
Super-Cationic Peptide Dendrimers—Synthesis and Evaluation as Antimicrobial Agents

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
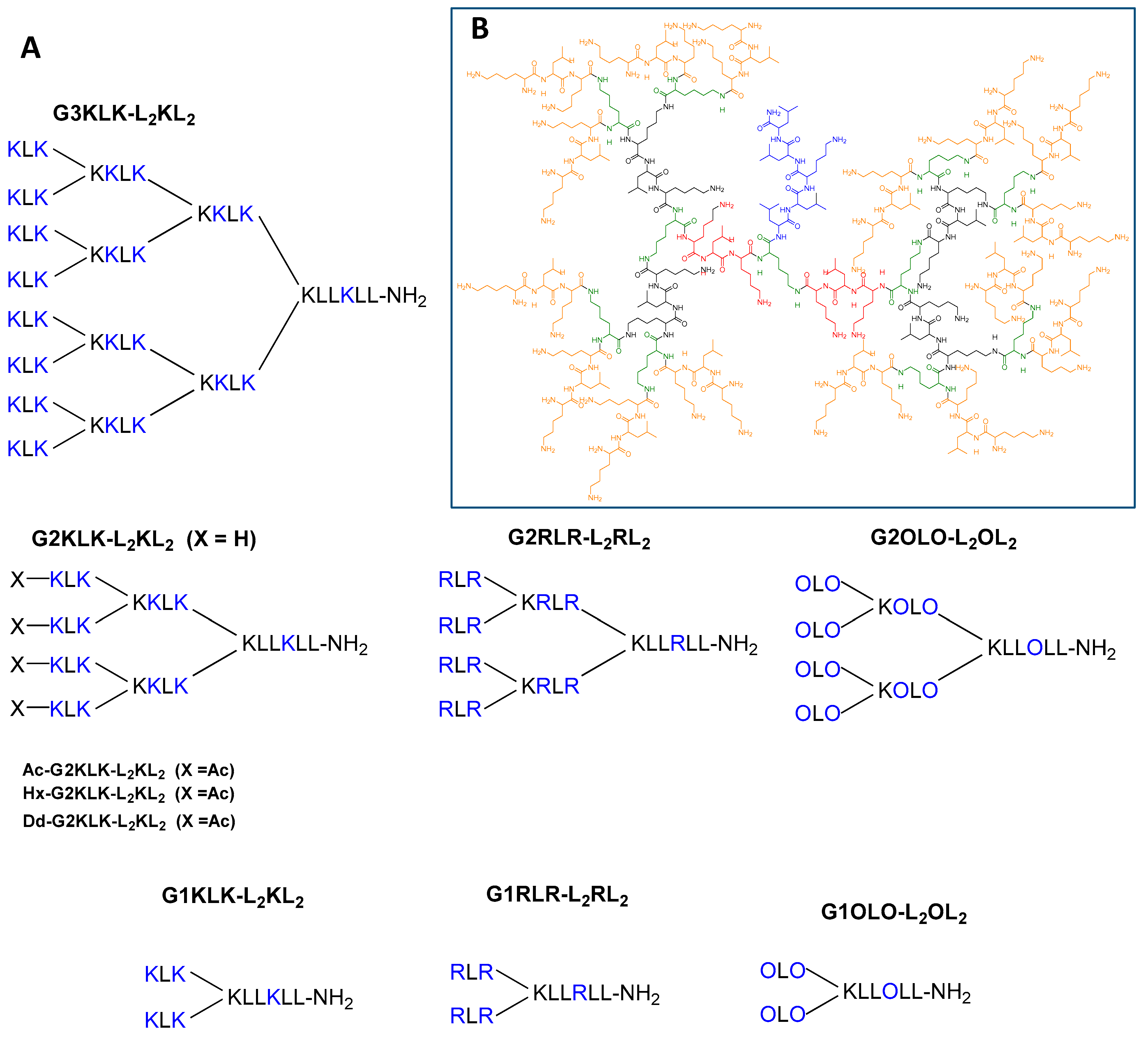
2.1. Design and Synthesis of Super-Cationic Peptide Dendrimers (SCPDs)
2.2. Antimicrobial Action (MIC and MBC)
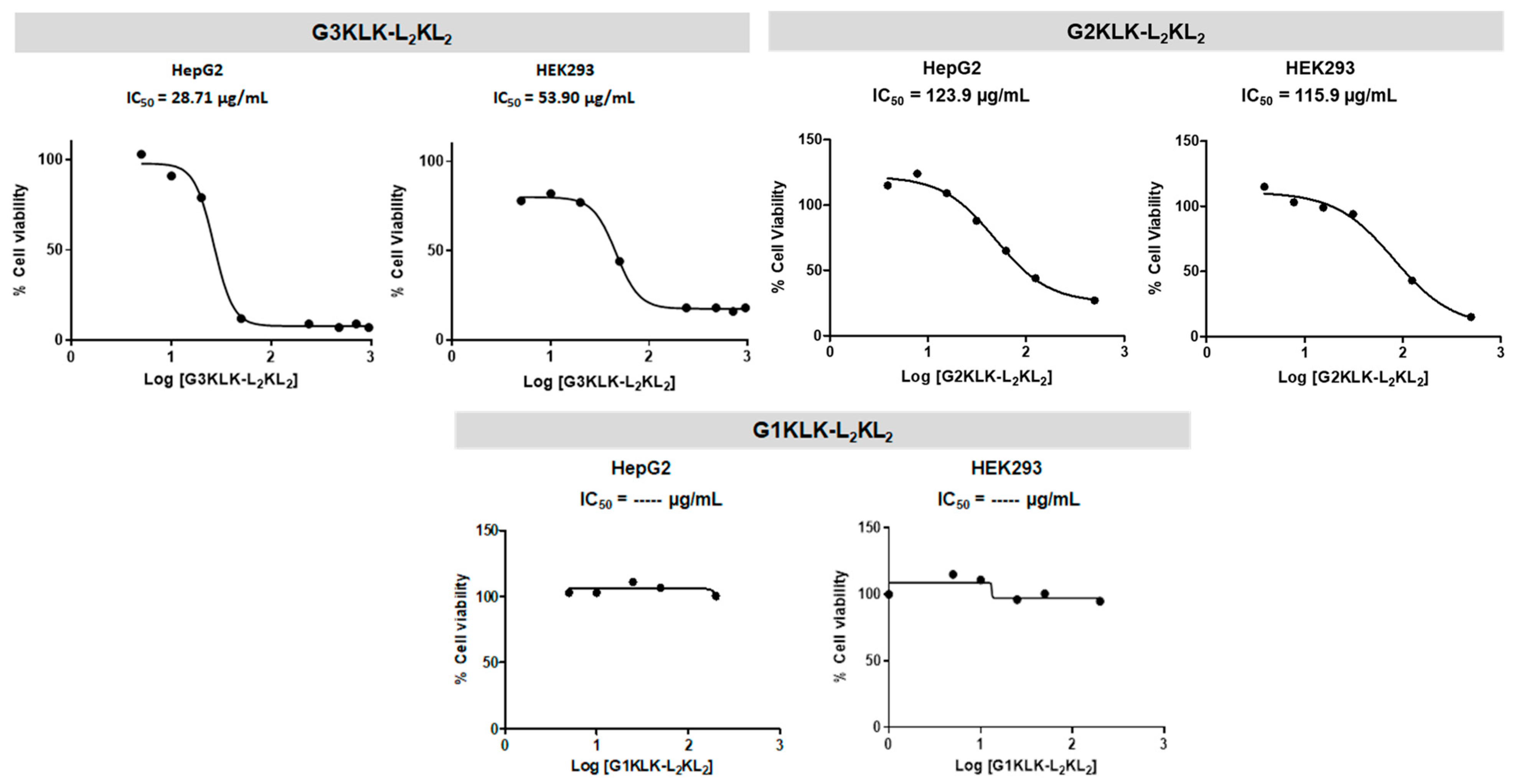
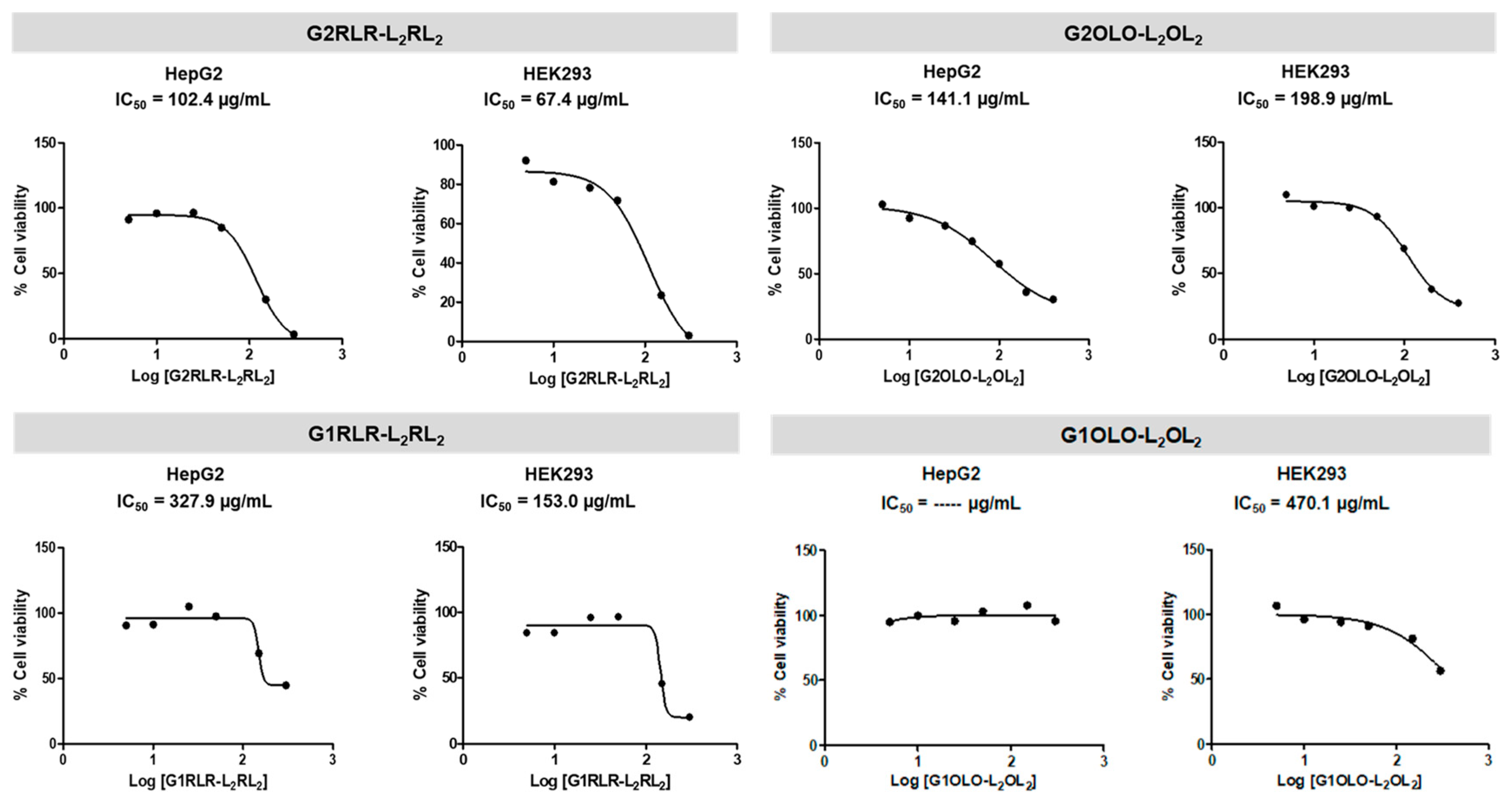
2.3. Cytotoxicity
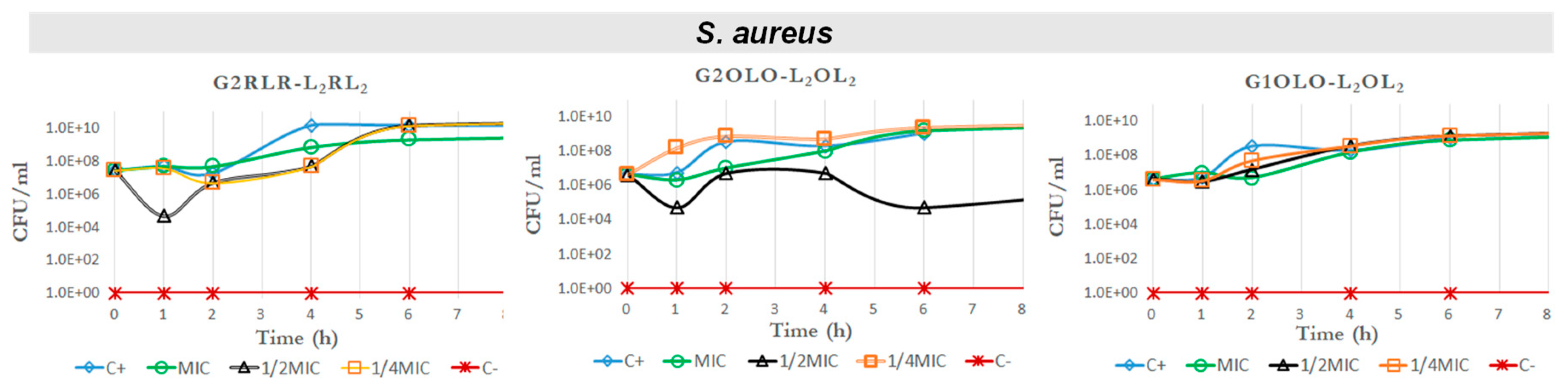
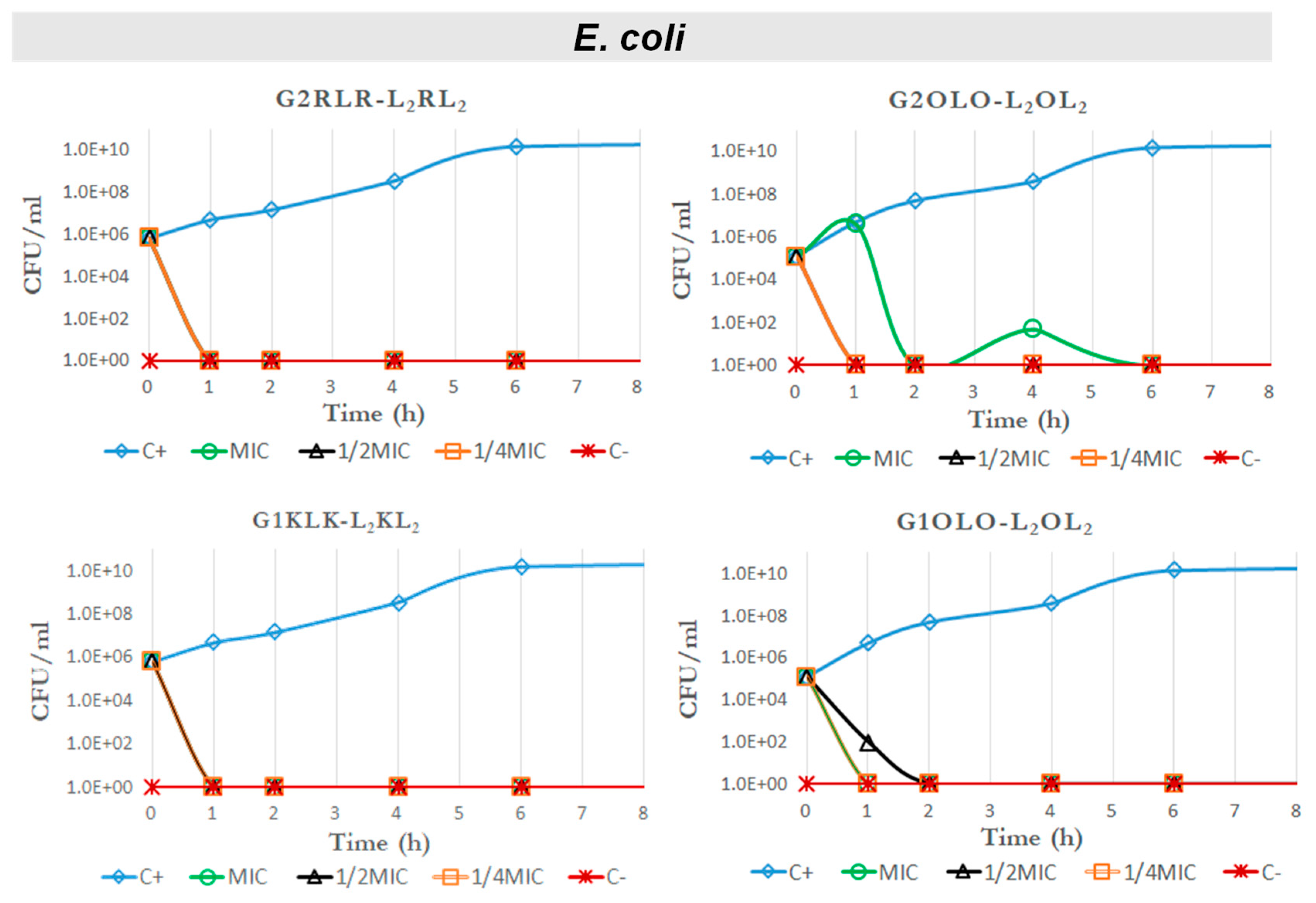
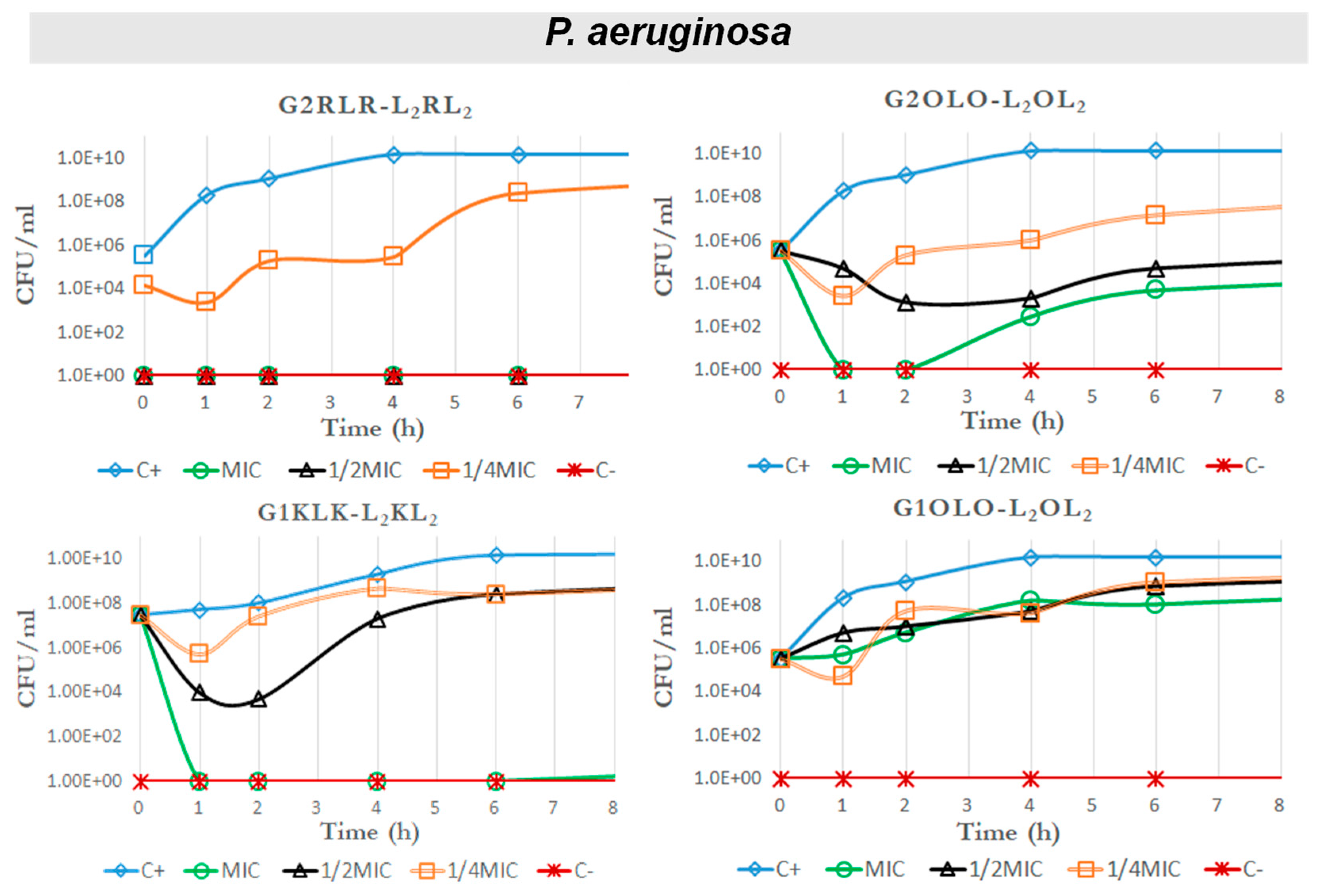
2.4. Time–Kill Curves
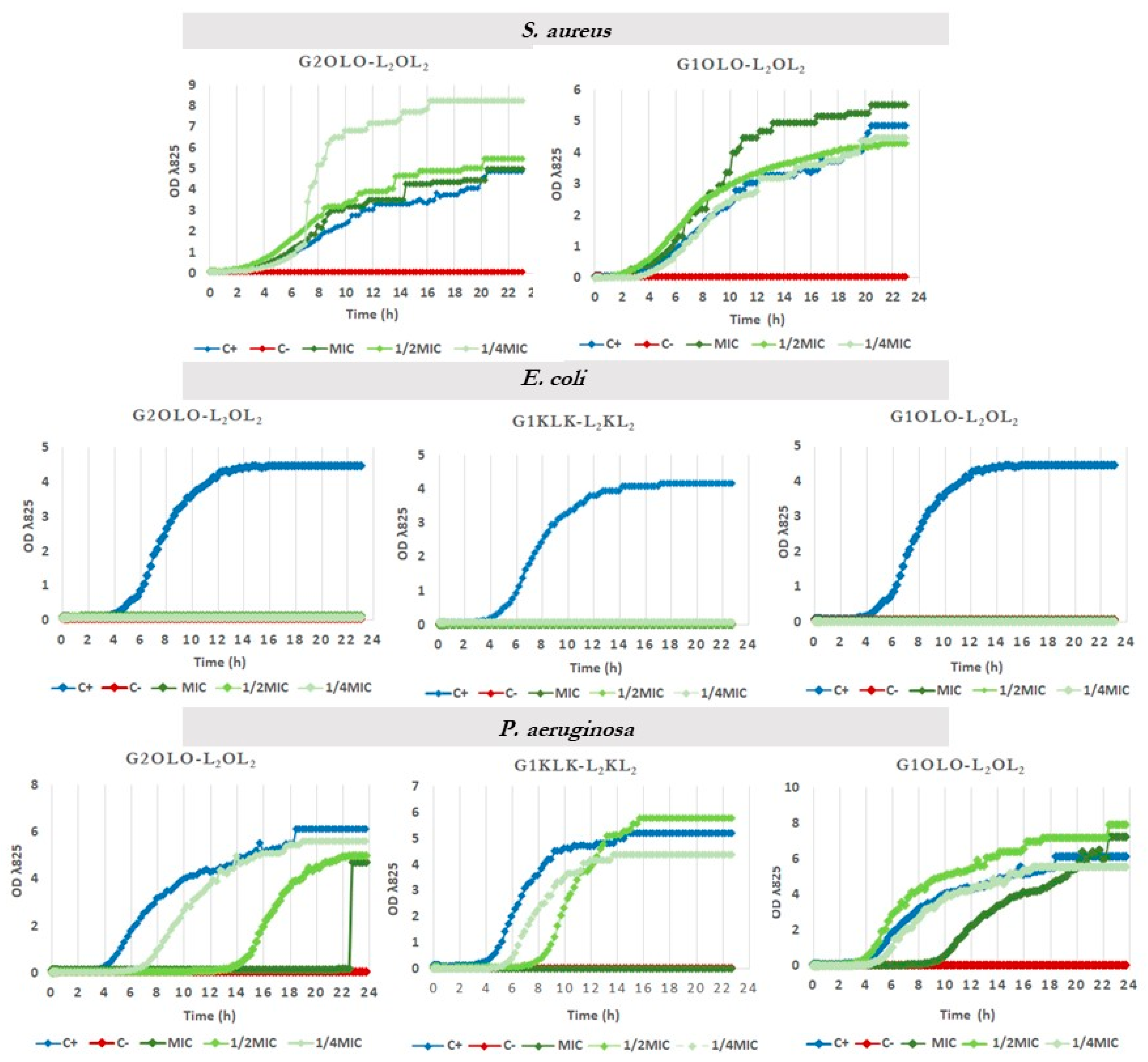
2.5. Effects on Growth Kinetics
3. Conclusions
4. Materials and Methods
4.1. Chemical Synthesis
4.2. Bacterial Strains, Media and Antimicrobial Substances
4.3. Determination of Minimum Inhibitory Concentrations
4.4. Cytotoxicity
4.5. Time–Kill Curves
4.6. Effect on Bacterial Growth Curves
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossolini, G.M.; Arena, F.; Pecile, P.; Pollini, S. Update on the antibiotic resistance crisis. Curr. Opin. Pharmacol. 2014, 18, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Boucher, H.W.; Talbot, G.H.; Benjamin, D.K., Jr.; Bradley, J.; Guidos, R.J.; Jones, R.N.; Murray, B.E.; Bonomo, R.A.; Gilbert, D. Infectious Diseases Society of America. 10×’20 progress—development of new drugs active against gram-negative bacilli: An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2013, 56, 1685–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draenert, R.; Seybold, U.; Grützner, E.; Bogner, J. Novel antibiotics: Are we still in the pre–post-antibiotic era? Infection 2015, 43, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, R.E.; Sahl, H.-G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide Antimicrobial Agents. Clin. Microbiol. Rev. 2006, 19, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudilla, H.; Fusté, E.; Cajal, Y.; Rabanal, F.; Vinuesa, T.; Viñas, M. Synergistic Antipseudomonal Effects of Synthetic Peptide AMP38 and Carbapenems. Molecules 2016, 21, 1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel Monaim, S.A.H.; Jad, Y.E.; El-Faham, A.; de la Torre, B.G.; Albericio, F. Teixobactin as a scaffold for unlimited new antimicrobial peptides: SAR study. Bioorg. Med. Chem. 2018, 26, 2788–2796. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.; Govender, T.; Kruger, H.G.; de la Torre, B.G.; Albericio, F. Short AntiMicrobial Peptides (SAMPs) as a class of extraordinary promising therapeutic agents. J. Pep. Sci. 2016, 22, 438–451. [Google Scholar] [CrossRef] [PubMed]
- Oren, Z.; Shai, Y. Mode of action of linear amphipathic α-helical antimicrobial peptides. Pep. Sci. 1998, 47, 451–463. [Google Scholar] [CrossRef]
- Comegna, D.; Benincasa, M.; Gennaro, R.; Izzo, I.; De Riccardis, F. Design, synthesis and antimicrobial properties of non-hemolytic cationic α-cyclopeptoids. Bioorg. Med. Chem. 2010, 18. [Google Scholar] [CrossRef] [PubMed]
- Claudon, P.; Violette, A.; Lamour, K.; Decossas, M.; Fournel, S.; Heurtault, B.; Godet, J.; Mély, Y.; Jamart-Grégoire, B.; Averlant-Petit, M.C. Consequences of isostructural main-chain modifications for the design of antimicrobial foldamers: Helical mimics of host-defense peptides based on a heterogeneous amide/urea backbone. Angew. Chem. Int. Ed. 2010, 49, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Mowery, B.P.; Lindner, A.H.; Weisblum, B.; Stahl, S.S.; Gellman, S.H. Structure–activity relationships among random nylon-3 copolymers that mimic antibacterial host-defense peptides. J. Am. Chem. Soc. 2009, 131, 9735–9745. [Google Scholar] [CrossRef] [PubMed]
- Stach, M.; Maillard, N.; Kadam, R.U.; Kalbermatter, D.; Meury, M.; Page, M.G.P.; Fotiadis, D.; Darbre, T.; Reymond, J.-L. Membrane disrupting antimicrobial peptide dendrimers with multiple amino termini. MedChemComm 2012, 3, 86–89. [Google Scholar] [CrossRef]
- Stach, M.; Siriwardena, T.N.; Koehler, T.; van Delden, C.; Darbre, T.; Reymond, J.-L. Combining Topology and Sequence Design for the Discovery of Potent Antimicrobial Peptide Dendrimers against Multidrug-Resistant Pseudomonas aeruginosa. Angew. Chem. Int. Ed. 2014, 53, 12827–12831. [Google Scholar] [CrossRef] [PubMed]
- Siriwardena, T.N.; Luescher, A.; Koehler, T.; van Delden, C.; Javor, S.; Reymond, J.L. Antimicrobial Peptide Dendrimer Chimera. Helv. Chim. Acta 2019, 102, e1900034. [Google Scholar] [CrossRef]
- Ghobrial, O.; Derendorf, H.; Hillman, J.D. Human serum binding and its effect on the pharmacodynamics of the lantibiotic MU1140. Eur. J. Phar. Sci. 2010, 41, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Eagle, H.; Musselman, A.D. The rate of bactericidal action of penicillin in vitro as a function of its concentration, and its paradoxically reduced activity at high concentrations against certain organisms. J. Exp. Med. 1948, 88, 99–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jad, Y.E.; El-Faham, A.; De La Torre, B.G.; Albericio, F. Solid-phase peptide synthesis, the state of the art: Challenges and opportunities. Pept.-Based Drug Discov. 2017, 59, 518–550. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; CLSI documents 2016; 26th Edition Information Supplement M100-S127:74-80; Clinical and Laboratory Standards Institute: Annapolis Junction, MD, USA, 2016. [Google Scholar]
- Tolosa, L.; Donato, M.T.; Gómez-Lechón, M.J. General cytotoxicity assessment by means of the MTT assay. In Protocols in In Vitro Hepatocyte Research; Springer: Berlin/Heidelberg, Germany, 2015; pp. 333–348. [Google Scholar]
- Lora-Tamayo, J.; Murillo, O.; Ariza, J. Clinical Use of Colistin in Biofilm-Associated Infections. Adv. Exp. Med. Biol. 2019, 1145, 181–195. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Dendrimer Sequence | Short Name | Residues (Basic/Hydrophobic) | Positive Charges | |
|---|---|---|---|---|
| 1 | (KLK)8(KKLK)4(KKLK)2KLLKLL-NH2 | G3KLK-L2KL2 | 54 (29/25) | 37 |
| 2 | (KLK)4(KKLK)2KLLKLL-NH2 | G2KLK-L2KL2 | 26 (13/13) | 17 |
| 3 | (Ac-KLK)4(KKLK)2KLLKLL-NH2 a | Ac-G2KLK-L2KL2 | 30 a (13/17) | 13 |
| 4 | (Hx-KLK)4(KKLK)2KLLKLL-NH2 | Hx-G2KLK-L2KL2 | 30 a (13/17) | 13 |
| 5 | (Dd-KLK)4(KKLK)2KLLKLL-NH2 | Dd-G2KLK-L2KL2 | 30 a (13/17) | 13 |
| 6 | (RLR)4(KRLR)2KLLRLL-NH2 | G2RLR-L2RL2 | 26 (13/13) | 17 |
| 7 | (OLO)4(KOLO)2KLLOLL-NH2 | G2OLO-L2OL2 | 26 (13/13) | 17 |
| 8 | (KLK)2KLLKLL-NH2 | G1KLK-L2KL2 | 12 (5/7) | 7 |
| 9 | (RLR)2KLLRLL-NH2 | G1RLR-L2RL2 | 12 (5/7) | 7 |
| 10 | (OLO)2KLLOLL-NH2 | G1OLO-L2OL2 | 12 (5/7) | 7 |
| Amtimicrobial Agents | Gram-Positive | Gram-Negative | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. aureus ATCC 29213 | B. subtilis ATCC 6051 | E. faecium ATCC 35667 | E. coli ATCC 25822 | P. aeruginosa ATCC 27853 | |||||||||||
| MIC | 50% HS * | MBC | MIC | 50% HS * | MBC | MIC | 50% HS * | MBC | MIC | 50% HS * | MBC | MIC | 50% HS | MBC | |
| G3KLK-L2KL2 | 16 | 16 | 16 | 4–8 | 8 | 8 | 64 | 64 | 64 | 32 | 32 | 32 | 16 | 16 | 16 |
| G2KLK-L2KL2 | 32 | 32 | 32 | 8 | 8 | 8 | 32 | 64 | 32 | 16 | 32 | 32 | 8–16 | 16 | 16 |
| Ac-G2KLK-L2KL2 | 64 | 64 | 128 | 8–16 | 16 | 16 | 64 | 128 | 128 | 8–16 | 16 | 16 | 8–16 | 16 | 16 |
| Hx-G2KLK-L2KL2 | 128 | 256 | 128 | 64 | 64 | 128 | 32 | 32 | 64 | 64 | 64 | 64 | 64 | 64 | 64 |
| Dd-G2KLK-L2KL2 | >128 | ND | ND | >128 | ND | ND | >128 | ND | ND | >128 | ND | ND | >128 | ND | ND |
| G2RLR-L2RL2 | 32 | 32 | 32 | 16 | 32 | 16 | 16 | 32 | 16 | 64 | 64 | 64 | 64 | 64 | 64 |
| G2OLO-L2OL2 | 32 | 32 | 32 | 32 | 32 | 32 | 8 | 16 | 8 | 32 | 32 | 32 | 32 | 32 | 32 |
| G1KLK-L2KL2 | 128 | >128 | 128 | 64 | 128 | 64 | >128 | ND | ND | 32 | 32 | 32 | 32 | 32 | 32 |
| G1RLR-L2RL2 | 16 | 32 | 16 | 32 | 32 | 32 | 4 | 8 | 4 | 8 | 16 | 8 | 16 | 32 | 16 |
| G1OLO-L2OL2 | 64 | 64 | 64 | 32 | 32 | 32 | 16 | 16 | 16 | 8 | 8 | 8 | 16 | 16 | 16 |
| Meropenem | 0.25 | ND | ND | 0.062 | ND | ND | <0.25 | ND | ND | 0.125 | ND | ND | 1 | ND | ND |
| Isolates | G3KLK -L2KL2 | G2KLK -L2KL2 | Ac-G2KLK -L2KL2 | Hx-G2KLK -L2KL2 | G2RLR-L2RL2 | G2OLO -L2OL2 | G1RLR-L2RL2 | G1OLO-L2OL2 | Van | Amp |
|---|---|---|---|---|---|---|---|---|---|---|
| B11970 | 32 | 64 | 128 | 64 | 32 | 64 | 16 | 64 | 1 | >512 |
| P10781 | 16 | 64 | 128 | 64 | 64 | 64 | 32 | 64 | 1 | >512 |
| P10747 | 16 | 64 | 128 | 64 | 64 | 64 | 32 | 64 | 1 | >512 |
| S37938 | 32 | 64 | 128 | 64 | 32 | 64 | 32 | 64 | 1 | >512 |
| S18155 | 32 | 64 | 128 | 64 | 64 | 64 | 16 | 64 | 1 | >512 |
| B13178 | 32 | 64 | 128 | 64 | 32 | 64 | 16 | 64 | 1 | >512 |
| 440260 | 32 | 64 | 128 | 64 | 32 | 64 | 16 | 64 | 1 | >512 |
| S18970 | 32 | 64 | 128 | 64 | 32 | 64 | 16 | 64 | 1 | >512 |
| P11520 | 16 | 64 | 128 | 64 | 32 | 64 | 16 | 64 | 1 | 512 |
| T5683 | 32 | 64 | 128 | 64 | 64 | 64 | 16 | 64 | 1 | >512 |
| Isolates | G3KLK -L2KL2 | G2KLK -L2KL2 | Ac-G2KLK -L2KL2 | Hx-G2KLK -L2KL2 | G2RLR -L2RL2 | G2OLO -L2OL2 | G1RLR-L2RL2 | G1OLO-L2OL2 | Van |
|---|---|---|---|---|---|---|---|---|---|
| 951245262 | 16 | 64 | 64 | 128 | 16 | 32 | 64 | 16 | >128 |
| 951234856 | 32 | 32 | 64 | 64 | 32 | 64 | 32 | 16 | >128 |
| 951208931 | 16 | 32 | 128 | 64 | 16 | 32 | 32 | 32 | >128 |
| 938636470 | 32 | 32 | 64 | 128 | 16 | 64 | 64 | 16 | >128 |
| 938666613 | 16 | 32 | 128 | 64 | 16 | 32 | 32 | 32 | >128 |
| 938600912 | 16 | 32 | 64 | 64 | 16 | 32 | 32 | 16 | >128 |
| 938072607 | 32 | 32 | 128 | 64 | 32 | 64 | 32 | 32 | >128 |
| 944414000 | 32 | 32 | 128 | 64 | 16 | 64 | 32 | 32 | >128 |
| 945530665 | 32 | 32 | 128 | 64 | 16 | 32 | 32 | 32 | >128 |
| U43821 | 32 | 32 | 128 | 64 | 16 | 32 | 32 | 32 | >128 |
| Isolates | G3KLK -L2KL2 | G2KLK -L2KL2 | Ac-G2KLK -L2KL2 | Hx-G2KLK -L2KL2 | G2RLR-L2RL2 | G2OLO -L2OL2 | G1RLR-L2RL2 | G1OLO-L2OL2 | Merop |
|---|---|---|---|---|---|---|---|---|---|
| VIM-1 BM-12 | >256 | 64 | 64 | 128 | 64 | 64 | 8 | 8 | >32 |
| NDM4-FEK | 256 | 32 | 64 | 64 | 32 | 32 | 8 | 8 | >32 |
| KPC L2l | 32 | 32 | 64 | 64 | 64 | 32 | 8 | 8 | 16 |
| OXA-48 501 | 16 | 32 | 32 | 128 | 64 | 16 | 16 | 16 | 2 |
| IMP JAP | 64 | 32 | 64 | >256 | 64 | 8 | 8 | 8 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramchuran, E.J.; Pérez-Guillén, I.; Bester, L.A.; Khan, R.; Albericio, F.; Viñas, M.; de la Torre, B.G. Super-Cationic Peptide Dendrimers—Synthesis and Evaluation as Antimicrobial Agents. Antibiotics 2021, 10, 695. https://doi.org/10.3390/antibiotics10060695
Ramchuran EJ, Pérez-Guillén I, Bester LA, Khan R, Albericio F, Viñas M, de la Torre BG. Super-Cationic Peptide Dendrimers—Synthesis and Evaluation as Antimicrobial Agents. Antibiotics. 2021; 10(6):695. https://doi.org/10.3390/antibiotics10060695
Chicago/Turabian StyleRamchuran, Estelle J., Isabel Pérez-Guillén, Linda A. Bester, René Khan, Fernando Albericio, Miguel Viñas, and Beatriz G. de la Torre. 2021. "Super-Cationic Peptide Dendrimers—Synthesis and Evaluation as Antimicrobial Agents" Antibiotics 10, no. 6: 695. https://doi.org/10.3390/antibiotics10060695
APA StyleRamchuran, E. J., Pérez-Guillén, I., Bester, L. A., Khan, R., Albericio, F., Viñas, M., & de la Torre, B. G. (2021). Super-Cationic Peptide Dendrimers—Synthesis and Evaluation as Antimicrobial Agents. Antibiotics, 10(6), 695. https://doi.org/10.3390/antibiotics10060695