
Xanthones Active against Multidrug Resistance and Virulence Mechanisms of Bacteria

 ,
,  , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
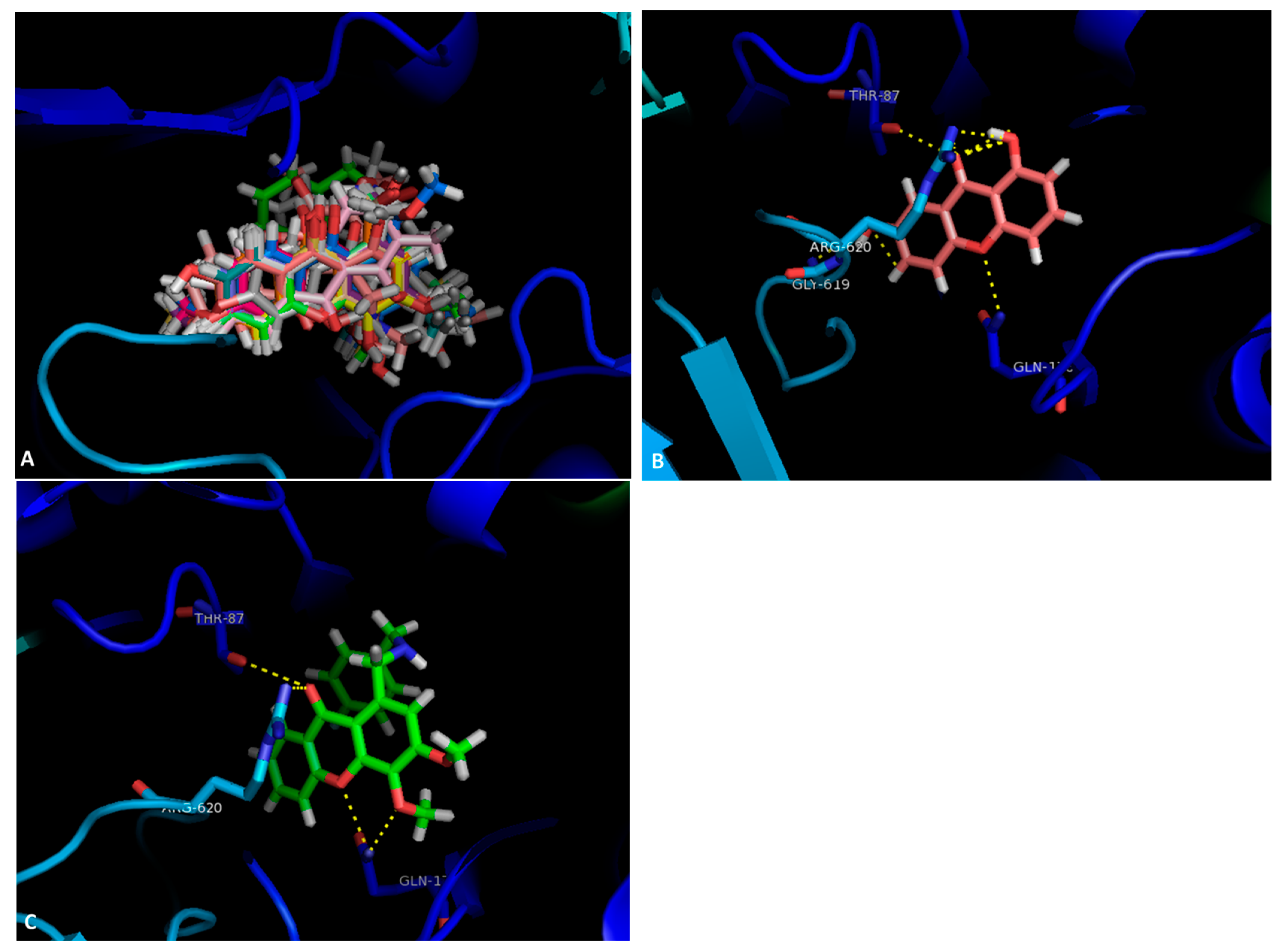
2.1. Docking Results
2.2. Antibacterial Activity
2.3. Efflux Pump Inhibition Assay
2.4. Inhibition of Biofilm Formation
2.5. Quorum Sensing Assay
2.6. Cytotoxicity Assay
3. Materials and Methods
3.1. Compounds
3.2. Culture Media and Chemicals
3.3. Docking Studies
3.4. Bacterial Strains
3.5. Antibacterial Assay
3.6. Efflux Pump Inhibition Assay
3.7. Inhibition of Biofilm Formation
3.8. Quorum Sensing Assay
3.9. Cytotoxicity Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AHL | Acyl-homoserine-lactone |
| ATCC | American Type Culture Collection |
| BCR | Binding core region |
| CCCP | Carbonyl cyanide 3-chlorophenylhydrazone |
| CS | Cytoplasmic side |
| CV026 | Chromobacterium violaceum CV026 |
| CV | Crystal violet |
| DMSO | Dimethyl sulfoxide |
| DTSSP | 3,3′-dithiobis(sulfosuccinimidyl propionate) |
| EB | Ethidium bromide |
| EZF | Sphingomonas paucimobilis EZF 10-17 |
| HH | Helical hairpin |
| HT | Hydrophobic trap |
| IC50 | Half-maximal inhibitory concentration |
| LB-A | Luria–Bertani agar |
| LB-B | Luria–Bertani broth |
| LD | Lipoyl domain |
| MFS | Major facilitator superfamily |
| MHB II | Cation-adjusted Mueller–Hinton broth |
| MIC | Minimum inhibitory concentration |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| OD | Optical density |
| PBS | Phosphate-buffered saline |
| PMZ | Prometazine |
| RF | Relative fluorescence |
| RFI | Relative fluorescence index |
| RND | Resistance-nodulation-division |
| SBS | Substrate-binding site |
| SD | Standard deviation |
| SE03 | Salmonella enterica serovar Typhimurium with the acrA gene deleted |
| TSA | Tryptic Soy agar |
| TSB | Tryptic Soy broth |
| QS | Quorum sensing |
| wt85 | Chromobacterium violaceum wild type 85 |
References
- Cheng, G.; Dai, M.; Ahmed, S.; Hao, H.; Wang, X.; Yuan, Z. Antimicrobial Drugs in Fighting against Antimicrobial Resistance. Front. Microbiol. 2016, 7, 470. [Google Scholar] [CrossRef] [Green Version]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Genet. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.A.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial Multidrug Efflux Pumps: Much More Than Antibiotic Resistance Determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Durães, F.; Pinto, M.; Sousa, E. Medicinal Chemistry Updates on Bacterial Efflux Pump Modulators. Curr. Med. Chem. 2019, 25, 6030–6069. [Google Scholar] [CrossRef]
- Pagès, J.-M.; Amaral, L. Mechanisms of drug efflux and strategies to combat them: Challenging the efflux pump of Gram-negative bacteria. Biochim. Biophys. Acta (BBA)–Proteins Proteom. 2009, 1794, 826–833. [Google Scholar] [CrossRef]
- Sharma, N.; Chhillar, A.K.; Dahiya, S.; Choudhary, P.; Punia, A.; Gulia, P. Antibiotic adjuvants: A promising approach to combat multidrug resistant bacteria. Curr. Drug Targets 2021, 22, 1. [Google Scholar] [CrossRef]
- Oliveira-Tintino, C.D.D.M.; Muniz, D.F.; Barbosa, C.R.D.S.; Pereira, R.L.S.; Begnini, I.M.; Rebelo, R.A.; da Silva, L.E.; Mireski, S.L.; Nasato, M.C.; Krautler, M.I.L.; et al. The 1,8-naphthyridines sulfonamides are NorA efflux pump inhibitors. J. Glob. Antimicrob. Resist. 2021, 24, 233–240. [Google Scholar] [CrossRef]
- Puzari, M.; Chetia, P. Virtual high-throughput screening and simulation studies of compounds from selected traditionally important medicinal plants for the identification of potential inhibitors of AcrB. J. Biomol. Struct. Dyn. 2020, 1–9. [Google Scholar] [CrossRef]
- de Araújo, A.C.; Freitas, P.R.; Barbosa, C.R.d.S.; Muniz, D.F.; Ribeiro-Filho, J.; Tintino, S.R.; Júnior, J.P.; Filho, J.M.; de Sousa, G.R.; Coutinho, H.D.; et al. Modulation of Drug Resistance by Limonene: Inhibition of Efflux Pumps in Staphylococcus aureus Strains RN-4220 and IS-58. Curr. Drug Metab. 2021, 22, 110–113. [Google Scholar] [CrossRef]
- Loureiro, D.R.P.; Soares, J.X.; Costa, J.C.; Magalhães, Á.F.; Azevedo, C.M.G.; Pinto, M.M.M.; Afonso, C.M.M. Structures, Activities and Drug-Likeness of Anti-Infective Xanthone Derivatives Isolated from the Marine Environment: A Review. Molecules 2019, 24, 243. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.; Palmeira, A.; Fernandes, C.; Resende, D.; Sousa, E.; Cidade, H.; Tiritan, M.; Correia-Da-Silva, M.; Cravo, S. From Natural Products to New Synthetic Small Molecules: A Journey through the World of Xanthones. Molecules 2021, 26, 431. [Google Scholar] [CrossRef]
- Kuete, V.; Alibert-Franco, S.; Eyong, K.; Ngameni, B.; Folefoc, G.; Nguemeving, J.; Tangmouo, J.; Fotso, G.; Komguem, J.; Ouahouo, B.; et al. Antibacterial activity of some natural products against bacteria expressing a multidrug-resistant phenotype. Int. J. Antimicrob. Agents 2011, 37, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Resende, D.I.S.P.; Pereira-Terra, P.; Inácio, Â.S.; Da Costa, P.M.; Pinto, E.; Sousa, E.; Pinto, M.M.M. Lichen Xanthones as Models for New Antifungal Agents. Molecules 2018, 23, 2617. [Google Scholar] [CrossRef] [Green Version]
- Resende, D.I.S.P.; Pereira-Terra, P.; Moreira, J.; Freitas-Silva, J.; Lemos, A.; Gales, L.; Pinto, E.; De Sousa, M.E.; Da Costa, P.M.; Pinto, M.M.M. Synthesis of a Small Library of Nature-Inspired Xanthones and Study of Their Antimicrobial Activity. Molecules 2020, 25, 2405. [Google Scholar] [CrossRef]
- Bessa, L.J.; Palmeira, A.; Gomes, A.S.; Vasconcelos, V.; Sousa, E.; Pinto, M.; Da Costa, P.M. Synergistic Effects Between Thioxanthones and Oxacillin Against Methicillin-Resistant Staphylococcus aureus. Microb. Drug Resist. 2015, 21, 404–415. [Google Scholar] [CrossRef]
- Nové, M.; Kincses, A.; Szalontai, B.; Rácz, B.; Blair, J.M.A.; González-Prádena, A.; Benito-Lama, M.; Domínguez-Álvarez, E.; Spengler, G. Biofilm Eradication by Symmetrical Selenoesters for Food-Borne Pathogens. Microorganisms 2020, 8, 566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvist, M.; Hancock, V.; Klemm, P. Inactivation of Efflux Pumps Abolishes Bacterial Biofilm Formation. Appl. Environ. Microbiol. 2008, 74, 7376–7382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, S.M. Role of efflux pumps in the antibiotic resistance of bacteria embedded in a biofilm. Virulence 2013, 4, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahmy, A.; Srinivasan, A.; Webber, M.A. The Relationship between Bacterial Multidrug Efflux Pumps and Biofilm Formation. In Efflux-Mediated Antimicrobial Resistance in Bacteria; Metzler, J.B., Ed.; Oxford University Press: Oxford, UK, 2016; pp. 651–663. [Google Scholar]
- Sabatini, S.; Piccioni, M.; Felicetti, T.; De Marco, S.; Manfroni, G.; Pagiotti, R.; Nocchetti, M.; Cecchetti, V.; Pietrella, D. Investigation on the effect of known potent S. aureus NorA efflux pump inhibitors on the staphylococcal biofilm formation. RSC Adv. 2017, 7, 37007–37014. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.; Zheng, C.; Liu, Y.; Qu, L.; Yu, H.; Han, L.; Zhang, Y.; Wang, T. Chemical and Biological Research on Herbal Medicines Rich in Xanthones. Molecules 2017, 22, 1698. [Google Scholar] [CrossRef] [Green Version]
- Mardanova, A.M.; Bogomol’Naya, L.M.; Romanova, Y.D.; Sharipova, M.R. Efflux systems in Serratia marcescens. Microbiology 2013, 82, 668–679. [Google Scholar] [CrossRef]
- Benomar, S.; Evans, K.C.; Unckless, R.L.; Chandler, J.R. Efflux Pumps in Chromobacterium Species Increase Antibiotic Resistance and Promote Survival in a Coculture Competition Model. Appl. Environ. Microbiol. 2019, 85, e00908-19. [Google Scholar] [CrossRef] [Green Version]
- Mouwakeh, A.; Kincses, A.; Nové, M.; Mosolygó, T.; Mohácsi-Farkas, C.; Kiskó, G.; Spengler, G. Nigella sativa essential oil and its bioactive compounds as resistance modifiers against Staphylococcus aureus. Phytother. Res. 2019, 33, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Resende, D.I.S.P.; Durães, F.; Maia, M.; Sousa, E.; Pinto, M.M.M. Recent advances in the synthesis of xanthones and azaxanthones. Org. Chem. Front. 2020, 7, 3027–3066. [Google Scholar] [CrossRef]
- Shi, X.; Chen, M.; Yu, Z.; Bell, J.M.; Wang, H.; Forrester, I.; Villarreal, H.; Jakana, J.; Du, D.; Luisi, B.F.; et al. In situ structure and assembly of the multidrug efflux pump AcrAB-TolC. Nat. Commun. 2019, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zárate, S.G.; Morales, P.; Świderek, K.; Bolanos-Garcia, V.M.; Bastida, A. A Molecular Modeling Approach to Identify Novel Inhibitors of the Major Facilitator Superfamily of Efflux Pump Transporters. Antibiotics 2019, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Shaheen, A.; Afridi, W.A.; Mahboob, S.; Sana, M.; Zeeshan, N.; Ismat, F.; Mirza, O.; Iqbal, M.; Rahman, M. Reserpine Is the New Addition into the Repertoire of AcrB Efflux Pump Inhibitors. Mol. Biol. Mosk 2019, 53, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Kincses, A.; Varga, B.; Csonka, Á.; Sancha, S.; Mulhovo, S.; Madureira, A.M.; Ferreira, M.-J.U.; Spengler, G. Bioactive compounds from the African medicinal plant Cleistochlamys kirkii as resistance modifiers in bacteria. Phytother. Res. 2018, 32, 1039–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, S.S.; Sobkowiak, B.; Parreira, R.; Edgeworth, J.D.; Viveiros, M.; Clark, T.G.; Couto, I. Genetic Diversity of norA, Coding for a Main Efflux Pump of Staphylococcus aureus. Front. Genet. 2019, 9, 710. [Google Scholar] [CrossRef]
- Nagem, T.J.; Oliveira, F.F.d. Xanthones and other constituents of Vismia parviflora. J. Braz. Chem. Soc. 1997, 8, 505–508. [Google Scholar] [CrossRef] [Green Version]
- Kato, L.; De Oliveira, C.M.A.; Vencato, I.; Lauriucci, C. Crystal structure of 1,7-dihydroxyxanthone from Weddellina squamulosa Tul. J. Chem. Crystallogr. 2005, 35, 23–26. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, srep42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, V.; Gil-Martins, E.; Rocha-Pereira, C.; Lemos, A.; Palmeira, A.; Puthongking, P.; Sousa, E.; Bastos, M.D.L.; Remião, F.; Silva, R. Oxygenated xanthones as P-glycoprotein modulators at the intestinal barrier: In vitro and docking studies. Med. Chem. Res. 2020, 29, 1041–1057. [Google Scholar] [CrossRef]
- Gibbons, S.; Oluwatuyi, M.; Kaatz, G.W. A novel inhibitor of multidrug efflux pumps in Staphylococcus aureus. J. Antimicrob. Chemother. 2003, 51, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Kaatz, G.W.; Moudgal, V.V.; Seo, S.M.; Kristiansen, J.E. Phenothiazines and Thioxanthenes Inhibit Multidrug Efflux Pump Activity in Staphylococcus aureus. Antimicrob. Agents Chemother. 2003, 47, 719–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the Mechanism of Bacterial Biofilms Resistance to Antimicrobial Agents. Open Microbiol. J. 2017, 11, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [Green Version]
- Parai, D.; Banerjee, M.; Dey, P.; Mukherjee, S.K. Reserpine attenuates biofilm formation and virulence of Staphylococcus aureus. Microb. Pathog. 2020, 138, 103790. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Cui, P.; Zhang, Y.; Zhang, W. Identification of Novel Efflux Proteins Rv0191, Rv3756c, Rv3008, and Rv1667c Involved in Pyrazinamide Resistance in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Tariq, A.; Sana, M.; Shaheen, A.; Ismat, F.; Mahboob, S.; Rauf, W.; Mirza, O.; Iqbal, M.; Rahman, M. Restraining the multidrug efflux transporter STY4874 of Salmonella Typhi by reserpine and plant extracts. Lett. Appl. Microbiol. 2019, 69, 161–167. [Google Scholar] [CrossRef]
- Romanova, N.A.; Wolffs, P.F.G.; Brovko, L.Y.; Griffiths, M.W. Role of Efflux Pumps in Adaptation and Resistance of Listeria monocytogenes to Benzalkonium Chloride. Appl. Environ. Microbiol. 2006, 72, 3498–3503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathania, R.; Sharma, A.; Gupta, V.K. Efflux pump inhibitors for bacterial pathogens: From bench to bedside. Indian J. Med. Res. 2019, 149, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Hagmann, W.K. The Many Roles for Fluorine in Medicinal Chemistry. J. Med. Chem. 2008, 51, 4359–4369. [Google Scholar] [CrossRef]
- Greenlee, W.F.; Osborne, R.; Dold, K.M.; Hudson, L.G.; Toscano, W.A. Toxicity of chlorinated aromatic compounds in animals and humans: In vitro approaches to toxic mechanisms and risk assessment. Environ. Heal. Perspect. 1985, 60, 69–76. [Google Scholar] [CrossRef]
- Pedro, M.; Cerqueira, F.; Sousa, M.E.; Nascimento, M.S.J.; Pinto, M. Xanthones as inhibitors of growth of human cancer cell lines and Their effects on the proliferation of human lymphocytes In Vitro. Bioorg. Med. Chem. 2002, 10, 3725–3730. [Google Scholar] [CrossRef]
- Pinto, E.; Afonso, C.; Duarte, S.; Vale-Silva, L.; Costa, E.; Sousa, E.; Pinto, M. Antifungal Activity of Xanthones: Evaluation of their Effect on Ergosterol Biosynthesis by High-performance Liquid Chromatography. Chem. Biol. Drug Des. 2011, 77, 212–222. [Google Scholar] [CrossRef]
- Eicher, T.; Cha, H.-J.; Seeger, M.A.; Brandstatter, L.; El-Delik, J.; Bohnert, J.A.; Kern, W.V.; Verrey, F.; Grutter, M.G.; Diederichs, K.; et al. Transport of drugs by the multidrug transporter AcrB involves an access and a deep binding pocket that are separated by a switch-loop. Proc. Natl. Acad. Sci. USA 2012, 109, 5687–5692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikolosko, J.; Bobyk, K.; Zgurskaya, H.I.; Ghosh, P. Conformational Flexibility in the Multidrug Efflux System Protein AcrA. Structure 2006, 14, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Koronakis, V.; Sharff, A.; Koronakis, E.; Luisi, B.F.; Hughes, C. Crystal structure of the bacterial membrane protein TolC central to multidrug efflux and protein export. Nat. Cell Biol. 2000, 405, 914–919. [Google Scholar] [CrossRef]
- Sussman, J.L.; Lin, D.; Jiang, J.; Manning, N.O.; Prilusky, J.; Ritter, O.; Abola, E.E. Protein Data Bank (PDB): Database of Three-Dimensional Structural Information of Biological Macromolecules. Acta Crystallogr. Sect. D Biol. Crystallogr. 1998, 54, 1078–1084. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aron, Z.; Opperman, T.J. The hydrophobic trap—the Achilles heel of RND efflux pumps. Res. Microbiol. 2018, 169, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; Beer, T.A.P.D.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The UniProt Consortium UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [CrossRef] [Green Version]
- Seeliger, D.; De Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Gajdács, M.; Spengler, G. Standard operating procedure (SOP) for disk diffusion-based quorum sensing inhibition assays. Acta Pharm. Hung. 2020, 89, 117–125. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Gajdács, M.; Spengler, G. The Role of Drug Repurposing in the Development of Novel Antimicrobial Drugs: Non-Antibiotic Pharmacological Agents as Quorum Sensing-Inhibitors. Antibiotics 2019, 8, 270. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, R.J.; Kincses, A.; Gajdács, M.; Spengler, G.; Dos Santos, D.J.V.A.; Molnár, J.; Ferreira, M.-J.U. Terpenoids from Euphorbia pedroi as Multidrug-Resistance Reversers. J. Nat. Prod. 2018, 81, 2032–2040. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound | Docking Score | ||||||||||||
| AcrB | AcrA | TolC | NorA | ||||||||||
| No. | R1 | R2 | R3 | R4 | R5 | R6 | SBS | HT | HH | LD | BCR | CS | |
| 1 | H | H | H | OH | H | H | −6.7 | −5.9 | −6.2 | −6.2 | −6.4 | −6.6 | −6.3 |
| 2 | H | OH | OH | H | H | H | −6.9 | −6.2 | −6.2 | −6.7 | −6.5 | −6.2 | −6.1 |
| 3 | OH | OH | H | H | H | H | −6.6 | −5.5 | −6.4 | −6.8 | −7.0 | −6.5 | −6.4 |
| 4 | OH | H | H | H | H | OH | −6.9 | −5.9 | −6.4 | −6.7 | −6.5 | −6.6 | −6.3 |
| 5 | OH | CH3 | OH | H | H | H | −6.9 | −6.2 | −6.4 | −6.3 | −6.6 | −6.2 | −5.5 |
| 6 | CH3 | H | OH | OH | H | H | −6.9 | −6.1 | −6.7 | −7.0 | −6.7 | −6.9 | −6.6 |
| 7 | CH3 | H | OH | OH | OH | H | −7.2 | −6.4 | −6.0 | −7.8 | −6.6 | −5.9 | −5.9 |
| 8 | CH3 | H | OCH3 | OCH3 | H | H | −6.8 | −5.9 | −5.7 | −5.9 | −6.6 | −5.1 | −5.4 |
| 9 | CH3 | Cl | OCH3 | OCH3 | H | H | −7.0 | −5.1 | −5.8 | −5.9 | −6.4 | −4.8 | −5.2 |
| 10 | CH3 | H | OCH3 | OCH3 | OCH3 | H | −7.1 | −5.5 | −5.3 | −6.1 | −6.6 | −4.8 | −5.2 |
| 11 | OCH3 | H | H | H | H | H | −6.8 | −5.1 | −6.4 | −6.0 | −6.7 | −6.8 | −6.8 |
| 12 | CH2Br | H | OCH3 | OCH3 | H | H | −7.1 | −5.0 | −5.3 | −5.5 | −6.2 | −3.4 | −5.7 |
| 13 | CH2Br | H | OCH3 | OCH3 | OCH3 | H | −7.0 | −5.0 | −5.1 | −5.7 | −6.1 | −2.9 | −5.6 |
| 14 | CHO | H | OCH3 | OH | H | H | −7.2 | −6.2 | −5.7 | −6.7 | −6.9 | −6.3 | −5.3 |
| 15 | CHO | H | OCH3 | OCH3 | H | H | −7.2 | −5.0 | −5.5 | −6.1 | −6.2 | −4.7 | −5.2 |
| 16 | CHO | H | OCH3 | OCH3 | OCH3 | H | −7.3 | −5.9 | −5.3 | −6.4 | −6.3 | −4.7 | −5.1 |
| 17 | CO2CH3 | H | OCH3 | OCH3 | OCH3 | H | −7.5 | −5.9 | −5.5 | −4.9 | −6.5 | −4.5 | −5.2 |
| 18 |  | H | OCH3 | OCH3 | H | H | −8.0 | 8.6 | −7.0 | −6.2 | −7.9 | −7.9 | −5.4 |
| 19 |  | H | OCH3 | OCH3 | H | H | −7.3 | −5.8 | −6.3 | −6.3 | −6.9 | −7.9 | −6.0 |
| 20 |  | H | OCH3 | OCH3 | H | H | −7.6 | −5.5 | −7.0 | −6.4 | −7.9 | −5.6 | −6.4 |
| D13-9001 | −9.7 | 26.5 | −6.2 | −5.1 | −7.4 | 0.7 | −4.7 | ||||||
| Doxorubicin | −8.9 | 15.4 | −7.2 | −5.6 | −7.2 | −0.5 | −4.7 | ||||||
| MBX-3132 | −7.9 | 2.9 | −7.9 | −6.2 | −7.7 | −6.2 | −7.0 | ||||||
| Minocycline | −8.7 | 26.7 | −6.2 | −5.4 | −7.7 | 1.3 | −4.9 | ||||||
| PAβN | −7.1 | −4.7 | −5.8 | −4.9 | −7.1 | −9.4 | −5.3 | ||||||
| Reserpine | −8.7 | 10.9 | 5.6 | 4.6 | −7.5 | 1.0 | −4.6 | ||||||
| RFI ± SD | ||
|---|---|---|
| Compound | S. aureus 272123 | SE03 |
| 1 | −0.36 ± 0.03 1 | −0.01 ± 0.05 6 |
| 2 | −0.69 ± 0.02 2 | −0.57 ± 0.02 7 |
| 3 | −0.05 ± 0.06 3 | 0.23 ± 0.04 8 |
| 4 | 0.13 ± 0.08 2 | 2.90 ± 0.71 7 |
| 5 | 0.16 ± 0.08 2 | 1.74 ± 0.22 7 |
| 6 | −0.91 ± 0.01 4 | −0.91 ± 0.01 9 |
| 7 | −0.92 ± 0.00 5 | −0.87 ± 0.02 10 |
| 8 | 0.15 ± 0.25 4 | 0.05 ± 0.11 9 |
| 9 | −0.04 ± 0.26 4 | 0.05 ± 0.09 9 |
| 10 | 0.18 ± 0.20 5 | 0.24 ± 0.05 10 |
| 11 | −0.10 ± 0.06 1 | 0.28 ± 0.09 6 |
| 12 | 0.11 ± 0.30 4 | 1.75 ± 1.57 9 |
| 13 | 0.53 ± 0.20 5 | 1.16 ± 0.64 10 |
| 14 | −0.28 ± 0.06 2 | 0.02 ± 0.02 7 |
| 15 | 0.08 ± 0.07 4 | 0.13 ± 0.09 9 |
| 16 | 5.49 ± 8.04 5 | 0.13 ± 0.15 10 |
| 17 | −0.34 ± 0.02 5 | −0.29 ± 0.08 10 |
| 18 | −0.01 ± 0.04 3 | 0.04 ± 0.04 8 |
| 19 | 0.02 ± 0.02 3 | 2.86 ± 0.14 8 |
| 20 | 1.08 ± 0.82 4 | 2.09 ± 0.05 11 |
| Reserpine | 1 0.31 ± 0.07 2 0.45 ± 0.04 3 0.84 ± 0.13 4 0.35 ± 0.14 5 0.16 ± 0.05 | --- |
| CCCP | --- | 6 0.23 ± 0.04 7 0.37 ± 0.04 8 0.33 ± 0.09 9 0.40 ± 0.03 10 0.27 ± 0.14 11 0.50 ± 0.11 |
| Inhibition of Biofilm Formation (%) ± SD | ||
|---|---|---|
| Compound | S. aureus ATCC 29213 | S. aureus 272123 |
| 4 | 0 1 | 94.21 ± 1.31 3 |
| 5 | 0 1 | 61.62 ± 16.51 3 |
| 11 | 0 1 | 39.95 ± 7.66 3 |
| 12 | 0 1 | 6.27 ± 1.41 3 |
| 16 | 0 2 | 58.98 ± 12.00 4 |
| 19 | 97.45 ± 0.62 2 | 65.03 ± 9.86 4 |
| 20 | 90.76 ± 1.62 2 | 77.17 ± 4.45 4 |
| Reserpine | 1 2.49 ± 1.99 2 22.29 ± 10.88 | 3 77.62 ± 10.44 4 63.42 ± 2.63 |
| Quorum Sensing Inhibition (mm) ± SD | |||
|---|---|---|---|
| Compound | S. marcescens | EZF + CV026 | wt85 |
| 4 | 0 | 0 | 0 |
| 5 | 0 | 0 | 0 |
| 11 | 0 | 0 | 0 |
| 12 | 0 | 30 ± 0.5 | 0 |
| 16 | 0 | 42 ± 0.8 | 0 |
| 19 | 0 | 0 | 0 |
| 20 | 0 | 0 | 0 |
| PMZ | 18 ± 0.8 | 40 ± 0.1 | 41 ± 0.5 |
| Compound | IC50 (µM) ± SD |
|---|---|
| 4 | >100 |
| 12 | 54.59 ± 5.30 |
| 16 | 26.93 ± 5.87 |
| 19 | >100 |
| 20 | 35.12 ± 4.86 |
| Doxorubicin | 12.05 ± 0.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durães, F.; Resende, D.I.S.P.; Palmeira, A.; Szemerédi, N.; Pinto, M.M.M.; Spengler, G.; Sousa, E. Xanthones Active against Multidrug Resistance and Virulence Mechanisms of Bacteria. Antibiotics 2021, 10, 600. https://doi.org/10.3390/antibiotics10050600
Durães F, Resende DISP, Palmeira A, Szemerédi N, Pinto MMM, Spengler G, Sousa E. Xanthones Active against Multidrug Resistance and Virulence Mechanisms of Bacteria. Antibiotics. 2021; 10(5):600. https://doi.org/10.3390/antibiotics10050600
Chicago/Turabian StyleDurães, Fernando, Diana I. S. P. Resende, Andreia Palmeira, Nikoletta Szemerédi, Madalena M. M. Pinto, Gabriella Spengler, and Emília Sousa. 2021. "Xanthones Active against Multidrug Resistance and Virulence Mechanisms of Bacteria" Antibiotics 10, no. 5: 600. https://doi.org/10.3390/antibiotics10050600
APA StyleDurães, F., Resende, D. I. S. P., Palmeira, A., Szemerédi, N., Pinto, M. M. M., Spengler, G., & Sousa, E. (2021). Xanthones Active against Multidrug Resistance and Virulence Mechanisms of Bacteria. Antibiotics, 10(5), 600. https://doi.org/10.3390/antibiotics10050600