miklos, S.; Tompkins, R.G.; Perkins, L.A.; Rahme, L.G. The Drosophila melanogaster Toll Pathway Participates in Resistance to Infection by the Gram-Negative Human Pathogen Pseudomonas aeruginosa. Infect. Immun. 2003, 71, 4059–4066. [Google Scholar] [CrossRef] [Green Version]
- Lau, G.W.; Ran, H.; Kong, F.; Hassett, D.J.; Mavrodi, D. Pseudomonas aeruginosa Pyocyanin Is Critical for Lung Infection in Mice. Infect. Immun. 2004, 72, 4275–4278. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Krishnan, G.; Goumnerov, B.; Tsongalis, J.; Tompkins, R.; Rahme, L.G. A quorum sensing-associated virulence gene of Pseudomonas aeruginosa encodes a LysR-like transcription regulator with a unique self-regulatory mechanism. Proc. Natl. Acad. Sci. USA 2001, 98, 14613–14618. [Google Scholar]
- Niwa, T. Role of Indoxyl Sulfate in the Progression of Chronic Kidney Disease and Cardiovascular Disease: Experimental and Clinical Effects of Oral Sorbent AST-120. Ther. Apher. Dial. 2011, 15, 120–124. [Google Scholar] [CrossRef]
- Hirakawa, H.; Uchida, M.; Kurabayashi, K.; Nishijima, F.; Takita, A.; Tomita, H. In vitro activity of AST-120 that suppresses indole signaling in Escherichia coli, which attenuates drug tolerance and virulence. PLoS ONE 2020, 15, e0232461. [Google Scholar]
- Costa, K.C.; Bergkessel, M.; Saunders, S.; Korlach, J.; Newman, D.K. Enzymatic Degradation of Phenazines Can Generate Energy and Protect Sensitive Organisms from Toxicity. mBio 2015, 6, e01520-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, M.; Lostroh, C.P.; Ogi, T.; Greenberg, E.P. Identification, Timing, and Signal Specificity of Pseudomonas aeruginosa Quorum-Controlled Genes: A Transcriptome Analysis. J. Bacteriol. 2003, 185, 2066–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, X.; Thouverez, M.; Talon, D.; Boillot, A.; Capellier, G.; Floriot, C.; Hélias, J.P. Endemicity, molecular diversity and colonisation routes of Pseudomonas aeruginosa in intensive care units. Intensive Care Med. 2001, 27, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Voggu, L.; Schlag, S.; Biswas, R.; Rosenstein, R.; Rausch, C.; Götz, F. Microevolution of Cytochrome bd Oxidase in Staphylococci and Its Implication in Resistance to Respiratory Toxins Released by Pseudomonas. J. Bacteriol. 2006, 188, 8079–8086. [Google Scholar] [CrossRef] [Green Version]
- Christofi, T.; Panayidou, S.; Dieronitou, I.; Michael, C.; Apidianakis, Y. Metabolic output defines Escherichia coli as a health-promoting microbe against intestinal Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Glasser, N.R.; Kern, S.E.; Newman, D.K. Phenazine redox cycling enhances anaerobic survival in Pseudomonas aeruginosa by facilitating generation of ATP and a proton-motive force. Mol. Microbiol. 2014, 92, 399–412. [Google Scholar]
- Wang, Y.; Wilks, J.C.; Danhorn, T.; Ramos, I.; Croal, L.; Newman, D.K. Phenazine-1-Carboxylic Acid Promotes Bacterial Biofilm Development via Ferrous Iron Acquisition. J. Bacteriol. 2011, 193, 3606–3617. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.; Sykes, D.A.; Watson, D.; Rutman, A.; Taylor, G.W.; Cole, P.J. Measurement of Pseudomonas aeruginosa phenazine pigments in sputum and assessment of their contribution to sputum sol toxicity for respiratory epithelium. Infect. Immun. 1988, 56, 2515–2517. [Google Scholar] [CrossRef] [Green Version]
- Wood, L.G.; Fitzgerald, D.A.; Lee, A.K.; Garg, M.L. Improved antioxidant and fatty acid status of patients with cystic fibrosis after antioxidant supplementation is linked to improved lung function1–3. Am. J. Clin. Nutr. 2003, 77, 150–159. [Google Scholar] [CrossRef]
- Lau, G.W.; Hassett, D.J.; Ran, H.; Kong, F. The role of pyocyanin in Pseudomonas aeruginosa infection. Trends Mol. Med. 2004, 10, 599–606. [Google Scholar] [CrossRef]
- Hirakawa, H.; Tomita, H. Interference of bacterial cell-to-cell communication: A new concept of antimicrobial chemotherapy breaks antibiotic resistance. Front. Microbiol. 2013, 4, 114. [Google Scholar] [CrossRef] [Green Version]
- Michalopoulos, A.S.; Livaditis, I.G.; Gougoutas, V. The revival of fosfomycin. Int. J. Infect. Dis. 2011, 15, e732–e739. [Google Scholar] [CrossRef] [Green Version]
- Montero, M.; Horcajada, J.P.; Sorlí, L.; Alvarez-Lerma, F.; Grau, S.; Riu, M.; Sala, M.; Knobel, H. Effectiveness and safety of colistin for the treatment of multidrug-resistant Pseudomonas aeruginosa infections. Infection 2009, 37, 461–465. [Google Scholar] [CrossRef]
- Chyan, Y.J.; Poeggeler, B.; Omar, R.A.; Chain, D.G.; Frangione, B.; Ghiso, J.; Pappolla, M.A. Potent neuroprotective properties against the Alzheimer beta-amyloid by an endogenous melatonin-related indole structure, indole-3-propionic acid. J. Biol. Chem. 1999, 274, 21937–21942. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, M.; Mukherjee, S.; Wang, H.; Li, H.; Sun, K.; Benechet, A.P.; Qiu, Z.; Maher, L.; Redinbo, M.R.; Phillips, R.S.; et al. Symbiotic Bacterial Metabolites Regulate Gastrointestinal Barrier Function via the Xenobiotic Sensor PXR and Toll-like Receptor 4. Immunity 2014, 41, 296–310. [Google Scholar] [CrossRef] [Green Version]
- Swimm, A.; Giver, C.R.; DeFilipp, Z.; Rangaraju, S.; Sharma, A.; Antonova, A.U.; Sonowal, R.; Capaldo, C.; Powell, D.; Qayed, M.; et al. Indoles derived from intestinal microbiota act via type I interferon signaling to limit graft-versus-host disease. Blood 2018, 132, 2506–2519. [Google Scholar] [CrossRef] [Green Version]
- Asai, M.; Kumakura, S.; Kikuchi, M. Review of the efficacy of AST-120 (KREMEZIN((R))) on renal function in chronic kidney disease patients. Ren. Fail. 2019, 41, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, H.; Kurabayashi, K.; Tanimoto, K.; Tomita, H. Oxygen Limitation Enhances the Antimicrobial Activity of Fosfomycin in Pseudomonas aeruginosa Following Overexpression of glpT Which Encodes Glycerol-3-Phosphate/Fosfomycin Symporter. Front. Microbiol. 2018, 9, 1950. [Google Scholar] [CrossRef] [Green Version]
- Hoang, T.T.; Karkhoff-Schweizer, R.R.; Kutchma, A.J.; Schweizer, H.P. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: Application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 1998, 212, 77–86. [Google Scholar] [CrossRef]
- Essar, D.W.; Eberly, L.; Hadero, A.; Crawford, I.P. Identification and characterization of genes for a second anthranilate synthase in Pseudomonas aeruginosa: Interchangeability of the two anthranilate synthases and evolutionary implications. J. Bacteriol. 1990, 172, 884–900. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | DNA Sequence (5′–3′) | Use |
|---|---|---|
| phzA1B1-delta1 phzA1B1-delta2 phzA1B1-delta3 phzA1B1-delta4 lasI-PF | gcgggatccctacaacctccggcattgc cttccaggatggcctcaggtgggaccgttcatgcgccgcctc ctcggaggcggcgcatgaacggtcccacctgaggccatcctg gcgaagcttcggcgctgttcggcatggg gcgatgcattcttgcctctcaggtcggc | phzA1 and phzB1 deletion construction phzA1 and phzB1 deletion construction phzA1 and phzB1 deletion construction phzA1 and phzB1 deletion construction pBBRlasI (PAO1)-P construction |
| lasI-PR rhlA-PF | gcgggtacccttcacttcctccaaatagg gcgatgcatcgccagagcgtttcgacac | pBBRlasI (PAO1)-P construction pBBRrhlA (PAO1)-P construction |
| rhlA-PR | gcgggtaccttcacacctcccaaaaattttcg | pBBRrhlA (PAO1)-P construction |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirakawa, H.; Takita, A.; Uchida, M.; Kaneko, Y.; Kakishima, Y.; Tanimoto, K.; Kamitani, W.; Tomita, H. Adsorption of Phenazines Produced by Pseudomonas aeruginosa Using AST-120 Decreases Pyocyanin-Associated Cytotoxicity. Antibiotics 2021, 10, 434. https://doi.org/10.3390/antibiotics10040434
Hirakawa H, Takita A, Uchida M, Kaneko Y, Kakishima Y, Tanimoto K, Kamitani W, Tomita H. Adsorption of Phenazines Produced by Pseudomonas aeruginosa Using AST-120 Decreases Pyocyanin-Associated Cytotoxicity. Antibiotics. 2021; 10(4):434. https://doi.org/10.3390/antibiotics10040434
Chicago/Turabian StyleHirakawa, Hidetada, Ayako Takita, Motoyuki Uchida, Yuka Kaneko, Yuto Kakishima, Koichi Tanimoto, Wataru Kamitani, and Haruyoshi Tomita. 2021. "Adsorption of Phenazines Produced by Pseudomonas aeruginosa Using AST-120 Decreases Pyocyanin-Associated Cytotoxicity" Antibiotics 10, no. 4: 434. https://doi.org/10.3390/antibiotics10040434
APA StyleHirakawa, H., Takita, A., Uchida, M., Kaneko, Y., Kakishima, Y., Tanimoto, K., Kamitani, W., & Tomita, H. (2021). Adsorption of Phenazines Produced by Pseudomonas aeruginosa Using AST-120 Decreases Pyocyanin-Associated Cytotoxicity. Antibiotics, 10(4), 434. https://doi.org/10.3390/antibiotics10040434