
Alteration of Intestinal Microbiome of Clostridioides difficile-Infected Hamsters during the Treatment with Specific Cow Antibodies

 , and
, and
Abstract
:1. Introduction
2. Results
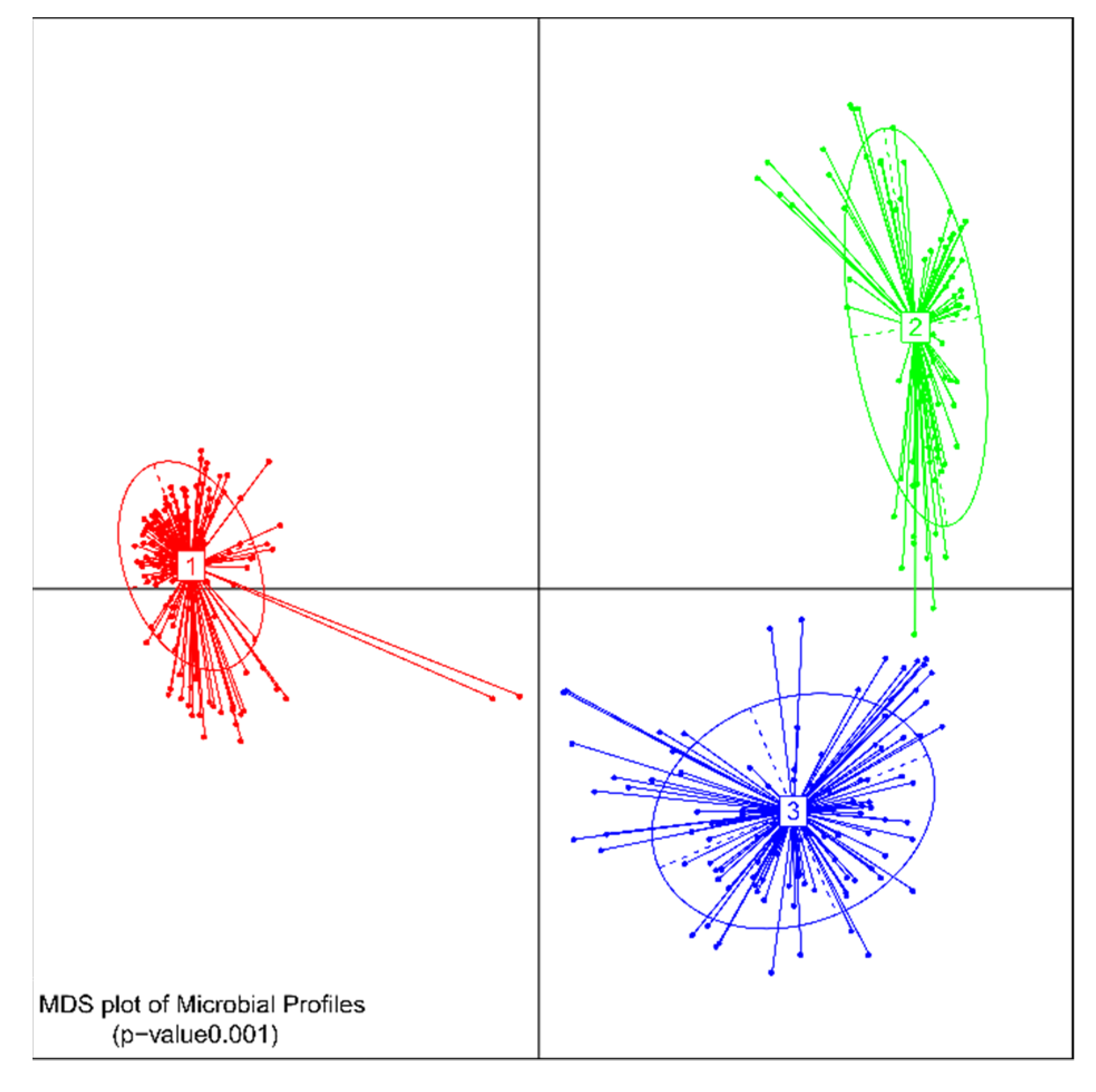
2.1. Comparison of Treatment Groups
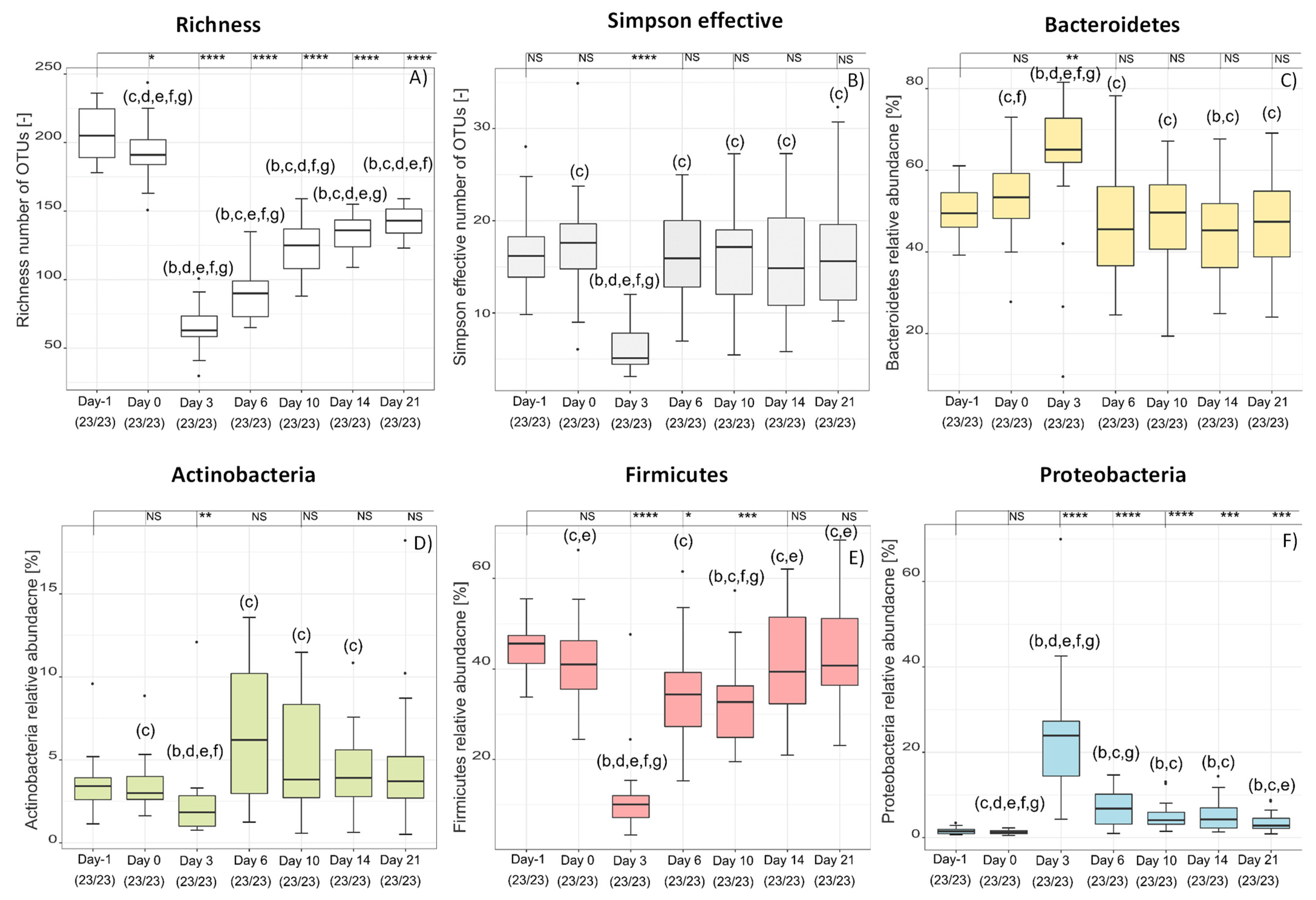
2.2. Over-Time Comparison of Treatment Groups
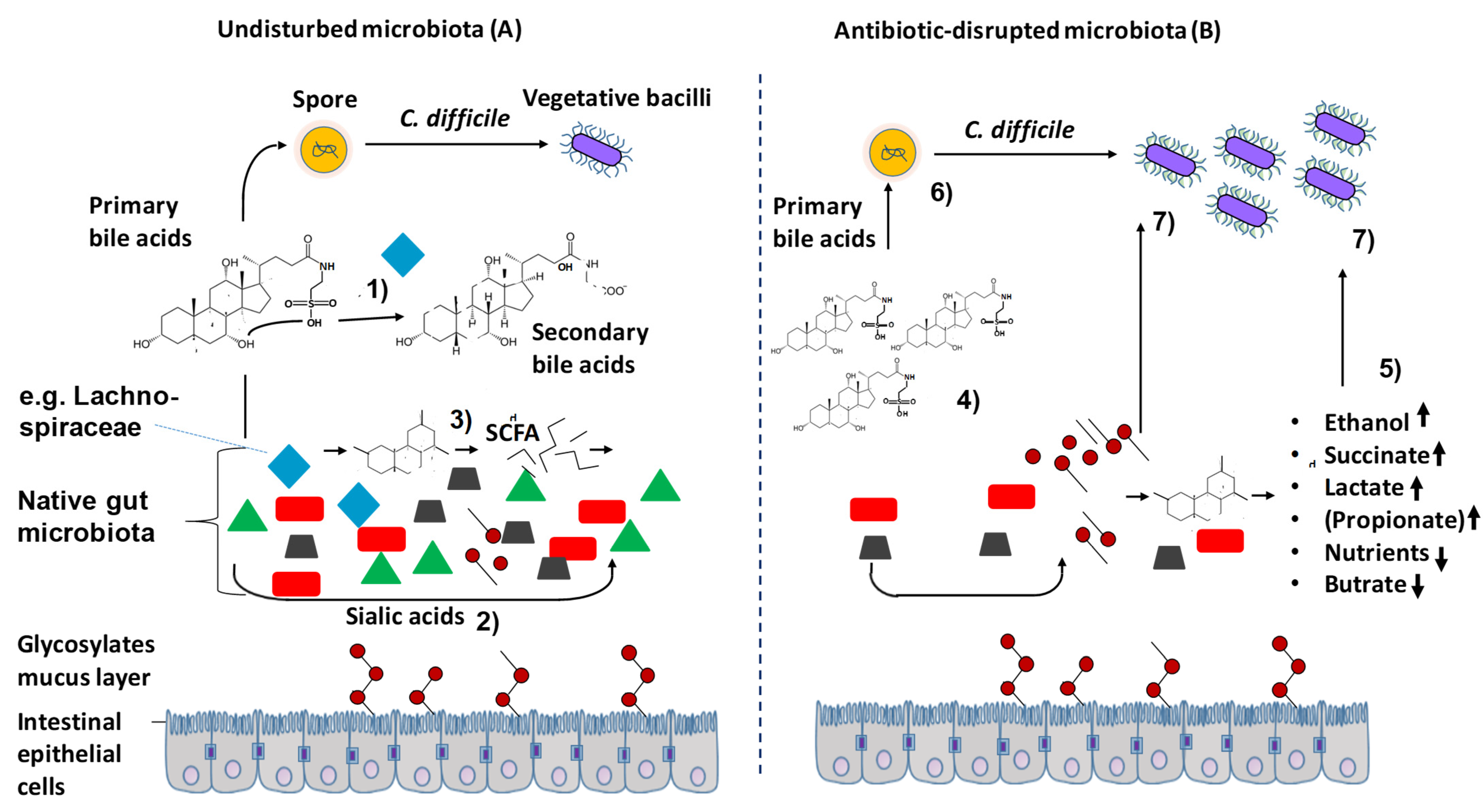
2.3. Expanding the Level of Understanding of the Role of the Microbiome in CDI
3. Materials and Methods
3.1. Study Design
3.2. Sample Collection
3.3. Microbiome Analysis by Sequencing of 16S rRNA
3.4. Data Processing
3.5. Determination of Intestinal Metabolites with Reversed-Phase High-Performance Liquid Chromatography (RP-HPLC)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phosphate [mg/mL] | Glucose [mg/mL] | Galactose [mg/mL] | Succinic Acid [mg/mL] | Lactic Acid [mg/mL] | Acetic Acid [mg/mL] | Propionic Acid [mg/mL] | Ethanol [mg/mL] | Butyric Acid [mg/mL] | Iso-Valeric Acid [mg/mL] | |
|---|---|---|---|---|---|---|---|---|---|---|
| Placebo | 2.68 | 1.23 | 0.23 | 0.09 | 0.34 | 1.77 | 0.42 | 0.18 | 0.40 | 0.60 |
| 1.44 | 0.00 | 0.21 | 2.05 | 0.25 | 3.90 | 1.15 | 0.76 | 0.25 | 0.70 | |
| 0.96 | 0.00 | 0.02 | 0.84 | 1.30 | 1.11 | 0.29 | 1.66 | 0.48 | 0.77 | |
| 1.36 | 0.00 | 0.13 | 2.01 | 0.97 | 2.37 | 1.11 | 1.43 | 0.24 | 0.50 | |
| 1.03 | 0.00 | 0.09 | 2.29 | 0.74 | 2.51 | 1.23 | 2.10 | 0.52 | 0.16 | |
| 1.06 | 0.00 | 0.18 | 1.37 | 1.10 | 2.03 | 0.96 | 3.57 | 0.48 | 0.14 | |
| 1.23 | 0.00 | 0.04 | 1.48 | 0.13 | 3.83 | 1.25 | 2.28 | 0.49 | 0.24 | |
| 1.54 | 0.24 | 0.24 | 1.87 | 1.53 | 4.04 | 1.18 | 1.89 | 0.82 | 0.32 | |
| WPI 1000 | 3.28 | 2.89 | 0.27 | 0.11 | 0.11 | 2.25 | 0.31 | 0.45 | 1.50 | 0.03 |
| 3.29 | 1.45 | 0.31 | 0.04 | 0.13 | 1.29 | 0.20 | 0.08 | 0.63 | 0.02 | |
| 2.84 | 1.74 | 0.27 | 0.03 | 0.16 | 1.29 | 0.17 | 0.05 | 0.90 | 0.70 | |
| 3.25 | 1.87 | 0.26 | 0.79 | 0.14 | 1.86 | 0.29 | 0.00 | 1.06 | 0.46 | |
| 1.60 | 0.67 | 0.16 | 0.03 | 0.13 | 0.76 | 0.11 | 0.02 | 0.49 | 0.01 | |
| WPI100 | 2.83 | 1.08 | 0.36 | 0.09 | 0.19 | 1.68 | 0.27 | 0.03 | 0.81 | 0.53 |
| 3.91 | 2.88 | 0.44 | 0.11 | 0.31 | 3.02 | 0.27 | 0.07 | 2.04 | 1.02 | |
| 1.02 | 0.04 | 0.11 | 0.19 | 0.25 | 0.64 | 0.03 | 0.96 | 0.28 | 0.02 | |
| 2.91 | 1.37 | 0.19 | 0.01 | 0.29 | 1.53 | 0.15 | 0.10 | 1.03 | 0.49 | |
| 3.06 | 1.47 | 0.23 | 0.04 | 0.18 | 1.66 | 0.18 | 0.03 | 1.05 | 0.39 | |
| 3.04 | 1.85 | 0.28 | 0.22 | 0.15 | 1.58 | 0.42 | 0.03 | 0.84 | 0.82 | |
| 3.34 | 1.98 | 0.32 | 0.05 | 0.09 | 1.86 | 0.27 | 0.28 | 1.05 | 0.61 | |
| 2.45 | 1.28 | 0.18 | 0.03 | 0.21 | 1.11 | 0.30 | 0.01 | 0.61 | 0.64 | |
| 3.08 | 1.52 | 0.51 | 0.05 | 0.12 | 1.60 | 0.31 | 0.06 | 0.99 | 0.53 | |
| WPI10000 | 3.00 | 1.85 | 0.29 | 0.27 | 0.31 | 1.73 | 0.27 | 0.12 | 1.12 | 0.04 |
| 3.02 | 1.44 | 0.59 | 0.02 | 0.10 | 1.68 | 0.20 | 0.07 | 1.00 | 0.04 | |
| 3.17 | 2.30 | 0.25 | 0.09 | 0.08 | 1.62 | 0.17 | 0.10 | 1.10 | 0.46 | |
| 3.06 | 1.76 | 0.18 | 0.01 | 0.09 | 1.09 | 0.12 | 0.01 | 0.77 | 0.44 | |
| 2.96 | 1.78 | 0.20 | 0.17 | 0.07 | 1.61 | 0.57 | 0.00 | 0.86 | 0.04 | |
| 3.08 | 1.64 | 0.27 | 0.05 | 0.13 | 1.34 | 0.25 | 0.09 | 0.75 | 0.62 | |
| 2.91 | 1.09 | 0.43 | 0.48 | 0.08 | 1.33 | 0.21 | 0.12 | 0.86 | 0.76 | |
| 2.81 | 1.80 | 0.28 | 0.04 | 0.11 | 1.44 | 0.15 | 0.02 | 0.89 | 0.44 | |
| 3.22 | 1.79 | 0.36 | 0.10 | 0.07 | 1.31 | 0.19 | 0.05 | 0.86 | 0.58 | |
| 3.48 | 2.00 | 0.29 | 0.05 | 0.16 | 1.51 | 0.20 | 0.04 | 1.25 | 0.61 | |
| Vancomycin | 1.73 | 0.00 | 0.14 | 0.82 | 0.80 | 2.56 | 1.09 | 0.64 | 1.21 | 0.33 |
| 1.67 | 0.00 | 0.06 | 1.36 | 0.41 | 2.03 | 0.26 | 1.07 | 0.98 | 0.33 | |
| 1.24 | 0.07 | 0.08 | 0.84 | 0.19 | 2.59 | 0.94 | 5.71 | 0.97 | 0.24 | |
| 2.61 | 0.77 | 0.20 | 0.03 | 0.22 | 0.84 | 0.13 | 0.14 | 0.63 | 0.02 | |
| 1.62 | 0.00 | 0.09 | 1.35 | 0.77 | 3.28 | 1.05 | 0.34 | 1.03 | 0.28 | |
| Vehicle | 1.02 | 0.00 | 0.09 | 0.43 | 0.44 | 0.71 | 0.09 | 0.25 | 0.43 | 0.58 |
| 0.90 | 0.00 | 0.13 | 0.89 | 0.00 | 1.39 | 0.47 | 1.88 | 0.35 | 0.07 | |
| 1.04 | 0.00 | 0.18 | 0.62 | 0.21 | 1.26 | 0.35 | 1.70 | 0.58 | 0.62 | |
| 0.80 | 0.10 | 0.07 | 0.13 | 0.20 | 1.20 | 0.07 | 0.74 | 0.44 | 0.04 | |
| 0.97 | 0.06 | 0.14 | 0.38 | 0.07 | 0.63 | 0.23 | 0.09 | 0.23 | 0.04 | |
| 1.28 | 0.15 | 0.23 | 1.41 | 0.25 | 3.34 | 1.30 | 1.92 | 1.07 | 0.24 |
| Variable 1 | Variable 1 | Correlation | p-Value | Number of Supporting Data |
|---|---|---|---|---|
| Phosphate/Citrate | Log value of vegetative C. difficile count | −0.86 | 2.9 × 10−13 | 43 |
| Phosphate/Citrate | Log value of C. difficile spore count | −0.89 | 8.88 × 10−16 | 43 |
| Glucose | Log value of vegetative C. difficile count | 0.85 | 1.67 × 10−09 | 31 |
| Glucose | Log value of C. difficile spore count | −0.92 | 1.05 × 10−13 | 31 |
| Galactose | Log value of vegetative C. difficile count | −0.67 | 8.79 × 10−7 | 43 |
| Galactose | Log value of C. difficile spore count | −0.68 | 5.89 × 10−7 | 43 |
| Butyric acid | Log value of vegetative C. difficile count | −0.54 | 1.78 × 10−4 | 43 |
| Butyric acid | Log value of C. difficile spore count | −0.51 | 4.83 × 10−4 | 43 |
| Ethanol | Log value of vegetative C. difficile count | 0.79 | 9.44 × 10−10 | 41 |
| Ethanol | Log value of C. difficile spore count | 0.82 | 4.03 × 10−11 | 41 |
| Succinic acid | Log value of vegetative C. difficile count | 0.74 | 1.61 × 10−8 | 43 |
| Succinic acid | Log value of C. difficile spore count | 0.77 | 1.11 × 10−9 | 43 |
| Lactic acid | Log value of vegetative C. difficile count | 0.61 | 2.10 × 10−5 | 42 |
| Lactic acid | Log value of C. difficile spore count | 0.67 | 1.24 × 10−6 | 42 |
| Richness | Log value of vegetative C. difficile count | −0.80 | 9.33 × 10−15 | 60 |
| Richness | Log value of C. difficile spore count | −0.81 | 5.55 × 10−15 | 60 |
| Shannon effective Index | Log value of vegetative C. difficile count | −0.61 | 2.22 × 10−7 | 60 |
| Shannon effective Index | Log value of C. difficile spore count | −0.63 | 8.06 × 10−8 | 60 |
| Simpson effective Index | Log value of vegetative C. difficile count | −0.41 | 1.04 × 10−3 | 60 |
| Simpson effective Index | Log value of C. difficile spore count | −0.44 | 4.86 × 10−4 | 60 |
| Simpson effective Index | Log value of C. difficile spore count | −0.80 | 9.33 × 10−15 | 60 |
References
- Lessa, F.C.; Mu, Y.; Bamberg, W.M.; Beldavs, Z.G.; Dumyati, G.K.; Dunn, J.R.; Farley, M.M.; Holzbauer, S.M.; Meek, J.I.; Phipps, E.C.; et al. Burden of Clostridium difficile infection in the United States. N. Engl. J. Med. 2015, 372, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Sartelli, M.; Malangoni, M.A.; Abu-Zidan, F.M.; Griffiths, E.A.; Di Bella, S.; McFarland, L.V.; Eltringham, I.; Shelat, V.G.; Velmahos, G.C.; Kelly, C.P.; et al. WSES guidelines for management of Clostridium difficile infection in surgical patients. World J. Emerg. Surg. 2015, 10, 38. [Google Scholar] [CrossRef]
- Hensgens, M.P.M.; Goorhuis, A.; Dekkers, O.M.; Kuijper, E.J. Time interval of increased risk for Clostridium difficile infection after exposure to antibiotics. J. Antimicrob. Chemother. 2012, 67, 742–748. [Google Scholar] [CrossRef]
- McFarland, L.V. Alternative treatments for Clostridium difficile disease: What really works? J. Med. Microbiol. 2005, 54, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidebrecht, H.-J.; Weiss, W.; Pulse, M.; Lange, A.; Gisch, K.; Kliem, H.; Mann, S.; Pfaffl, M.; Kulozik, U.; von Eichel-Streiber, C. Treatment and prevention of recurrent Clostridium difficile infection with functionalized bovine antibody-enriched whey in a hamster primary infection model. Toxins 2019, 11, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dissel, J.T.; de Groot, N.; Hensgens, C.M.; Numan, S.; Kuijper, E.J.; Veldkamp, P.; van’t Wout, J. Bovine antibody-enriched whey to aid in the prevention of a relapse of Clostridium difficile-associated diarrhoea: Preclinical and preliminary clinical data. J. Med. Microbiol. 2005, 54, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Lyerly, D.M.; Bostwick, E.; Binion, S.; Wilkins, T.D. Passive immunization of hamsters against disease caused by Clostridium difficile by use of bovine immunoglobulin G concentrate. Infect. Immun. 1991, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sponseller, J.K.; Steele, J.A.; Schmidt, D.J.; Kim, H.B.; Beamer, G.; Sun, X.; Tzipori, S. Hyperimmune bovine colostrum as a novel therapy to combat Clostridium difficile infection. J. Infect. Dis. 2015, 211, 1334–1341. [Google Scholar] [CrossRef] [Green Version]
- Hutton, M.L.; Cunningham, B.A.; Mackin, K.E.; Lyon, S.A.; James, M.L.; Rood, J.I.; Lyras, D. Bovine antibodies targeting primary and recurrent Clostridium difficile disease are a potent antibiotic alternative. Sci. Rep. 2017, 7, 3665. [Google Scholar] [CrossRef]
- Mattila, E.; Anttila, V.-J.; Broas, M.; Marttila, H.; Poukka, P.; Kuusisto, K.; Pusa, L.; Sammalkorpi, K.; Dabek, J.; Koivurova, O.-P.; et al. A randomized, double-blind study comparing Clostridium difficile immune whey and metronidazole for recurrent Clostridium difficile-associated diarrhoea: Efficacy and safety data of a prematurely interrupted trial. Scand. J. Infect. Dis. 2008, 40, 702–708. [Google Scholar] [CrossRef]
- Numan, S.C.; Veldkamp, P.; Kuijper, E.J.; van den Berg, R.J.; van Dissel, J.T. Clostridium difficile-associated diarrhoea: Bovine anti-Clostridium difficile whey protein to help aid the prevention of relapses. Gut 2007, 56, 888–889. [Google Scholar] [CrossRef] [Green Version]
- Antonopoulos, D.A.; Huse, S.M.; Morrison, H.G.; Schmidt, T.M.; Sogin, M.L.; Young, V.B. Reproducible community dynamics of the gastrointestinal microbiota following antibiotic perturbation. Infect. Immun. 2009, 77, 2367–2375. [Google Scholar] [CrossRef] [Green Version]
- Clavel, T.; Lagkouvardos, I.; Hiergeist, A. Microbiome sequencing: Challenges and opportunities for molecular medicine. Expert Rev. Mol. Diagn. 2016, 16, 795–805. [Google Scholar] [CrossRef]
- Theriot, C.M.; Young, V.B. Microbial and metabolic interactions between the gastrointestinal tract and Clostridium difficile infection. Gut Microbes. 2014, 5, 86–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trzasko, A.; Leeds, J.A.; Praestgaard, J.; Lamarche, M.J.; McKenney, D. Efficacy of LFF571 in a hamster model of Clostridium difficile infection. Antimicrob. Agents Chemother. 2012, 56, 4459–4462. [Google Scholar] [CrossRef] [Green Version]
- Abt, M.C.; McKenney, P.T.; Pamer, E.G. Clostridium difficile colitis: Pathogenesis and host defence. Nat. Rev. Microbiol. 2016, 14, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Loo, V.G.; Bourgault, A.-M.; Poirier, L.; Lamothe, F.; Michaud, S.; Turgeon, N.; Toye, B.; Beaudoin, A.; Frost, E.H.; Gilca, R.; et al. Host and pathogen factors for Clostridium difficile infection and colonization. N. Engl. J. Med. 2011, 365, 1693–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnenburg, J.L.; Xu, J.; Leip, D.D.; Chen, C.-H.; Westover, B.P.; Weatherford, J.; Buhler, J.D.; Gordon, J.I. Glycan foraging in vivo by an intestine-adapted bacterial symbiont. Science 2005, 307, 1955–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.M.W.; de Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Theriot, C.M.; Koenigsknecht, M.J.; Carlson, P.E.; Hatton, G.E.; Nelson, A.M.; Li, B.; Huffnagle, G.B.; Li, J.Z.; Young, V.B. Antibiotic-induced shifts in the mouse gut microbiome and metabolome increase susceptibility to Clostridium difficile infection. Nat. Commun. 2014, 5, 3114. [Google Scholar] [CrossRef]
- Bazanella, M.; Maier, T.V.; Clavel, T.; Lagkouvardos, I.; Lucio, M.; Maldonado-Gòmez, M.X.; Autran, C.; Walter, J.; Bode, L.; Schmitt-Kopplin, P.; et al. Randomized controlled trial on the impact of early-life intervention with bifidobacteria on the healthy infant fecal microbiota and metabolome. Am. J. Clin. Nutr. 2017, 106, 1274–1286. [Google Scholar] [CrossRef] [Green Version]
- Lagkouvardos, I.; Kläring, K.; Heinzmann, S.S.; Platz, S.; Scholz, B.; Engel, K.-H.; Schmitt-Kopplin, P.; Haller, D.; Rohn, S.; Skurk, T.; et al. Gut metabolites and bacterial community networks during a pilot intervention study with flaxseeds in healthy adult men. Mol. Nutr. Food Res. 2015, 59, 1614–1628. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagkouvardos, I.; Joseph, D.; Kapfhammer, M.; Giritli, S.; Horn, M.; Haller, D.; Clavel, T. IMNGS: A comprehensive open resource of processed 16S rRNA microbial profiles for ecology and diversity studies. Sci. Rep. 2016, 6, 33721. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Lagkouvardos, I.; Fischer, S.; Kumar, N.; Clavel, T. Rhea: A transparent and modular R pipeline for microbial profiling based on 16S rRNA gene amplicons. PeerJ 2017, 5, e2836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Bittinger, K.; Charlson, E.S.; Hoffmann, C.; Lewis, J.; Wu, G.D.; Collman, R.G.; Bushman, F.D.; Li, H. Associating microbiome composition with environmental covariates using generalized UniFrac distances. Bioinformatics 2012, 28, 2106–2113. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heidebrecht, H.-J.; Lagkouvardos, I.; Reitmeier, S.; Hengst, C.; Kulozik, U.; Pfaffl, M.W. Alteration of Intestinal Microbiome of Clostridioides difficile-Infected Hamsters during the Treatment with Specific Cow Antibodies. Antibiotics 2021, 10, 724. https://doi.org/10.3390/antibiotics10060724
Heidebrecht H-J, Lagkouvardos I, Reitmeier S, Hengst C, Kulozik U, Pfaffl MW. Alteration of Intestinal Microbiome of Clostridioides difficile-Infected Hamsters during the Treatment with Specific Cow Antibodies. Antibiotics. 2021; 10(6):724. https://doi.org/10.3390/antibiotics10060724
Chicago/Turabian StyleHeidebrecht, Hans-Jürgen, Ilias Lagkouvardos, Sandra Reitmeier, Claudia Hengst, Ulrich Kulozik, and Michael W. Pfaffl. 2021. "Alteration of Intestinal Microbiome of Clostridioides difficile-Infected Hamsters during the Treatment with Specific Cow Antibodies" Antibiotics 10, no. 6: 724. https://doi.org/10.3390/antibiotics10060724
APA StyleHeidebrecht, H.-J., Lagkouvardos, I., Reitmeier, S., Hengst, C., Kulozik, U., & Pfaffl, M. W. (2021). Alteration of Intestinal Microbiome of Clostridioides difficile-Infected Hamsters during the Treatment with Specific Cow Antibodies. Antibiotics, 10(6), 724. https://doi.org/10.3390/antibiotics10060724