
Draft Genome Sequence and Biofilm Production of a Carbapenemase-Producing Klebsiella pneumoniae (KpR405) Sequence Type 405 Strain Isolated in Italy



 ,
,
Abstract
:1. Introduction
2. Results
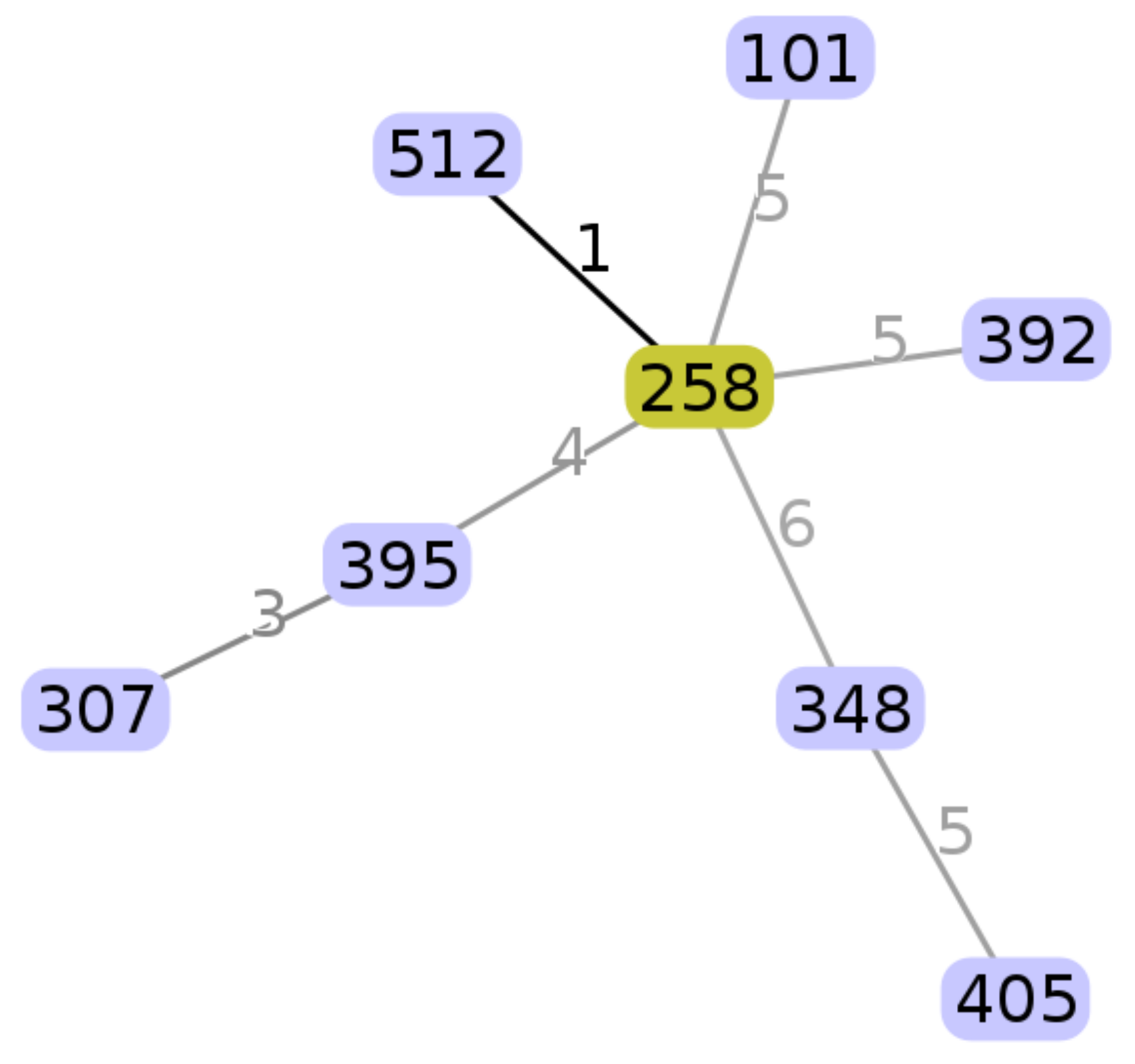
2.1. Molecular Genotyping
2.2. Biofilm Formation Detection
3. Discussion
4. Materials and Methods
4.1. Patient Information, Bacterial Isolation, and Identification
4.2. Molecular Genotyping
4.3. Quantitative Biofilm Production Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Podschun, R.; Ullmann, U. Klebsiella spp. as Nosocomial Pathogens: Epidemiology, Taxonomy, Typing Methods, and Pathogenicity Factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broberg, C.A.; Palacios, M.; Miller, V.L. Klebsiella: A long way to go towards understanding this enigmatic jet-setter. F1000Prime Rep. 2014, 6, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derakhshan, S.; Peerayeh, S.N.; Bakhshi, B. Association Between Presence of Virulence Genes and Antibiotic Resistance in ClinicalKlebsiella PneumoniaeIsolates. Lab. Med. 2016, 47, 306–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitout, J.D.; Nordmann, P.; Poirel, L. Carbapenemase-Producing Klebsiella pneumoniae, a Key Pathogen Set for Global Nos-ocomial Dominance. Antimicrob. Agents Chemother. 2015, 29, 5873–5874. [Google Scholar] [CrossRef] [Green Version]
- Muggeo, A.; Guillard, T.; Klein, F.; Reffuveille, F.; François, C.; Babosan, A.; Bajolet, O.; Bertrand, X.; de Champs, C. Spread of Klebsiella pneumoniae ST395 non-susceptible to carbapenems and re-sistant to fluoroquinolones in North-Eastern France. J. Glob. Antimicrob. Resist. 2018, 13, 98–103. [Google Scholar] [CrossRef]
- Andrade, L.N.; Curiao, T.; Ferreira, J.C.; Longo, J.M.; Clímaco, E.C.; Martinez, R.; Bellissimo-Rodrigues, F.; Basile-Filho, A.; Evaristo, M.A.; Del Peloso, P.F.; et al. Dissemination of blaKPC-2 by the Spread of Klebsiella pneumoniae Clonal Complex 258 Clones (ST258, ST11, ST437) and Plasmids (IncFII, IncN, IncL/M) among Enterobacteriaceae Species in Brazil. Antimicrob. Agents Chemother. 2011, 55, 3579–3583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geraci, D.; Bonura, C.; Giuffrè, M.; Saporito, L.; Graziano, G.; Aleo, A.; Fasciana, T.; Di Bernardo, F.; Stampone, T.; Palma, D.; et al. Is the monoclonal spread of the ST258, KPC-3-producing clone being replaced in southern Italy by the dissemination of multiple clones of carbapenem-nonsusceptible, KPC-3-producing Klebsiella pneumoniae? Clin. Microbiol. Infect. 2015, 21, e15–e17. [Google Scholar] [CrossRef] [Green Version]
- Bonura, C.; Giuffrè, M.; Aleo, A.; Fasciana, T.; Di Bernardo, F.; Stampone, T.; Giammanco, A.; Palma, D.M.; Mammina, C. The MDR-GN Working Group an Update of the Evolving Epidemic of blaKPC Carrying Klebsiella pneumoniae in Sicily, Italy, 2014: Emergence of Multiple Non-ST258 Clones. PLoS ONE 2015, 10, e0132936. [Google Scholar] [CrossRef] [Green Version]
- Mammina, C.; Bonura, C.; Di Bernardo, F.; Aleo, A.; Fasciana, T.; Sodano, C.; Saporito, M.A.; Verde, M.S.; Tetamo, R.; Palma, D.M. Ongoing spread of colistin-resistant Klebsiella pneumoniae in different wards of an acute general hospital, Italy, June to December 2011. Euro Surveill. 2012, 17, 20248. [Google Scholar] [CrossRef]
- Bengoechea, J.A.; Pessoa, J.S. Klebsiella pneumoniae infection biology: Living to counteract host defences. FEMS Microbiol. Rev. 2019, 43, 123–144. [Google Scholar] [CrossRef] [Green Version]
- Mammina, C.; Bonura, C.; Aleo, A.; Fasciana, T.; Brunelli, T.; Pesavento, G.; Degl’Innocenti, R.; Nastasi, A. Sequence type 101 (ST101) as the predominant carbapenem-non-susceptible Klebsiella pneumoniae clone in an acute general hospital in Italy. Int. J. Antimicrob. Agents 2012, 39, 543–545. [Google Scholar] [CrossRef] [PubMed]
- Gona, F.; Barbera, F.; Pasquariello, A.C.; Grossi, P.; Gridelli, B.; Mezzatesta, M.L.; Caio, C.; Stefani, S.; Conaldi, P.G. In vivo multiclonal transfer of blaKPC-3 from Klebsiella pneumoniae to Escherichia coli in surgery patients. Clin. Microbiol. Infect. 2014, 20, O633–O635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasciana, T.; Gentile, B.; Aquilina, M.; Ciammaruconi, A.; Mascarella, C.; Anselmo, A.; Fortunato, A.; Fillo, S.; Petralito, G.; Lista, F.; et al. Co-existence of virulence factors and antibiotic resistance in new Klebsiella pneumoniae clones emerging in south of Italy. BMC Infect. Dis. 2019, 19, 2019. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: New A Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubero, M.; Marti, S.; Domínguez, M.Á.; González-Díaz, A.; Berbel, D.; Ardanuy, C. Hy-pervirulent Klebsiella pneumoniae serotype K1 clinical isolates form robust biofilms at the air-liquid interface. PLoS ONE 2019, 14, e0222628. [Google Scholar] [CrossRef]
- Huynh, D.T.N.; Kim, A.Y.; Kim, Y.R. Identification of Pathogenic Factors in Klebsiella pneumoni-ae Using Impedimetric Sensor Equipped with Biomimetic Surfaces. Sensors 2017, 17, 1406. [Google Scholar] [CrossRef]
- Nirwati, H.; Sinanjung, K.; Fahrunissa, F.; Wijaya, F.; Napitupulu, S.; Hati, V.P.; Hakim, M.S.; Meliala, A.; Aman, A.T.; Nuryastuti, T. Biofilm formation and antibiotic resistance of Klebsiella pneumoniae isolated from clinical samples in a tertiary care hospital, Klaten, Indonesia. BMC Proc. 2019, 13 (Suppl. 11), 20. [Google Scholar] [CrossRef]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.; Phan, H.T.; Findlay, J.; Stoesser, N.; Pankhurst, L.; Navickaite, I.; De Maio, N.; Eyre, D.W.; Toogood, G.; Orsi, N.M.; et al. Covert dissemination of car-bapenemase-producing Klebsiella pneumoniae (KPC) in a successfully controlled outbreak: Long- and short-read whole-genome sequencing demonstrate multiple genetic modes of transmission. J. Antimicrob. Chemother. 2017, 72, 3025–3034. [Google Scholar] [CrossRef]
- Rimoldi, S.G.; Gentile, B.; Pagani, C.; Di Gregorio, A.; Anselmo, A.; Palozzi, A.M.; Fortunato, A.; Pittiglio, V.; Ridolfo, A.L.; Gismondo, M.R.; et al. Whole genome sequencing for the molecular characterization of carbapenem-resistant Klebsiella pneumoniae strains iso-lated at the Italian ASST Fatebenefratelli Sacco Hospital, 2012–2014. BMC Infect. Dis. 2017, 17, 666. [Google Scholar] [CrossRef]
- Gentile, B.; Grottola, A.; Orlando, G.; Serpini, G.F.; Venturelli, C.; Meschiari, M.; Anselmo, A.; Fillo, S.; Fortunato, A.; Lista, F.; et al. A Retrospective Whole-Genome Sequencing Analysis of Carbapenem and Colistin-Resistant Klebsiella pneumoniae Nosocomial Strains Isolated during an MDR Surveillance Program. Antibiotics 2020, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Tumbarello, M.; Trecarichi, E.M.; De Rosa, F.G.; Giannella, M.; Giacobbe, D.R.; Bassetti, M.; Losito, A.R.; Bartoletti, M.; Del Bono, V.; Corcione, S.; et al. Infections caused by KPC-producing Klebsiella pneumoniae: Differences in therapy and mortalityin a multicentre study. J. Antimicrob. Chemother. 2015, 70, 2133–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daikos, G.L.; Tsaousi, S.; Tzouvelekis, L.S.; Anyfantis, I.; Psichogiou, M.; Argyropoulou, A.; Stefanou, I.; Sypsa, V.; Miriagou, V.; Nepka, M.; et al. Carbapenemase-Producing Klebsiella pneumoniae Bloodstream Infections: Lowering Mortality by Antibiotic Combination Schemes and the Role of Carbapenems. Antimicrob. Agents Chemother. 2014, 58, 2322–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zusman, O.; Avni, T.; Leibovici, L.; Adler, A.; Friberg, L.; Stergiopoulou, T.; Carmeli, Y.; Paul, M. Systematic review and me-ta-analysis of in vitro synergy of polymyxins and carbapenems. Antimicrob. Agents Chemother. 2013, 57, 5104–5111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mataraci Kara, E.; Ozbek Celik, B. Investigation of the effects of various antibiotics against Klebsiella pneu-moniae biofilms on in vitro catheter model. J. Chemother. 2018, 30, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, P.; Gulotta, G.; Casuccio, A.; Pantuso, G.; Raineri, M.; Farulla, C.A.; Bonventre, S.; Guadagnino, G.; Ingrassia, D.; Cocorullo, G.; et al. KPC—3 Klebsiella pneumoniaeST258 clone infection in postoperative abdominal surgery patients in an intensive care setting: Analysis of a case series of 30 patients. BMC Anesth. 2013, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Brhelova, E.; Antonova, M.; Pardy, F.; Kocmanova, I.; Mayer, J.; Racil, Z.; Lengerova, M. Investigation of the next-generation sequencing data of Klebsiella pneumoniae using web-based tools. J. Med. Microbiol. 2017, 66, 1673–1683. [Google Scholar] [CrossRef]
- Francisco, A.P.; Bugalho, M.; Ramirez, M.; Carrico, J.A. Global Optimal eBURST analysis of Multilocus typing data using a raphic matroid approach. BMC Bioinform. 2009, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Francisco, A.P.; Vaz, C.; Monteiro, P.T.; Melo-Cristino, J.; Ramirez, M.; Carriço, J.A. PHYLOViZ: Phylogenetic inference and data visualization for sequence based typing methods. BMC Bioinform. 2012, 13, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calà, C.; Amodio, E.; Di Carlo, E.; Virruso, R.; Fasciana, T.; Giammanco, A. Biofilm production in Staphylococcus epidermidis strains, isolated from the skin of hospitalized patients: Genetic and phenotypic characteristics. New Microbiol. 2015, 38, 521–529. [Google Scholar]
- Vuotto, C.; Longo, F.; Pascolini, C.; Donelli, G.; Balice, M.P.; Libori, M.F.; Tiracchia, V.; Salvia, A.; Varaldo, P.E. Varaldo. Biofilm formation and antibiotic resistance in Klebsiella pneumoniae urinary strains. J. Appl. Microbiol. 2017, 123, 1003–1018. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Locus | Putative Function of Gene | Lengh (bp) | ST 258 | ST 512 | ST 395 | ST 101 | ST 392 | ST 348 | ST 307 | ST 405 |
|---|---|---|---|---|---|---|---|---|---|---|
| rpoB | beta subunit of RNA polymerase B | 501 | 1 | 1 | 1 | 1 | 4 | 15 | 1 | 4 |
| gap A | Glyceraldehyde 3-phoshate dehydrogenase | 450 | 3 | 54 | 3 | 2 | 3 | 2 | 4 | 2 |
| mdh | Malate deydrogenase | 477 | 1 | 1 | 2 | 1 | 6 | 20 | 2 | 62 |
| Pgi | Phosphoglucose isomerase | 432 | 1 | 1 | 4 | 5 | 1 | 10 | 52 | 3 |
| pho E | Phosphoporine E | 420 | 1 | 1 | 1 | 4 | 7 | 12 | 1 | 10 |
| inf B | Translation initiation factor 2 | 318 | 3 | 3 | 1 | 6 | 4 | 1 | 1 | 1 |
| ton B | Periplasmic energy trasducer | 414 | 79 | 79 | 4 | 6 | 40 | 16 | 7 | 110 |
| Resistance Genes | ||||||
|---|---|---|---|---|---|---|
| Beta-Lactamase | Aminoglycoside | Heavy Metal | Quinolone | Efflux Systems and Regulator Systems | ||
| Gene alleles | blaKPC-3,blaCTX-M-15, blaOXA-1, blaSHV-76, blaTEM-1 | aac3 lla-c2, aac6p Ib-b-cr, aph3pp Ib-2, aph6 Id-1 | pcoA-2, pcoB-3, pcoC-1, pcoD-2, pcoE-1, pcoR-1, pcoS-2, silC-3, silE-3, silR-2, silS-2 | gyrA-4, gyrB-1, parC-2, qnrB-1 | acrR-1, envR-15, fis-1, marA-2, marR-1, oqxR-4, rob-21, sdiA-6, soxR-2, soxS-4 | |
| Virulence Factors | ||||||
| Gene alleles | type-3 fimbriae, biofilm formation, host cell adherence/capsule | yersiniabactin receptor | aerobactin/regulator system | yersiniabactin system | microcin E492 system components | |
| Gene alleles | mrkA-4, mrkB-1, mrkF-38, rkH-15, mrkI-18,mrkJ-1/wzi-143, wzc-937 | fyuA | irp1-148, irp2-145/kvgA-2 | ybtA-1, ybtA-39, ybtE-69, ybtP-4, ybtQ-60, ybtS-6, ybtT-39, ybtU-2, ybtX-62 | mceA-1, mceC-1, mceD-3, mceE-2, mceH-5 | |
| OD 570 | |
|---|---|
| Negative control (median values) | 0.059 |
| Standard deviation | 0.005 |
| Cut-off (median values) | 0.071 |
| K. pneumoniae ST405 (median values) | 0.215 |
| ATCC 700603 (median values) | 0.765 |
| ATCC 13883 (median values) | 0.102 |
| Antibiotics | MIC Values µg/mL |
|---|---|
| IMP | >8 |
| MEP | >8 |
| ETP | >1 |
| CIP | >1 |
| AUG | >8/2 |
| CXM | >8 |
| CTX | >4 |
| CAZ | >8 |
| FEP | >8 |
| SXT | >4/76 |
| CN | >4 |
| ATM | >16 |
| TZP | >16/4 |
| FOS | 32 |
| TGC | 2 |
| CS | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fasciana, T.; Ciammaruconi, A.; Gentile, B.; Di Carlo, P.; Virruso, R.; Tricoli, M.R.; Palma, D.M.; Pitarresi, G.L.; Lista, F.; Giammanco, A. Draft Genome Sequence and Biofilm Production of a Carbapenemase-Producing Klebsiella pneumoniae (KpR405) Sequence Type 405 Strain Isolated in Italy. Antibiotics 2021, 10, 560. https://doi.org/10.3390/antibiotics10050560
Fasciana T, Ciammaruconi A, Gentile B, Di Carlo P, Virruso R, Tricoli MR, Palma DM, Pitarresi GL, Lista F, Giammanco A. Draft Genome Sequence and Biofilm Production of a Carbapenemase-Producing Klebsiella pneumoniae (KpR405) Sequence Type 405 Strain Isolated in Italy. Antibiotics. 2021; 10(5):560. https://doi.org/10.3390/antibiotics10050560
Chicago/Turabian StyleFasciana, Teresa, Andrea Ciammaruconi, Bernardina Gentile, Paola Di Carlo, Roberta Virruso, Maria Rita Tricoli, Daniela Maria Palma, Giovanna Laura Pitarresi, Florigio Lista, and Anna Giammanco. 2021. "Draft Genome Sequence and Biofilm Production of a Carbapenemase-Producing Klebsiella pneumoniae (KpR405) Sequence Type 405 Strain Isolated in Italy" Antibiotics 10, no. 5: 560. https://doi.org/10.3390/antibiotics10050560
APA StyleFasciana, T., Ciammaruconi, A., Gentile, B., Di Carlo, P., Virruso, R., Tricoli, M. R., Palma, D. M., Pitarresi, G. L., Lista, F., & Giammanco, A. (2021). Draft Genome Sequence and Biofilm Production of a Carbapenemase-Producing Klebsiella pneumoniae (KpR405) Sequence Type 405 Strain Isolated in Italy. Antibiotics, 10(5), 560. https://doi.org/10.3390/antibiotics10050560