
Combined Effects of Sulfamethoxazole and Erythromycin on a Freshwater Microalga, Raphidocelis subcapitata: Toxicity and Oxidative Stress




Abstract
:
1. Introduction
2. Results and Discussion
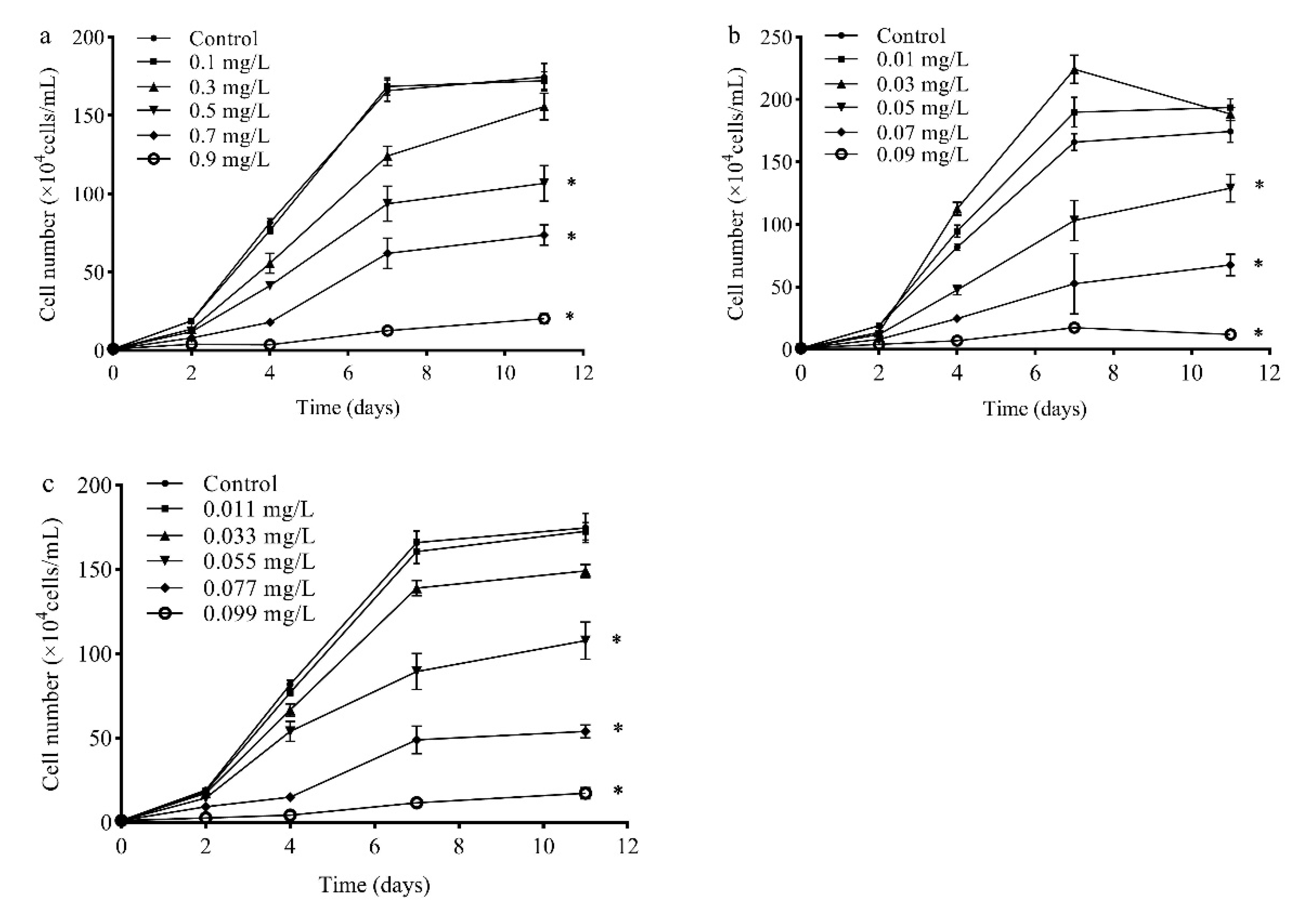
2.1. Toxicity of SMZ, SMX, and Their Mixture
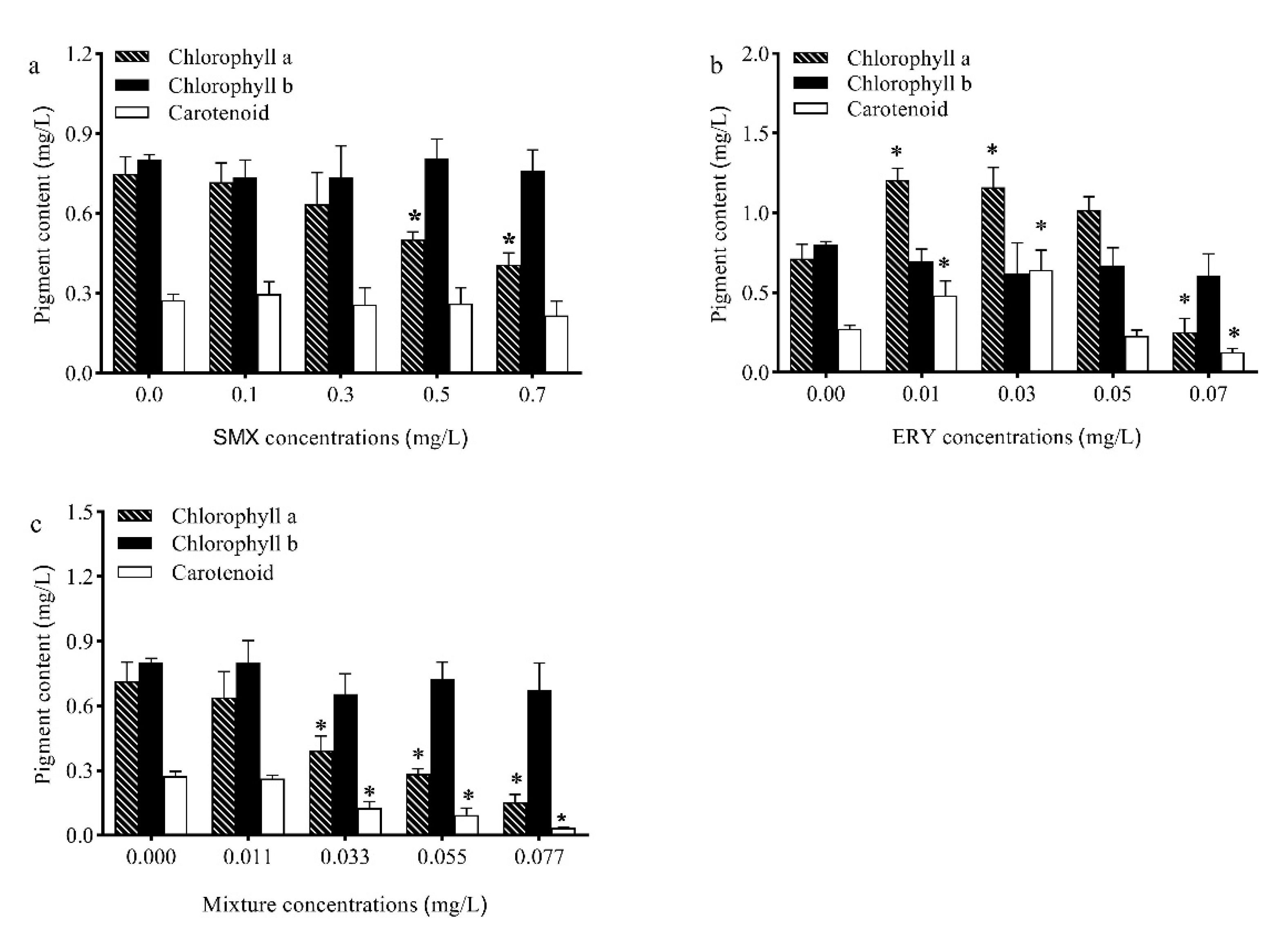
2.2. Effects of Antibiotics on Pigments
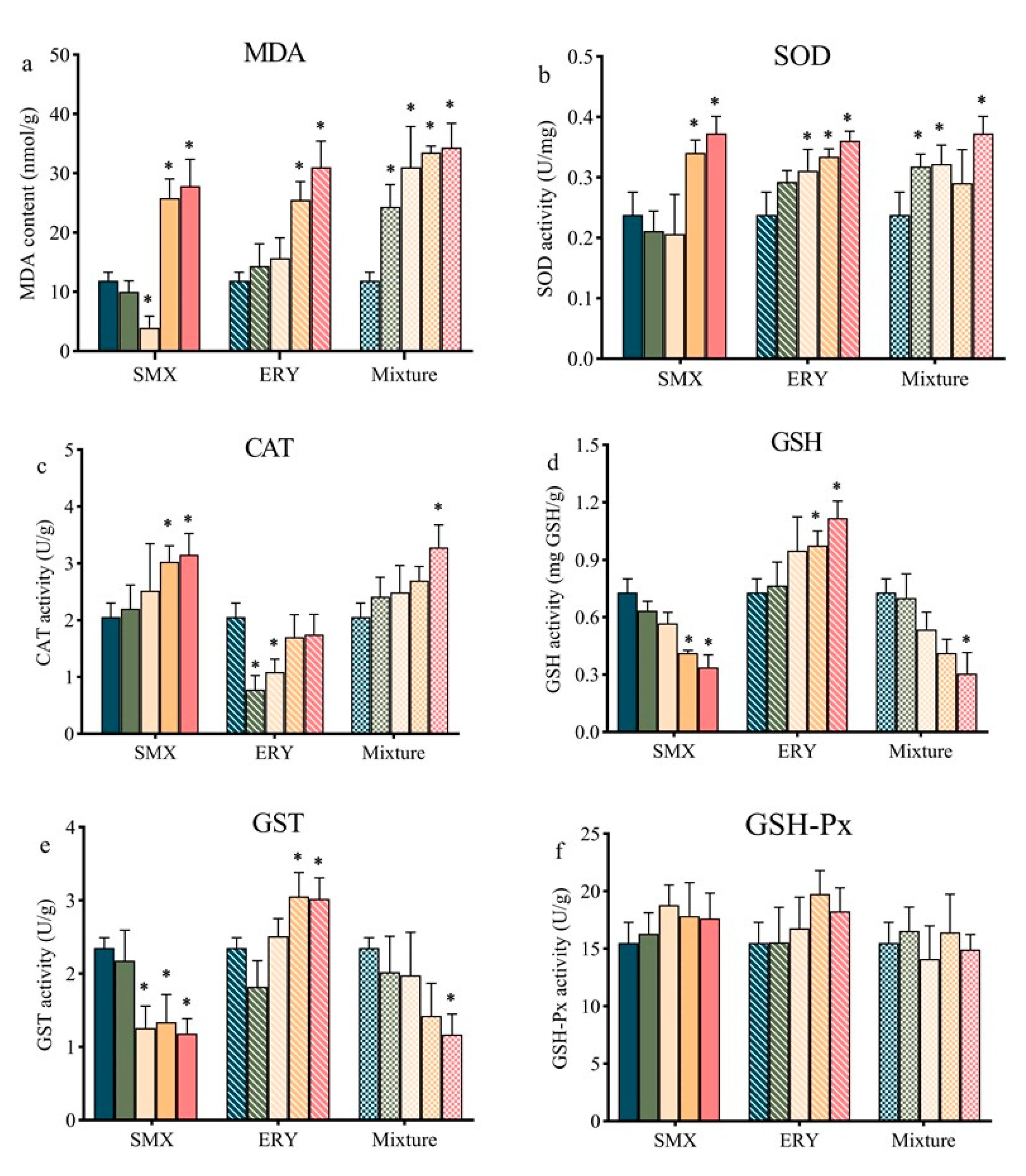
2.3. Effects of Antibiotics on Antioxidant Responses
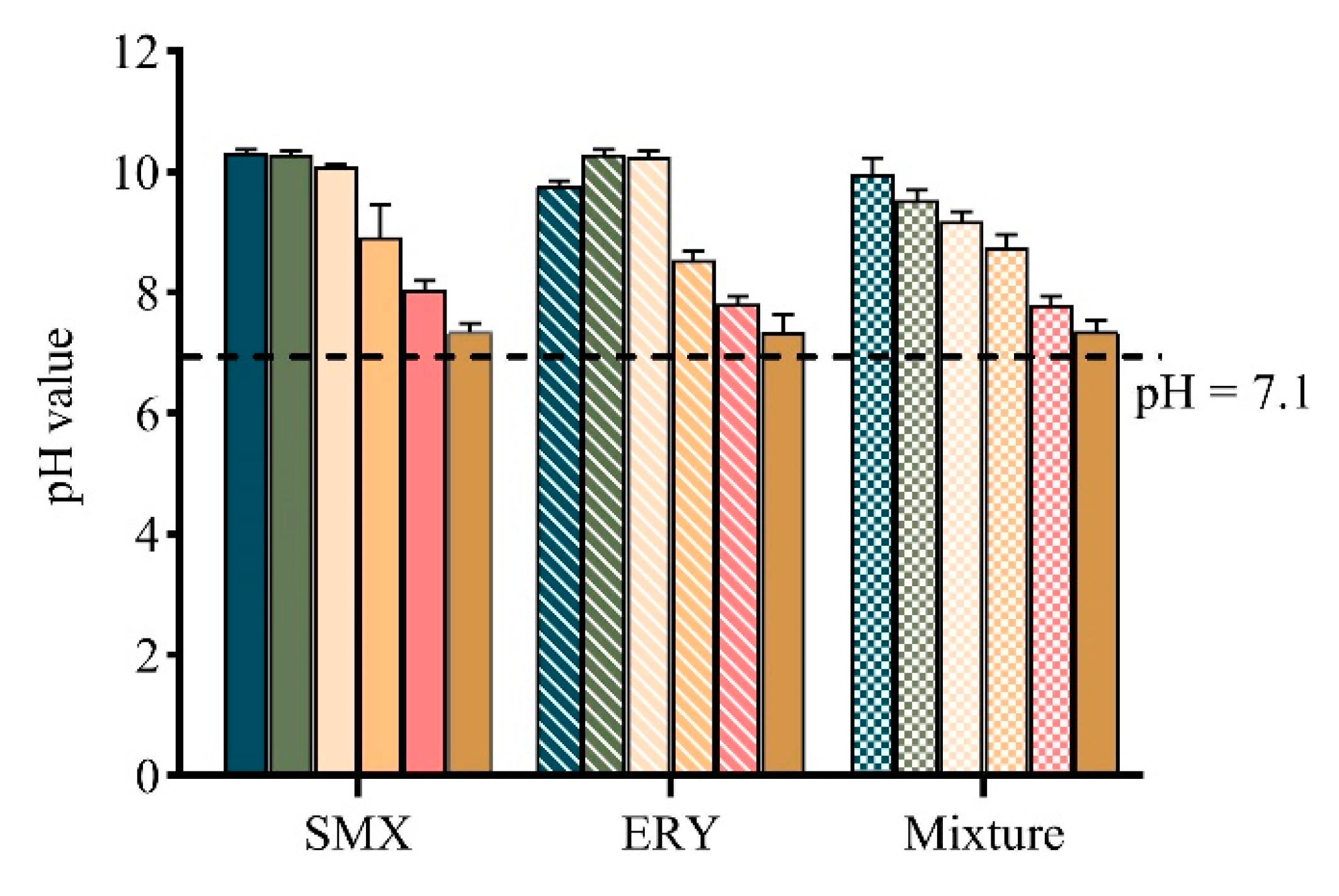
2.4. pH Value Change
3. Materials and Methods
3.1. Test Algae and Culture
3.2. Antibiotics
3.3. Procedures for the Growth Inhibition and Ecotoxicological Risk Assessment
3.4. Photosynthetic Pigment Content
3.5. Analysis of Antioxidant Responses
3.6. Antibiotic Analyses
3.7. Statistics Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Borecka, M.; Bialk-Bielinska, A.; Halinski, L.P.; Pazdro, K.; Stepnowski, P.; Stolte, S. The influence of salinity on the toxicity of selected sulfonamides and trimethoprim towards the green algae Chlorella vulgaris. J. Hazard. Mater. 2016, 308, 179–186. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, S.; Zhang, W.; Xiong, W.; Ye, Q.; Hou, X.; Wang, C.; Wang, P. Life cycle assessment of advanced wastewater treatment processes: Involving 126 pharmaceuticals and personal care products in life cycle inventory. J. Environ. Manag. 2019, 238, 442–450. [Google Scholar] [CrossRef]
- Patel, M.; Kumar, R.; Kishor, K.; Mlsna, T.; Pittman, C.U., Jr.; Mohan, D. Pharmaceuticals of emerging concern in aquatic systems: Chemistry, occurrence, effects, and removal methods. Chem. Rev. 2019, 119, 3510–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, Q.; Shi, X.; Yu, G.; Huang, J.; Wang, B. Assessing the persistence of pharmaceuticals in the aquatic environment: Challenges and needs. Emerg. Contam. 2016, 2, 145–147. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.Q.; Kim, S.J.; Kurade, M.B.; Govindwar, S.; Abou-Shanab, R.A.I.; Kim, J.R.; Roh, H.S.; Khan, M.A.; Jeon, B.H. Combined effects of sulfamethazine and sulfamethoxazole on a freshwater microalga, Scenedesmus obliquus: Toxicity, biodegradation, and metabolic fate. J. Hazard. Mater. 2019, 370, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Parus, A.; Karbowska, B. Marine algae as natural indicator of environmental cleanliness. Water Air Soil Pollut. 2020, 231. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.Q.; Kurade, M.B.; Abou-Shanab, R.A.; Ji, M.K.; Choi, J.; Kim, J.O.; Jeon, B.H. Biodegradation of carbamazepine using freshwater microalgae Chlamydomonas mexicana and Scenedesmus obliquus and the determination of its metabolic fate. Bioresour. Technol. 2016, 205, 183–190. [Google Scholar] [CrossRef]
- Bellingeri, A.; Bergami, E.; Grassi, G.; Faleri, C.; Redondo-Hasselerharm, P.; Koelmans, A.A.; Corsi, I. Combined effects of nanoplastics and copper on the freshwater alga Raphidocelis subcapitata. Aquat. Toxicol. 2019, 210, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Xu, W.; Na, J.; Lv, Z.; Zhang, Y. How long-term exposure of environmentally relevant antibiotics may stimulate the growth of Prorocentrum lima: A probable positive factor for red tides. Environ. Pollut. 2019, 255, 113149. [Google Scholar] [CrossRef]
- Baruah, P.; Chaurasia, N. Ecotoxicological effects of alpha-cypermethrin on freshwater alga Chlorella sp.: Growth inhibition and oxidative stress studies. Environ. Toxicol. Pharmacol. 2020, 76, 103347. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yan, X.; Shen, Y.; Di, M.; Wang, J. Antibiotics in surface water and sediments from Hanjiang River, Central China: Occurrence, behavior and risk assessment. Ecotoxicol. Environ. Saf. 2018, 157, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, H.; Du, J.; Qu, Y.; Shen, C.; Tan, F.; Chen, J.; Quan, X. Occurrence, removal, and risk assessment of antibiotics in 12 wastewater treatment plants from Dalian, China. Environ. Sci. Pollut. Res. Int. 2017, 24, 16478–16487. [Google Scholar] [CrossRef]
- Johnson, A.C.; Keller, V.; Dumont, E.; Sumpter, J.P. Assessing the concentrations and risks of toxicity from the antibiotics ciprofloxacin, sulfamethoxazole, trimethoprim and erythromycin in European rivers. Sci. Total Environ. 2015, 511, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Huang, X.; Witter, J.D.; Spongberg, A.L.; Wang, K.; Wang, D.; Liu, J. Occurrence of pharmaceuticals and personal care products and associated environmental risks in the central and lower Yangtze river, China. Ecotoxicol. Environ. Saf. 2014, 106, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.Q.; Kurade, M.B.; Patil, D.V.; Min, J.; Jeon, B.H. Biodegradation and metabolic fate of levofloxacin via a freshwater green alga, Scenedesmus obliquus in synthetic saline wastewater. Algal Res. 2017, 25, 54–61. [Google Scholar] [CrossRef]
- Rpra, B.; Dm, B.; Hn, C.; Ai, D. Resilience and self-regulation processes of microalgae under UV radiation stress. J. Photochem. Photobiol. C Photochem. Rev. 2020, 43, 100322. [Google Scholar] [CrossRef]
- Białk-Bielińska, A.; Stolte, S.; Arning, J.; Uebers, U.; Böschen, A.; Stepnowski, P.; Matzke, M. Ecotoxicity evaluation of selected sulfonamides. Chemosphere 2011, 85, 928–933. [Google Scholar] [CrossRef]
- Ferrari, B.; Mons, R.; Vollat, B.; Fraysse, B.; Paxēaus, N. Environmental risk assessment of six human pharmaceuticals: Are the current environmental risk assessment procedures sufficient for the protection of the aquatic environment? Environ. Toxicol. Chem. 2004, 23, 1344–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isidori, M.; Lavorgna, M.; Nardelli, A.; Pascarella, L.; Parrella, A. Toxic and genotoxic evaluation of six antibiotics on non-target organisms. Sci. Total Environ. 2005, 346, 87–98. [Google Scholar] [CrossRef]
- Meinertz, J.R.; Schreier, T.M.; Bernardy, J.A. Chronic toxicity of erythromycin thiocyanate to Daphnia magna in a flow-through, continuous exposure test system. Bull Environ. Contam. Toxicol. 2011, 87, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.D.; Soares, E.V. Impact of erythromycin on a non-target organism: Cellular effects on the freshwater microalga Pseudokirchneriella subcapitata. Aquat Toxicol. 2019, 208, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Nie, X.; Wang, X.; Chen, J.; Zitko, V.; An, T. Response of the freshwater alga Chlorella vulgaris to trichloroisocyanuric acid and ciprofloxacin. Environ. Toxicol. Chem. 2008, 27. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Le Manac’h, S.G.; Maccario, S.; Labrecque, M.; Lucotte, M.; Juneau, P. Differential effects of glyphosate and aminomethyl phosphonic acid (AMPA) on photosynthesis and chlorophyll metabolism in willow plants. Pestic. Biochem. Physiol. 2016, 130, 65–70. [Google Scholar] [CrossRef]
- Souza, L.R.R.; Bernardes, L.E.; Barbetta, M.F.S.; da Veiga, M. Iron oxide nanoparticle phytotoxicity to the aquatic plant Lemna minor: Effect on reactive oxygen species (ROS) production and chlorophyll a/chlorophyll b ratio. Environ. Sci. Pollut. Res. Int. 2019, 26, 24121–24131. [Google Scholar] [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Oxidative damage and antioxidative system in algae. Toxicol. Rep. 2019, 6, 1309–1313. [Google Scholar] [CrossRef]
- Biller, P.; Ross, A.B.; Skill, S.C.; Lea-Langton, A.; Balasundaram, B.; Hall, C.; Riley, R.; Llewellyn, C.A. Nutrient recycling of aqueous phase for microalgae cultivation from the hydrothermal liquefaction process. Algal Res. 2012, 1, 70–76. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, W.; Li, J.; Yuan, M.; Zhang, J.; Xu, F.; Xu, H.; Zheng, X.; Wang, L. Ecotoxicological effects of sulfonamides and fluoroquinolones and their removal by a green alga (Chlorella vulgaris) and a cyanobacterium (Chrysosporum ovalisporum). Environ. Pollut. 2020, 263, 114554. [Google Scholar] [CrossRef]
- Sathasivam, R.; Ki, J.S. Differential transcriptional responses of carotenoid biosynthesis genes in the marine green alga Tetraselmis suecica exposed to redox and non-redox active metals. Mol. Biol. Rep. 2019, 46, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Paliwal, C.; Pancha, I.; Ghosh, T.; Maurya, R.; Chokshi, K.; Bharadwaj, S.V.; Mishra, S. Selective carotenoid accumulation by varying nutrient media and salinity in Synechocystis sp CCNM 2501. Bioresour. Technol. 2015, 197, 363–368. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-mediated cellular signaling. Oxid. Med. Cell Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, H.; Li, J.; Pan, X.; Sun, Z.; Ye, C.; Jin, G.; Fu, Z. Effects of streptomycin on growth of algae Chlorella vulgaris and Microcystis aeruginosa. Environ. Toxicol. 2012, 27, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Shakunthala, N. New cytochrome P450 mechanisms: Implications for understanding molecular basis for drug toxicity at the level of the cytochrome. Expert Opin. Drug Met. Toxicol. 2009, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brezovsek, P.; Elersek, T.; Filipic, M. Toxicities of four anti-neoplastic drugs and their binary mixtures tested on the green alga Pseudokirchneriella subcapitata and the cyanobacterium Synechococcus leopoliensis. Water Res. 2014, 52, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Tormos, K.; Anso, E.; Hamanaka, R.; Eisenbart, J.; Joseph, J.; Kalyanaraman, B.; Chandel, N. Mitochondrial complex III ROS regulate adipocyte differentiation. Cell Metab. 2011, 14, 537–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateo, L.; Toffer, K.L.; Kawula, T.H. The sodA gene of Haemophilus ducreyi encodes a hydrogen peroxide-inhibitable superoxide dismutase. Gene 1998, 207, 251–257. [Google Scholar] [CrossRef]
- Hu, C.; Liu, X.; Li, X.; Zhao, J. Evaluation of growth and biochemical indicators of Salvinia natans exposed to zinc oxide nanoparticles and zinc accumulation in plants. Environ. Sci. Pollut. R. 2014, 21, 732–739. [Google Scholar] [CrossRef]
- Behne, D.; Kyriakopoulos, A. Mammalian selenium-containing proteins. Annu. Rev. Nutr. 2001. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lopez-Delgado, H.; Dat, J.F.; Scott, I.M. Hydrogen peroxide and glutathion-associated mechanisms of acclimatory stress tolerance and signaling. Physiol. Plant. 2010, 100, 241–254. [Google Scholar] [CrossRef]
- Alla, M.; Hassan, N.M. Changes of antioxidants levels in two maize lines following atrazine treatments. Plant. Physiol. Bioch. 2006, 44, 202–210. [Google Scholar] [CrossRef]
- Mehdy, M.C. Active oxygen species in plant defense against pathogens. Plant Physiol. 1994, 105, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Schafhauser, B.H.; Kristofco, L.A.; de Oliveira, C.M.R.; Brooks, B.W. Global review and analysis of erythromycin in the environment: Occurrence, bioaccumulation and antibiotic resistance hazards. Environ. Pollut. 2018, 238, 440–451. [Google Scholar] [CrossRef]
- Ogawa, K.; Nakatsugawa, T.; Yasuzaki, M. Heavy metacercarial infections of cyprinid fishes in Uji River. Fish. Ence 2010, 70, 132–140. [Google Scholar] [CrossRef]
- Lin, J.H.; Kao, W.C.; Tsai, K.P.; Chen, C.Y. A novel algal toxicity testing technique for assessing the toxicity of both metallic and organic toxicants. Water Res. 2005, 39, 1869–1877. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.W.; Jung, C.; Li, H.; Yu, M.; Flora, J.R.; Boateng, L.K.; Her, N.; Zoh, K.D.; Yoon, Y. Adsorption characteristics of diclofenac and sulfamethoxazole to graphene oxide in aqueous solution. Chemosphere 2015, 136, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Lekkerkerker-Teunissen, K.; Benotti, M.J.; Snyder, S.A.; van Dijk, H.C. Transformation of atrazine, carbamazepine, diclofenac and sulfamethoxazole by low and medium pressure UV and UV/H2O2 treatment. Sep. Purif. Technol. 2012, 96, 33–43. [Google Scholar] [CrossRef]
- Vasickova, J.; Hvezdova, M.; Kosubova, P.; Hofman, J. Ecological risk assessment of pesticide residues in arable soils of the Czech Republic. Chemosphere 2019, 216, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Pleiter, M.; Gonzalo, S.; Rodea-Palomares, I.; Leganes, F.; Rosal, R.; Boltes, K.; Marco, E.; Fernandez-Pinas, F. Toxicity of five antibiotics and their mixtures towards photosynthetic aquatic organisms: Implications for environmental risk assessment. Water Res. 2013, 47, 2050–2064. [Google Scholar] [CrossRef]
- Afkari, A. An investigation to the vermicompost efficacy on the activity level of antioxidant enzymes and photosynthetic pigments of borage under Salinity Stress Conditions. Russ. Agric. Sci. 2018, 44, 310–317. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (Days) | 2 | 4 | 7 | 11 |
|---|---|---|---|---|
| EC50 of SMX (mg/L) | 0.76 | 0.49 | 1.27 | 1.55 |
| RQ of SMX surface water | 5.35 a | 8.31 a | 3.20 a | 2.63 a |
| RQ of SMX wastewater | 15.66 a | 23.22 a | 9.37 a | 7.68 a |
| EC50 of ERY (mg/L) | 0.069 | 0.044 | 0.046 | 0.082 |
| RQ of ERY surface water | 5.07 b | 7.95 b | 7.61 b | 4.27 b |
| RQ of ERY wastewater | 12.02 b | 18.86 b | 18.24 b | 10.12 b |
| EC50 of mixture (mg/L) | 0.16 | 0.06 | 0.08 | 0.07 |
| RQ of mixture surface water | 27.63 c | 73.67 c | 55.25 c | 63.14 c |
| RQ of mixture wastewater | 79.44 c | 211.83 c | 158.88 c | 181.57 c |
| Algae species | EC50 of SMX (mg/L) | Major Focus of the Study | References |
|---|---|---|---|
| Chlorella vulgaris | 48 h EC50 = 1.51 96 h EC50 = 0.98 | Ecotoxicological evaluation | [17] |
| Synechococcus leopolensis Raphidocelis subcapitata Raphidocelis subcapitata | 96 h EC50 = 0.0268 96 h EC50 = 0.146 72 h EC50 = 0.52 | Ecotoxicological evaluation Ecotoxicological evaluation Ecotoxicological evaluation | [18,19] |
| Scenedesmus obliquus | 96 h EC50 = 0.018 | Ecotoxicology evaluation, modeling of toxicity, risk assessment | [5] |
| Algae species Raphidocelis subcapitata Dunaliella tertiolecta Raphidocelis subcapitata | EC50 of ERY (mg/L) 72 h EC50 = 0.02 96 h EC50 = 0.0272 h EC50 = 0.38 | Ecotoxicological evaluation Ecotoxicological evaluation | [20,21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; He, D.; Chang, F.; Dang, C.; Fu, J. Combined Effects of Sulfamethoxazole and Erythromycin on a Freshwater Microalga, Raphidocelis subcapitata: Toxicity and Oxidative Stress. Antibiotics 2021, 10, 576. https://doi.org/10.3390/antibiotics10050576
Zhang Y, He D, Chang F, Dang C, Fu J. Combined Effects of Sulfamethoxazole and Erythromycin on a Freshwater Microalga, Raphidocelis subcapitata: Toxicity and Oxidative Stress. Antibiotics. 2021; 10(5):576. https://doi.org/10.3390/antibiotics10050576
Chicago/Turabian StyleZhang, Yibo, Da He, Fang Chang, Chenyuan Dang, and Jie Fu. 2021. "Combined Effects of Sulfamethoxazole and Erythromycin on a Freshwater Microalga, Raphidocelis subcapitata: Toxicity and Oxidative Stress" Antibiotics 10, no. 5: 576. https://doi.org/10.3390/antibiotics10050576
APA StyleZhang, Y., He, D., Chang, F., Dang, C., & Fu, J. (2021). Combined Effects of Sulfamethoxazole and Erythromycin on a Freshwater Microalga, Raphidocelis subcapitata: Toxicity and Oxidative Stress. Antibiotics, 10(5), 576. https://doi.org/10.3390/antibiotics10050576