
Dynamics of Diversity and Abundance of Sulfonamide Resistant Bacteria in a Silt Loam Soil Fertilized by Compost


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microcosm Experiment
2.2. Sulfadiazine Quantitation
2.3. TC-DNA Extraction, Real Time qPCR of sul1, sul2 and 16S rRNA Gene
2.4. High Throughput Sequencing Analysis of Bacterial sul1, sul2 and 16S rRNA Gene Amplicon
3. Results
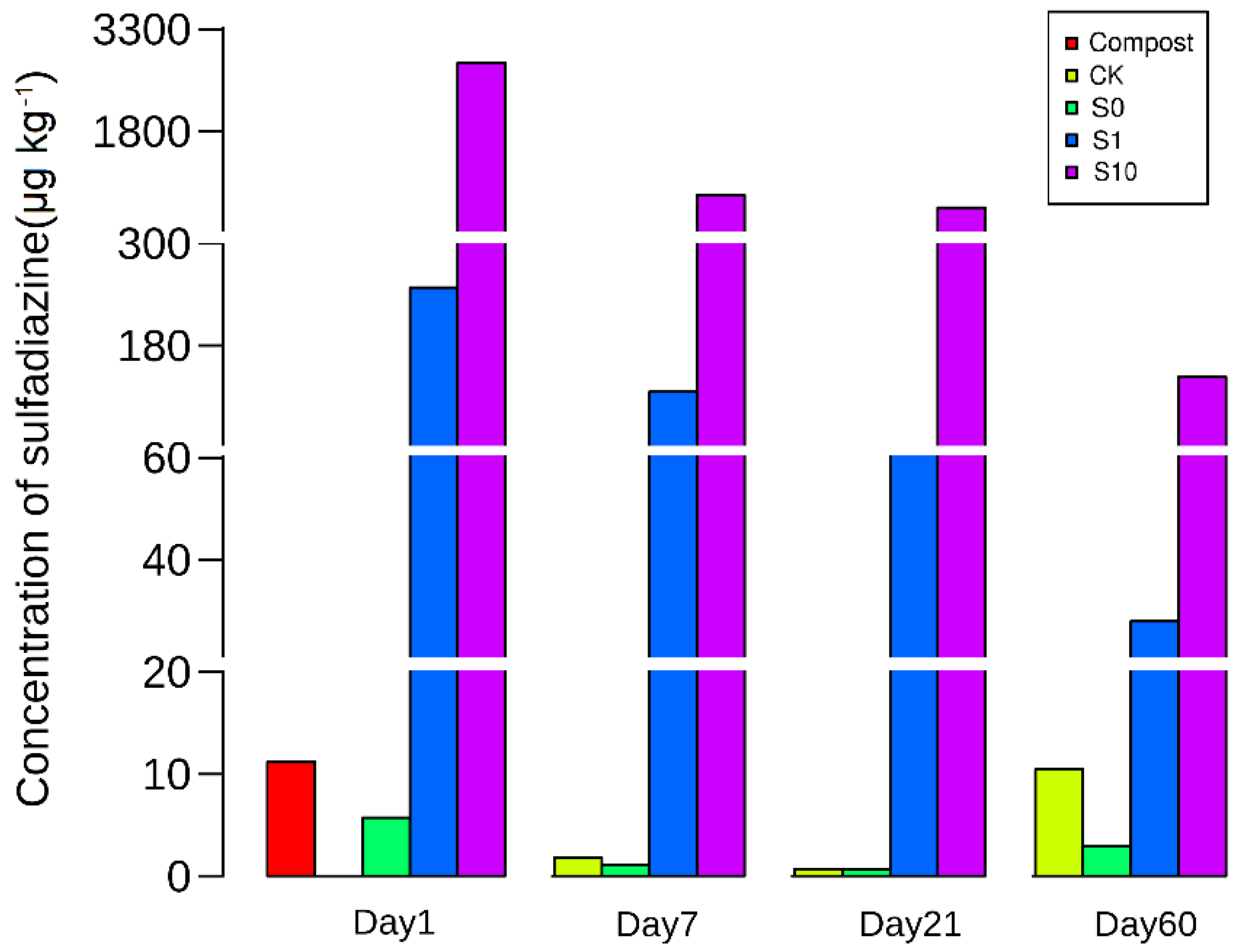
3.1. Concentration of Sulfadiazine
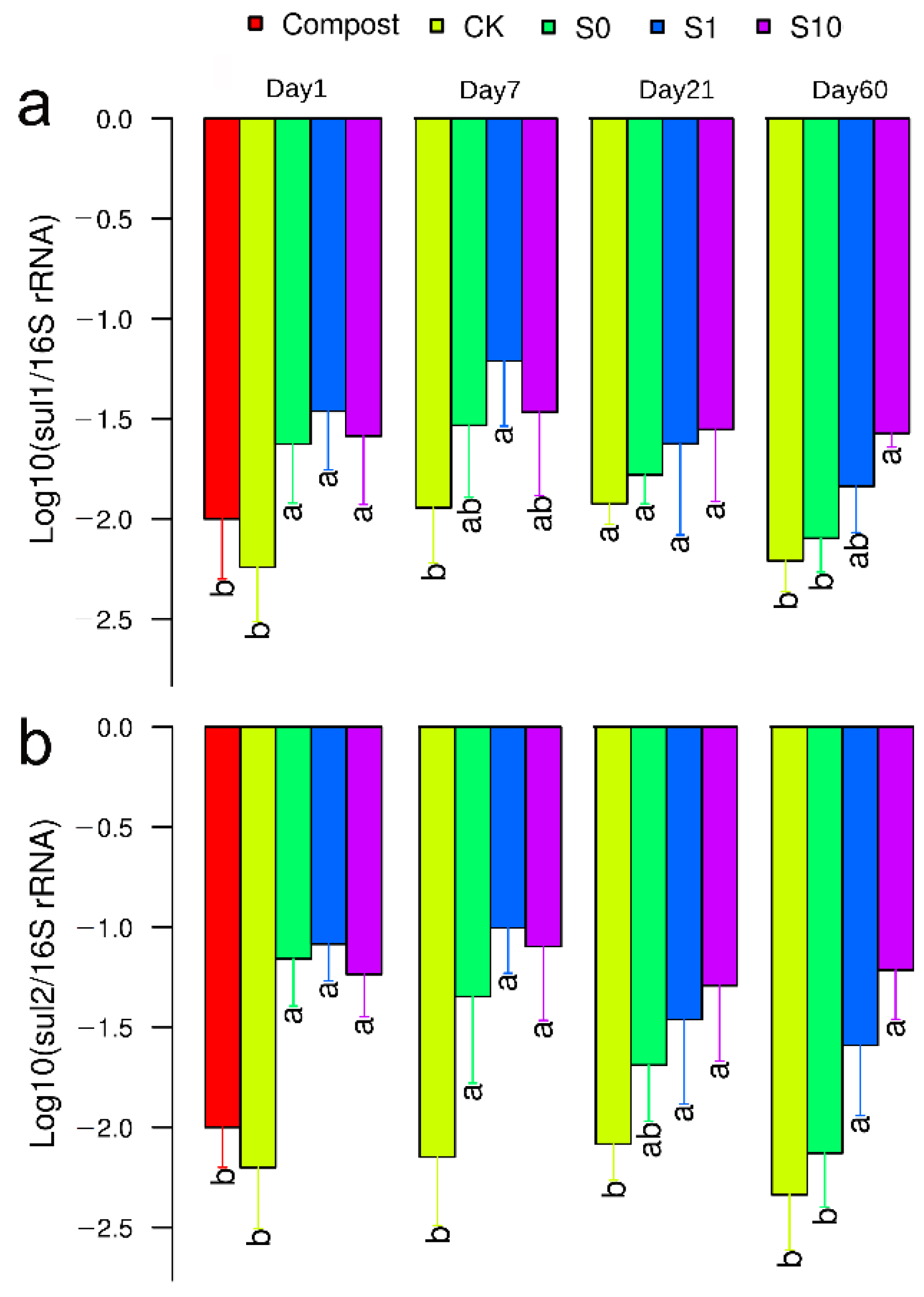
3.2. Abundance of Bacterial sul1 and sul2 Genes
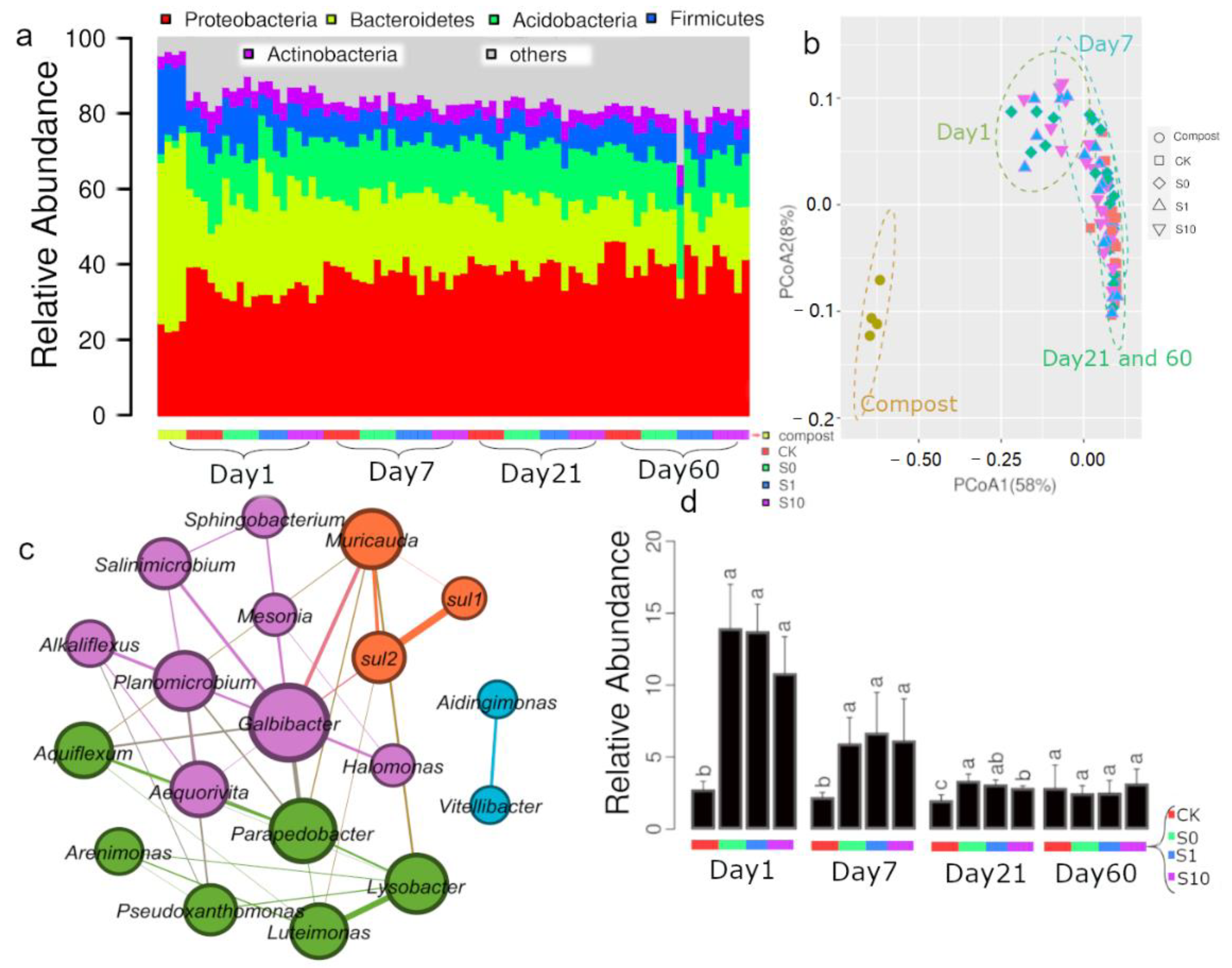
3.3. Bacterial Community Composition
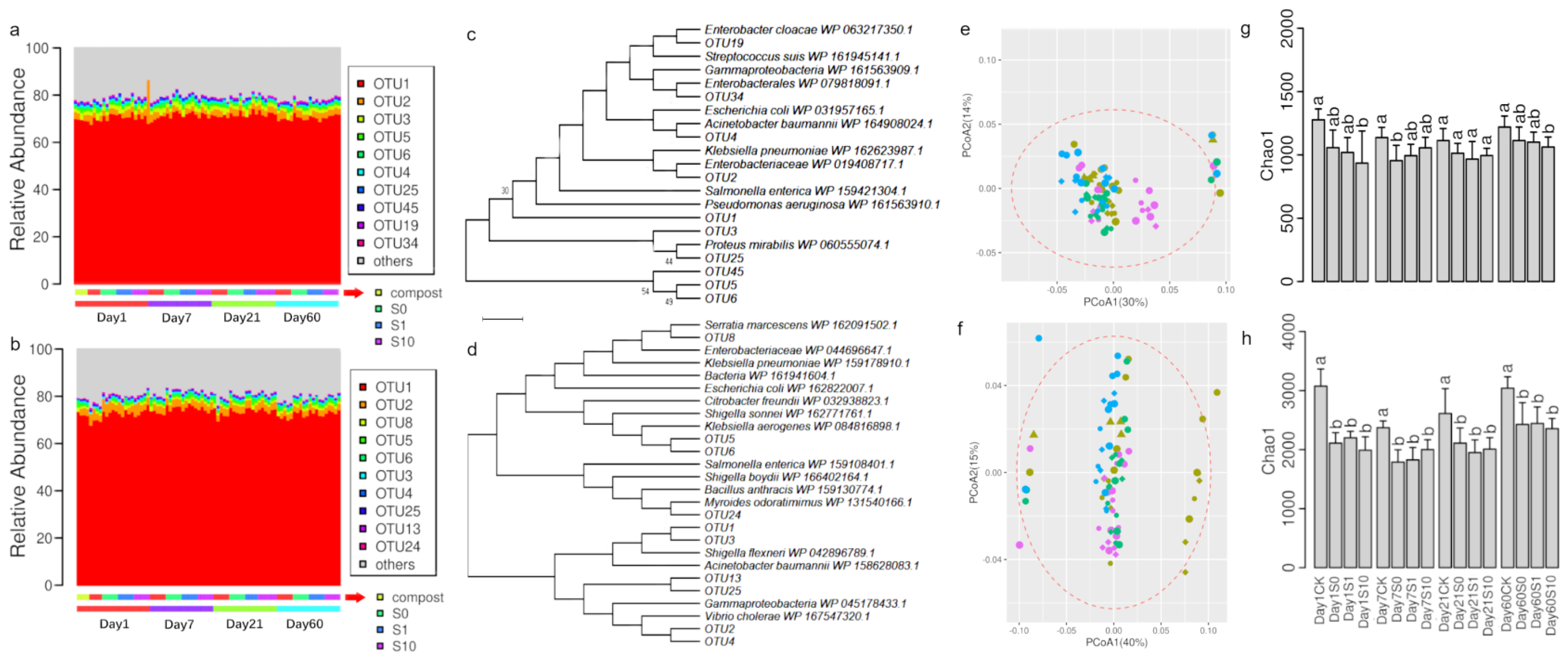
3.4. Diversity of sul1 and sul2 Genes
4. Discussion
4.1. Diversities of sul1 or sul2 Were Extremely High but Only Few Common Dominant Genotypes were Prevalent in Soil or Compost
4.2. Compost Fertilization Elevated the Abundance of sul1 and sul2 in Soils, and the Co-Introduced Sulfadiazine may Facilitate the Persistence of Such Resistance
4.3. Resilience of Soil Bacterial Community to the Perturbation of Compost
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, Q.Q.; Ying, G.G.; Pan, C.G.; Liu, Y.S.; Zhao, J.L. Comprehensive evaluation of antibiotics emission and fate in the river basins of china: Source analysis, multimedia modeling, and linkage to bacterial resistance. Environ. Sci. Technol. 2015, 49, 6772–6782. [Google Scholar] [CrossRef]
- Yao, L.H.; Li, Y.; Li, Z.Q.; Shen, D.S.; Feng, H.J.; Zhou, H.H.; Wang, M.Z. Prevalence of fluoroquinolone, macrolide and sulfonamide-related resistance genes in landfills from East China, mainly driven by MGEs. Ecotoxicol. Environ. Saf. 2019, 190, 110131. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Johnson, T.A.; Su, J.Q.; Qiao, M.; Guo, G.X.; Stedtfeld, R.D.; Hashsham, S.A.; Tiedje, J.M. Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proc. Natl. Acad. Sci. USA 2013, 110, 3435–3440. [Google Scholar] [CrossRef]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.A.; Dantas, G. The shared antibiotic resistome of soil bacteria and human pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef]
- Liu, Y.W.; Cheng, D.M.; Xue, J.M.; Weaver, L.; Wakelin, S.A.; Feng, Y.; Li, Z.J. Changes in microbial community structure during pig manure composting and its relationship to the fate of antibiotics and antibiotic resistance genes. J. Hazard. Mater. 2020, 389, 122082. [Google Scholar] [CrossRef]
- Pruden, A.; Pei, R.T.; Storteboom, H.; Carlson, K. Antibiotic resistance genes as emerging contaminants: Studies in northern Colorado. Environ. Sci. Technol. 2006, 40, 7445–7450. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, F.; Udikovickolic, N.; Andrew, S.M.; Handelsman, J. Diverse antibiotic resistance genes in dairy cow manure. mBio 2014, 5, e01017-13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Gillings, M.; Simonet, P.; Stekel, D.; Banwart, S.; Penuelas, J. Microbial mass movements. Science 2017, 357, 1099–1100. [Google Scholar] [CrossRef]
- Mullen, R.A.; Hurst, J.J.; Naas, K.M.; Sassoubre, L.M.; Aga, D.S. Assessing uptake of antimicrobials by Zea mays L. and prevalence of antimicrobial resistance genes in manure-fertilized soil. Sci. Total Environ. 2018, 646, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.C.; Radl, V.; Schloterhai, B.; Jechalke, S.; Heuer, H.; Smalla, K.; Schloter, M. Dynamics of soil bacterial communities in response to repeated application of manure containing sulfadiazine. PLoS ONE 2014, 9, e92958. [Google Scholar] [CrossRef] [PubMed]
- Urra, J.; Alkorta, I.; Lanzén, A.; Mijangos, I.; Garbisu, C. The application of fresh and composted horse and chicken manure affects soil quality, microbial composition and antibiotic resistance. Appl. Soil Ecol. 2019, 135, 73–84. [Google Scholar] [CrossRef]
- Lin, H.; Sun, W.C.; Yu, Q.G.; Ma, J.W. Acidic conditions enhance the removal of sulfonamide antibiotics and antibiotic resistance determinants in swine manure. Environ. Pollut. 2020, 263, 114439. [Google Scholar] [CrossRef] [PubMed]
- Wolters, B.; Ding, G.C.; Kreuzig, R.; Smalla, K. Full-scale mesophilic biogas plants using manure as c-source: Bacterial community shifts along the process cause changes in the abundance of resistance genes and mobile genetic elements. FEMS Microbiol. Ecol. 2016, 92, fiv163. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Sun, W.; Gu, J.; Wang, X.J.; Sun, J.J.; Yin, Y.N.; Duan, M.L. Variable effects of oxytetracycline on antibiotic resistance gene abundance and the bacterial community during aerobic composting of cow manure. J. Hazard. Mater. 2016, 315, 61–69. [Google Scholar] [CrossRef]
- Liu, H.; Pu, C.J.; Yu, X.L.; Sun, Y.; Chen, J.H. Removal of tetracyclines, sulfonamides, and quinolones by industrial-scale composting and anaerobic digestion processes. Environ. Sci. Pollut. Res. 2018, 25, 35835–35844. [Google Scholar] [CrossRef]
- Lin, H.; Chapman, S.J.; Freitag, T.E.; Kyle, C.; Ma, J.W.; Yang, Y.Y.; Zhang, Z.L. Fate of tetracycline and sulfonamide resistance genes in a grassland soil amended with different organic fertilizers. Ecotoxicol. Environ. Saf. 2019, 170, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Yang, X.H.; Jiao, S.J.; Zhang, J.; Ye, B.P.; Gao, S.X. Sulfonamide-resistant bacteria and their resistance genes in soils fertilized with manures from Jiangsu Province, Southeastern China. PLoS ONE 2014, 9, e112626. [Google Scholar] [CrossRef]
- Wu, M.; Han, H.; Zheng, X.N.; Bai, M.H.; Xu, T.; Ding, G.C.; Li, J. Dynamics of oxytetracycline and resistance genes in soil under long-term intensive compost fertilization in Northern China. Environ. Sci. Pollut. Res. 2019, 26, 21381–21393. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Junker, E.; Helmuth, R. Incidence of the recently described sulfonamide resistance gene sul3 among german salmonella enterica strains isolated from livestock and food. Antimicrob. Agents Chemother. 2004, 48, 2712–2715. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Hu, H.W.; Chen, Q.L.; Singh, B.K.; Yan, H.; Chen, D.; He, J.Z. Transfer of antibiotic resistance from manure-amended soils to vegetable microbiomes. Environ. Int. 2019, 130, 104912. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.L.; An, X.L.; Li, H.; Zhu, Y.G.; Su, J.Q.; Cui, L. Do manure-borne or indigenous soil microorganisms influence the spread of antibiotic resistance genes in manured soil? Soil Biol. Biochem. 2017, 114, 229–237. [Google Scholar] [CrossRef]
- Tien, Y.C.; Li, B.; Zhang, T.; Scott, A.; Murray, R.; Sabourin, L.; Marti, R.; Topp, E. Impact of dairy manure pre-application treatment on manure composition, soil dynamics of antibiotic resistance genes, and abundance of antibiotic-resistance genes on vegetables at harvest. Sci. Total Environ. 2017, 581–582, 32–39. [Google Scholar] [CrossRef]
- Liu, W.B.; Ling, N.; Guo, J.J.; Ruan, Y.; Wang, M.; Shen, Q.R.; Guo, S.W. Dynamics of the antibiotic resistome in agricultural soils amended with different sources of animal manures over three consecutive years. J. Hazard. Mater. 2021, 401, 123399. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wei, C.J.; Zhang, W.J.; Liu, Y.W.; Li, Z.J.; Hu, H.Y.; Xue, J.M.; Davis, M. A simple and economic method for simultaneous determination of 11 antibiotics in manure by solid-phase extraction and high-performance liquid chromatography. J. Soils Sediments 2016, 16, 2242–2251. [Google Scholar] [CrossRef]
- Suzuki, M.T.; Taylor, L.T.; DeLong, E.F. Quantitative analysis of small-subunit rRNA genes in mixed microbial populations via 5′-nuclease assays. Appl. Environ. Microbiol. 2000, 66, 4605–4614. [Google Scholar] [CrossRef]
- Heuer, H.; Smalla, K. Manure and sulfadiazine synergistically increased bacterial antibiotic resistance in soil over at least two months. Environ. Microbiol. 2007, 9, 657–666. [Google Scholar] [CrossRef]
- Heuer, H.; Focks, A.; Lamshöft, M.; Smalla, K.; Matthies, M.; Spiteller, M. Fate of sulfadiazine administered to pigs and its quantitative effect on the dynamics of bacterial resistance genes in manure and manured soil. Soil Biol. Biochem. 2008, 40, 1892–1900. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Li, H.X.; Ding, X.Y.; Chen, C.; Zheng, X.N.; Han, H.; Li, C.N.; Gong, J.Y.; Xu, T.; Li, Q.X.; Ding, G.C.; et al. Enrichment of phosphate solubilizing bacteria during late developmental stages of eggplant (Solanum melongena L.). FEMS Microbiol. Ecol. 2019, 95, fiz023. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.C.; Heuer, H.; Smalla, K. Dynamics of bacterial communities in two unpolluted soils after spiking with phenanthrene: Soil type specific and common responders. Front. Microbiol. 2012, 3, 290. [Google Scholar] [CrossRef]
- Ding, G.C.; Bai, M.H.; Han, H.; Li, H.X.; Ding, X.Y.; Yang, H.F.; Xu, T.; Li, J. Microbial taxonomic, nitrogen cycling, and phosphorus recycling community composition during long-term organic greenhouse farming. FEMS Microbiol. Ecol. 2019, 95, fiz042. [Google Scholar] [CrossRef] [PubMed]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. In Proceedings of the Third International Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009; pp. 1–2. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Chen, Q.W.; Zhang, J.Y.; Guan, T.S.; Chen, Y.C.; Shi, W.Q. Critical roles of cyanobacteria as reservoir and source for antibiotic resistance genes. Environ. Int. 2020, 144, 106034. [Google Scholar] [CrossRef]
- Pawlowski, A.C.; Wang, W.; Koteva, K.; Barton, H.A.; McArthur, A.G.; Wright, G.D. A diverse intrinsic antibiotic resistome from a cave bacterium. Nat. Commun. 2016, 7, 13803. [Google Scholar] [CrossRef]
- Zhou, Y.T.; Niu, L.L.; Zhu, S.Y.; Lu, H.J.; Liu, W.P. Occurrence, abundance, and distribution of sulfonamide and tetracycline resistance genes in agricultural soils across china. Sci. Total Environ. 2017, 599–600, 1977–1983. [Google Scholar] [CrossRef]
- Willms, I.M.; Kamran, A.; Aßmann, N.F.; Krone, D.; Bolz, S.H.; Fiedler, F.; Nacke, H. Discovery of novel antibiotic resistance determinants in forest and grassland soil metagenomes. Front. Microbiol. 2019, 10, 460. [Google Scholar] [CrossRef]
- Johnson, C.M.; Grossman, A.D. Integrative and conjugative elements (ICEs): What they do and how they work. Annu. Rev. Genet. 2015, 49, 577–601. [Google Scholar] [CrossRef]
- Zhao, R.X.; Yu, K.; Zhang, J.Y.; Zhang, G.J.; Huang, J.; Ma, L.P.; Deng, C.F.; Li, X.Y.; Li, B. Deciphering the mobility and bacterial hosts of antibiotic resistance genes under antibiotic selection pressure by metagenomic assembly and binning approaches. Water Res. 2020, 186, 116318. [Google Scholar] [CrossRef]
- Wei, H.W.; Ma, J.Y.; Su, Y.L.; Xie, B. Effect of nutritional energy regulation on the fate of antibiotic resistance genes during composting of sewage sludge. Bioresour. Technol. 2019, 297, 122513. [Google Scholar] [CrossRef]
- Hall, J.P.J.; Harrison, E. Bacterial evolution: Resistance is a numbers game. Nat. Microbiol. 2016, 1, 16235. [Google Scholar] [CrossRef]
- Heuer, H.; Solehati, Q.; Zimmerling, U.; Kleineidam, K.; Schloter, M.; Muller, T.; Focks, A.; Thielebruhn, S.; Smalla, K. Accumulation of sulfonamide resistance genes in arable soils due to repeated application of manure containing sulfadiazine. Appl. Environ. Microbiol. 2011, 77, 2527–2530. [Google Scholar] [CrossRef]
- Jechalke, S.; Heuer, H.; Siemens, J.; Amelung, W.; Smalla, K. Fate and effects of veterinary antibiotics in soil. Trends Microbiol. 2014, 22, 536–545. [Google Scholar] [CrossRef]
- Knapp, C.W.; Engemann, C.A.; Hanson, M.L.; Keen, P.L.; Hall, K.J.; Graham, D.W. Indirect evidence of transposon-mediated selection of antibiotic resistance genes in aquatic systems at low-level oxytetracycline exposures. Environ. Sci. Technol. 2008, 42, 5348–5353. [Google Scholar] [CrossRef] [PubMed]
- Cirz, R.T.; Chin, J.K.; Andes, D.R.; de Crécy Lagard, V.; Craig, W.A.; Romesberg, F.E. Inhibition of mutation and combating the evolution of antibiotic resistance. PLoS Biol. 2005, 3, 1024–1033. [Google Scholar] [CrossRef]
- Kohanski, M.A.; DePristo, M.A.; Collins, J.J. Sublethal antibiotic treatment leads to multidrug resistance via radical-induced mutagenesis. Mol. Cell 2010, 37, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, D.M.; Rosenberg, S.M. What is mutation? A chapter in the series: How microbes ‘jeopardize’ the modern synthesis. PLoS Genet. 2019, 15, e1007995. [Google Scholar] [CrossRef] [PubMed]
- Kunin, V.; Engelbrektson, A.; Ochman, H.; Hugenholtz, P. Wrinkles in the rare biosphere: Pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 2010, 12, 118–123. [Google Scholar] [CrossRef]
- Hyma, K.E.; Lacher, D.W.; Nelson, A.M.; Bumbaugh, A.C.; Whittam, T.S. Evolutionary genetics of a new pathogenic Escherichia species: Escherichia albertii and related Shigella boydii strains. J. Bacteriol. 2005, 187, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Kuhle, V.; Abrahams, G.L.; Hensel, M. Intracellular Salmonella enterica redirect exocytic transport processes in a Salmonella pathogenicity island 2-dependent manner. Traffic 2010, 7, 716–730. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Harald, S.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed]
- Pomakova, D.K.; Hsiao, C.B.; Beanan, J.M.; Olson, R.; Macdonald, U.; Keynan, Y.; Russo, T.A. Clinical and phenotypic differences between classic and hypervirulent Klebsiella pneumonia: An emerging and under-recognized pathogenic variant. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 981–989. [Google Scholar] [CrossRef]
- Hamidian, M.; Ambrose, S.J.; Hall, R.M. A large conjugative Acinetobacter baumannii plasmid carrying the sul2 sulphonamide and strAB streptomycin resistance genes. Plasmid 2016, 87–88, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.S.; Moura, A.; Henriques, I.; Brown, C.J.; Rogers, L.M.; Top, E.M.; Correia, A. Comparative genomics of IncP-1ε plasmids from water environments reveals diverse and unique accessory genetic elements. Plasmid 2013, 70, 412–419. [Google Scholar] [CrossRef]
- Fahrenfeld, N.; Knowlton, K.; Krometis, L.A.; Hession, W.C.; Xia, K.; Lipscomb, E.; Libuit, K.; Green, B.L.; Pruden, A. Effect of manure application on abundance of antibiotic resistance genes and their attenuation rates in soil: Field-scale mass balance approach. Environ. Sci. Technol. 2014, 48, 2643–2650. [Google Scholar] [CrossRef]
- Ho, Y.B.; Zakaria, M.P.; Latif, P.A.; Saari, N. Degradation of veterinary antibiotics and hormone during broiler manure composting. Bioresour. Technol. 2013, 131, 476–484. [Google Scholar] [CrossRef]
- Yang, J.F.; Ying, G.G.; Yang, L.H.; Zhao, J.L.; Liu, F.; Tao, R.; Yu, Z.Q.; Peng, P.A. Degradation behavior of sulfadiazine in soils under different conditions. J. Environ. Sci. Health Part B 2009, 44, 241–248. [Google Scholar] [CrossRef]
- Boxall, A.B.A.; Blackwell, P.; Cavallo, R.; Kay, P.; Tolls, J. The sorption and transport of a sulphonamide antibiotic in soil systems. Toxicol. Lett. 2002, 131, 19–28. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequences of Probes | Sequences of Forward Primer (5′-3′) | Sequences of Reverse Primer (5′-3′) | References |
|---|---|---|---|---|
| 16S rRNA | CTTGTACACACCGCCCGTC | CGGTGAATACGTTCYCGG | GGWTACCTTGTTACGACTT | [25] |
| sul1 | CAGCGAGCCTTGCGGCGG | CCGTTGGCCTTCCTGTAAAG | TTGCCGATCGCGTGAAGT | [26] |
| sul2 | CGGTGCTTCTGTCTGTTTCGCGC | CGGCTGCGCTTCGATT | CGCGCGCAGAAAGGATT | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, H.; Bai, M.; Chen, Y.; Gong, Y.; Wu, M.; Yang, H.; Chen, Q.; Xu, T.; Wei, Y.; Ding, G.; et al. Dynamics of Diversity and Abundance of Sulfonamide Resistant Bacteria in a Silt Loam Soil Fertilized by Compost. Antibiotics 2021, 10, 699. https://doi.org/10.3390/antibiotics10060699
Han H, Bai M, Chen Y, Gong Y, Wu M, Yang H, Chen Q, Xu T, Wei Y, Ding G, et al. Dynamics of Diversity and Abundance of Sulfonamide Resistant Bacteria in a Silt Loam Soil Fertilized by Compost. Antibiotics. 2021; 10(6):699. https://doi.org/10.3390/antibiotics10060699
Chicago/Turabian StyleHan, Hui, Mohan Bai, Yanting Chen, Yali Gong, Ming Wu, Hefa Yang, Qing Chen, Ting Xu, Yuquan Wei, Guochun Ding, and et al. 2021. "Dynamics of Diversity and Abundance of Sulfonamide Resistant Bacteria in a Silt Loam Soil Fertilized by Compost" Antibiotics 10, no. 6: 699. https://doi.org/10.3390/antibiotics10060699
APA StyleHan, H., Bai, M., Chen, Y., Gong, Y., Wu, M., Yang, H., Chen, Q., Xu, T., Wei, Y., Ding, G., & Li, J. (2021). Dynamics of Diversity and Abundance of Sulfonamide Resistant Bacteria in a Silt Loam Soil Fertilized by Compost. Antibiotics, 10(6), 699. https://doi.org/10.3390/antibiotics10060699