
Staphylococcal Biofilm on the Surface of Catheters: Electron Microscopy Evaluation of the Inhibition of Biofilm Growth by RNAIII Inhibiting Peptide


 ,
,  , , and
, , and 
Abstract
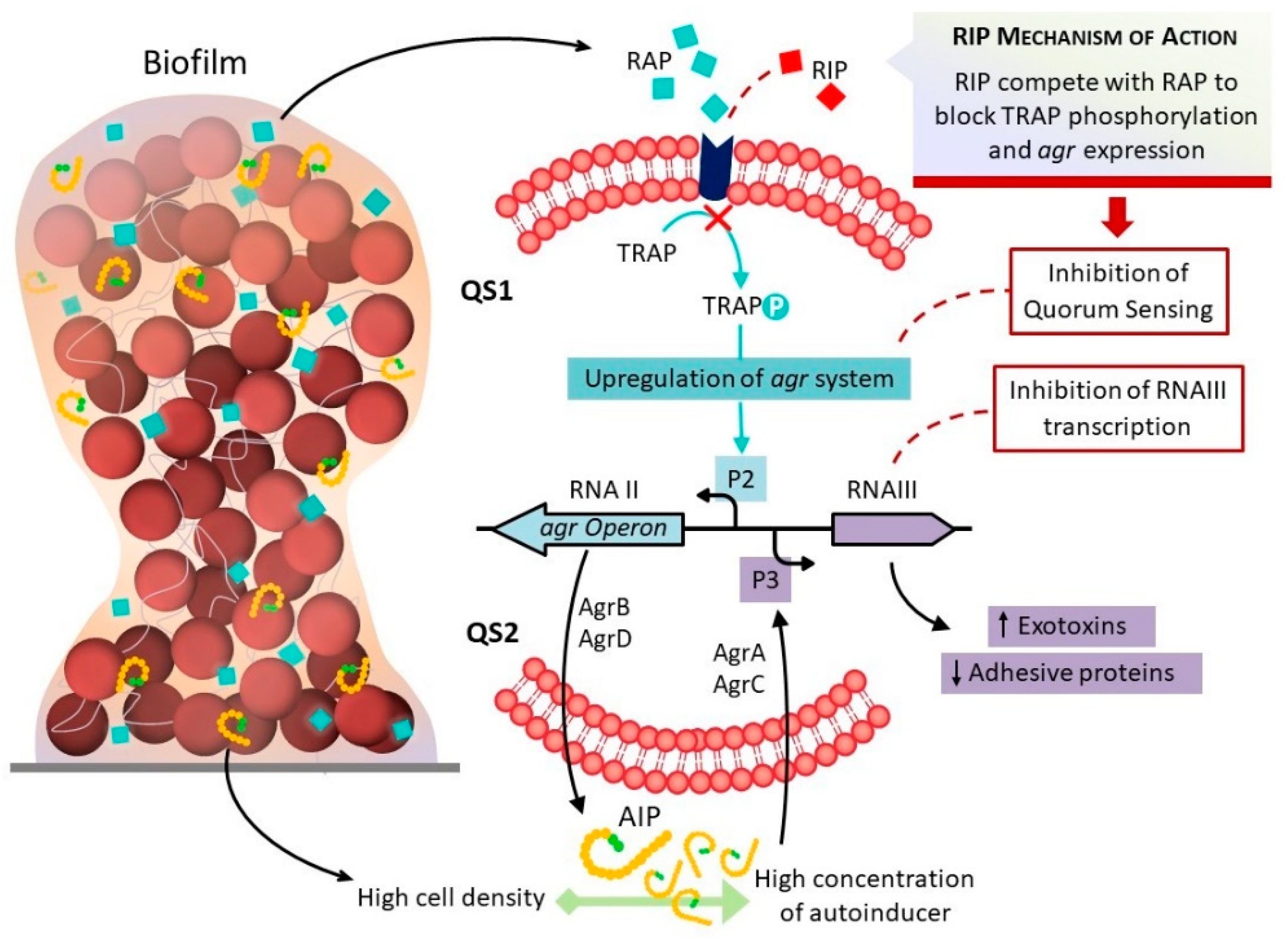
:1. Introduction
2. Results
2.1. Analysis of Biofilm Formation Using the Polystyrene Plate Adherence Method
2.2. Detection of the Presence and Expression of icaADBC Genes by RT-PCR
2.3. Analysis of the Inhibition of Biofilm Formation on Catheter Tips by RIP Using Scanning Electron Microscopy
2.4. Analysis of the Inhibition of Biofilm Formation by RIP Using Polystyrene Plates
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Identification of Microorganisms
4.3. Identification of Staphylococcus
4.4. Analysis of Biofilm Formation by the Polystyrene Plate Adherence Method
4.5. Nucleic Acid Extraction and Amplification (PCR)
4.6. Visualization of the Amplified Products
4.7. RNA Extraction and Gene Expression Analysis of the Ica Operon by RT-PCR
4.8. cDNA Synthesis
4.9. RT-PCR
4.10. Analysis of the Inhibition of Biofilm Formation on Catheter Tips by RIP Using Scanning Electron Microscopy
4.11. Analysis of the Inhibition of Biofilm Formation by RIP Using Polystyrene Plates
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morales, M.; Mendez-Alvarez, S.; Martin-Lopes, J.V.; Marreiro, C.; Freytes, C.O. Biofilm: The microbial “bunker” for intravascular catheter-related infection. Support. Care Cancer 2004, 12, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Nilsdotter-Augustiinsson, A.; Claesson, C.; Lindgren, P.E.; Lundqvist-Gustafsson, H.; Ohman, L. Adherence of Staphylococcus epidermidis to extracellular matrix proteins and effects of fibrinogen-bound bacteria on oxidase activity and apoptosis in neutrophils. APMS 2005, 113, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Gotz, F. Staphylococcus and biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.; Pozzi, C.; Houston, P.; Smyth, D.; Humphreys, H.; Robinson, D.A.; O’Gara, J.P. Association between methicillin susceptibility and biofilm regulation in Staphylococcus aureus isolates from device-related infections. J. Clin. Microbiol. 2007, 45, 1379–1388. [Google Scholar] [CrossRef] [Green Version]
- Donlan, M.R. Biofilm: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–980. [Google Scholar] [CrossRef]
- Cafiso, V.; Bertuccio, T.; Santagati, M.; Campanile, F.; Amicosante, G.; Perilli, M.G.; Selan, L.; Artini, M.; Nicoletti, G.; Stefani, S. Presence of the ica operon in clinical isolates of Staphylococcus epidermidis and its role in biofilm production. Clin. Microbiol. Infect. 2004, 10, 1081–1088. [Google Scholar] [CrossRef] [Green Version]
- Mack, D.; Fischer, W.; Krokotsch, A.; Leopold, K.; Hartmann, R.; Egge, H.; Laufs, R. The intercellular adhesin involved in biofilm accumulation of Staphylococcus epidermidis is a linear b-1,6-linked glucosaminoglycan: Purification and structural analysis. J. Bacteriol. 1996, 178, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maira-Litran, T.; Kropec, A.; Abeygunawardana, C.; Joyce, J.; Mark, G., III; Goldmann, D.A.; Pier, G.B. Immunochemical properties of the staphylococcal poly-N-acetylglucosamine surface polysaccharide. Infect. Immun. 2002, 70, 4433–4440. [Google Scholar] [CrossRef] [Green Version]
- Heilmann, C.; Schweitzer, O.; Gerke, C.; Vanittanakom, N.; Mack, D.; Götz, F. Molecular basis of intercellular adhesion in the biofilm-forming Staphylococcus epidermidis. Mol. Microbiol. 1996, 20, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Cucarella, C.; Solano, C.; Valle, J.; Amorena, B.; Lasa, I.; Penadés, J.R. Bap, a Staphylococcus aureus surface protein involved in biofilm formation. J. Bacteriol. 2001, 183, 2888–2896. [Google Scholar] [CrossRef] [Green Version]
- Vadyvaloo, V.; Otto, M. Molecular genetics of Staphylococcus epidermidis on indwelling medical devices. Int. J. Artif. Organs 2005, 28, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Cunha, M.L.R.S. Staphylococcus aureus and Coagulase-negative staphylococci. In Virulence, Antimicrobial Resistance and Molecular Epidemiology; Nova Science Publishers: New York, NY, USA, 2014. [Google Scholar]
- Balaban, N.; Giacometti, A.; Cirioni, O.; Gov, Y.; Ghiselli, R.; Mocchegiani, F.; Viticchi, C.; Del Prete, M.S.; Saba, V.; Scalise, G.; et al. Use the quorum-sensing inhibitor RNAIII-inhibiting peptide to prevent biofilm formation in vivo by drug-resistant Staphylococcus epidermidis. J. Infect. Dis. 2003, 187, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Balaban, N.; Cirioni, O.; Giacometti, A.; Ghiselli, R.; Braunstein, J.B.; Silvestri, C.; Mocchegiani, F.; Saba, V.; Scalise, G. Treatment of Staphylococcus aureus biofilm infection by the quorum sensing inhibitor RIP. Antimicrob. Agents Chemother. 2007, 51, 2226–2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonetti, O.; Cirioni, O.; Ghiselli, R.; Goteri, G.; Scalise, A.; Orlando, F.; Silvestri, C.; Riva, A.; Saba, V.; Madanahally, K.D.; et al. RNAIII-inhibiting peptide enhances healing of wounds infected with methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2008, 52, 2205–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, L.N.; Jonsson, I.M.; Singh, V.K.; Tarkowski, A.; Stewart, G.S. Inactivation of traP has no effect on the agr quorum-sensing system or virulence of Staphylococcus aureus. Infect. Immun. 2007, 75, 4519–4527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, R.P.; Arvidson, S.; Novick, P. A Nonsense mutation in agrA accounts for the defect in agr expression and the avirulence of Staphylococcus aureus 8325-4 traP:kan. Infect. Immun. 2007, 75, 4534–4540. [Google Scholar] [CrossRef] [Green Version]
- Balaban, N.; Goldkorn, T.; Gov, Y.; Hirshberg, M.; Koyfman, N.; Mathews, H.R.; Nhan, R.T.; Singh, B.; Uziel, O. Regulation of Staphylococcus aureus pathogenesis via target of RNAIII-activating protein (TRAP). J. Biol. Chem. 2001, 276, 2658–2667. [Google Scholar] [CrossRef] [Green Version]
- Korem, M.; Gov, Y.; Madanahally, D.K.; Balaban, N. Transcriptional profiling (microarray analysis) of TRAP, a master regulator of staphylococcal virulence. Infect. Immun. 2005, 73, 6220–6228. [Google Scholar] [CrossRef] [Green Version]
- Balaban, N.; Stoodley, P.; Fux, C.A.; Wilson, S.; Costerton, J.W.; Dell’Acqua, G. Prevention of staphylococcal biofilm-associated infections by the quorum sensing inhibitor RIP. Clin. Orthop. Relat. Res. 2005, 437, 48–54. [Google Scholar] [CrossRef]
- Marrie, T.J.; Costerton, J.W. Scanning and transmission electron microscopy of in situ bacterial colonization of intravenous and intraarterial catheters. J. Clin. Microbiol. 1984, 19, 687–693. [Google Scholar] [CrossRef] [Green Version]
- Fredheim, E.G.A.; Klingenberg, C.; Rohde, H.; Frankenberger, S.; Gaustad, P.; Flægstad, T.; Sollid, J.E. Biofilm formation by Staphylococcus haemolyticus. J. Clin. Microbiol. 2009, 47, 1172–1180. [Google Scholar] [CrossRef] [Green Version]
- Kini, G.D.; Patel, K.; Parris, A.R.; Tang, J.S. Unusual presentation of endocarditis caused by Staphylococcus warneri. Open Microbiol. J. 2010, 4, 103–105. [Google Scholar] [CrossRef] [Green Version]
- Frank, K.L.; Patel, R. Poly-N-acetylglucosamine is not a major component of the extracellular matrix in biofilms formed by icaADBC-positive Staphylococcus lugdunensis isolates. Infect. Immun. 2007, 75, 4728–4742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.L.; Zhang, S.K.; Chen, X.X.; Chen, J.Y.; Zhong, B. Growth properties of Staphylococcus aureus in biofilm formed on polystyrene plate. Afr. J. Microbiol. Res. 2012, 6, 3284–3291. [Google Scholar]
- Koneman, E.W.; Allen, S.D.; Janda, W.M.; Winn, W.C.; Schreckenberger, P.C. Koneman’s Color Atlas and Textbook of Diagnostic Microbiology, 6th ed.; JB Lippincott: Philadelphia, PA, USA, 1997. [Google Scholar]
- Couto, I.; Pereira, S.; Miragaia, M.; Sanches, I.S.; Lencastre, H. Identification of clinical staphylococcal isolates from humans by Internal Transcribed Spacer PCR. J. Clin. Microbiol. 2001, 39, 3099–3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, G.D.; Simpsonv, W.A.; Yonger, J.J.; Baddor, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative Staphylococci to plastic tissue culture plates: A quantitative model for the adherence of Staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, A.; Cunha, M.L.R.S. Comparison of methods for the detection of biofilm production in coagulase-negative staphylococci. BMC Res. Notes 2010, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Arciola, C.R.; Baldassarri, L.; Montanaro, L. Presence of icaA and icaD genes and slime production in a collection of staphylococcal strains from catheter-associated infections. J. Clin. Microbiol. 2001, 39, 2151–2156. [Google Scholar] [CrossRef] [Green Version]
- Arciola, C.R.; Gamberini, S.; Campoccia, D.; Visai, L.; Speziale, P.; Baldassari, L.; Montanaro, L. A multiplex PCR method for the detection of all five individual genes of ica locus in Staphylococcus epidermidis. A survey on 400 clinical isolates from prosthesis-associated infections. J. Biomed. Mater. Res. A 2005, 75A, 408–413. [Google Scholar] [CrossRef]
- Rohde, H.; Burandt, E.C.; Sienssem, N.; Frommelt, L.; Burdesk, C.; Wurster, S.; Scherp, S.; Davies, A.P.; Harris, L.G.; Horstikotte, M.A.; et al. Polysaccharide intercellular adhesin or protein factors in biofilm accumulation of Staphylococcus epidermidis and Staphylococcus aureus isolated from prosthetic hip and knee joint infections. Biomaterials 2007, 28, 1711–1720. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Plate Adherence Test | ||
|---|---|---|---|
| SA | WA | NA | |
| S. aureus, % (n) | 12.5 (1) | 31.7 (13) | 23.8 (36) |
| S. epidermidis, % (n) | 50.0 (4) | 22.0 (9) | 24.5 (37) |
| S. saprophyticus, % (n) | 12.5 (1) | 29.3 (12) | 4.6 (7) |
| S. haemolyticus, % (n) | 12.5 (1) | 4.9 (2) | 11.3 (17) |
| S. hominis, % (n) | - | 2.4 (1) | 12.6 (19) |
| S. warneri, % (n) | - | 7.3 (3) | 11.3 (17) |
| S. lugdunensis, % (n) | 12.5 (1) | 2.4 (1) | 11.9 (18) |
| Total number of isolates, % (n) | 100 (8) | 100 (41) | 100 (151) |
| Species | PCR | RT-PCR | Plate Adherence Test |
|---|---|---|---|
| S. aureus | ADBC | ADBC | SA |
| S. epidermidis | ADBC | ADBC | SA |
| S. epidermidis | ADBC | DBC | SA |
| S. saprophyticus | ADBC | DBC | SA |
| S. haemolyticus | ADBC | A | SA |
| S. haemolyticus | CD | C | WA |
| S. hominis | ADC | - | WA |
| S. warneri | DC | - | WA |
| S. lugdunensis | AD | AD | SA |
| S. lugdunensis | A | - | WA |
| Strains | Plate Adherence Test | OD TSB Alone + 2% Glucose | OD TSB + 2% Glucose with 200 µg/mL RIP | Biofilm Inhibition (%) |
|---|---|---|---|---|
| S. saprophyticus ACDB | SA | 0.315 | 0.109 | 65.4 |
| S. epidermidis ACDB | SA | 0.319 | 0.110 | 65.5 |
| S. epidermidis CDB | SA | 0.236 | 0.055 | 76.7 |
| S. haemolyticus A | SA | 0.261 | 0.074 | 71.6 |
| S. hominis | WA | 0.118 | 0.077 | 34.7 |
| S. warneri | WA | 0.176 | 0.049 | 72.1 |
| S. lugdunensis AD | SA | 0.287 | 0.042 | 85.4 |
| S. aureus ACDB | SA | 0.441 | 0.398 | 9.7 |
| S. epidermidis ATCC 35983 | WA | 0.193 | 0.037 | 80.8 |
| S. epidermidis ATCC 12228 | NA | 0.077 | 0.055 | 28.6 |
| Primer | 5′ to 3′ Nucleotide Sequence | Amplified Product (bp) | Reference |
|---|---|---|---|
| icaAF | TCT CTT GCA GGA GCA ATC AA | 187 | [30] |
| icaAR | TCA GGC ACT AAC ATC CAG CA | ||
| icaAF | TGG CTG TAT TAA GCG AAG TC | 669 | [32] |
| icaAR | CCT CTG TCT GGG CTT GAC C | ||
| icaBF | CTG ATC AAG AAT TTA AAT CAC AAA | 302 | [31] |
| icaBR | AAA GTC CCA TAA GCC TGT TT | ||
| icaCF | TAA CTT TAG GCG CAT ATG TTT | 400 | [31] |
| icaCR | TTC CAG TTA GGC TGG TAT TG | ||
| icaDF | ATG GTC AAG CCC AGA CAG AG | 198 | [30] |
| icaDR | CGT GTT TTC AAC ATT TAA TGC AA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, A.; Pinheiro-Hubinger, L.; Pereira, V.C.; Riboli, D.F.M.; Martins, K.B.; Romero, L.C.; Cunha, M.d.L.R.d.S.d. Staphylococcal Biofilm on the Surface of Catheters: Electron Microscopy Evaluation of the Inhibition of Biofilm Growth by RNAIII Inhibiting Peptide. Antibiotics 2021, 10, 879. https://doi.org/10.3390/antibiotics10070879
de Oliveira A, Pinheiro-Hubinger L, Pereira VC, Riboli DFM, Martins KB, Romero LC, Cunha MdLRdSd. Staphylococcal Biofilm on the Surface of Catheters: Electron Microscopy Evaluation of the Inhibition of Biofilm Growth by RNAIII Inhibiting Peptide. Antibiotics. 2021; 10(7):879. https://doi.org/10.3390/antibiotics10070879
Chicago/Turabian Stylede Oliveira, Adilson, Luiza Pinheiro-Hubinger, Valéria Cataneli Pereira, Danilo Flávio Moraes Riboli, Katheryne Benini Martins, Letícia Calixto Romero, and Maria de Lourdes Ribeiro de Souza da Cunha. 2021. "Staphylococcal Biofilm on the Surface of Catheters: Electron Microscopy Evaluation of the Inhibition of Biofilm Growth by RNAIII Inhibiting Peptide" Antibiotics 10, no. 7: 879. https://doi.org/10.3390/antibiotics10070879
APA Stylede Oliveira, A., Pinheiro-Hubinger, L., Pereira, V. C., Riboli, D. F. M., Martins, K. B., Romero, L. C., & Cunha, M. d. L. R. d. S. d. (2021). Staphylococcal Biofilm on the Surface of Catheters: Electron Microscopy Evaluation of the Inhibition of Biofilm Growth by RNAIII Inhibiting Peptide. Antibiotics, 10(7), 879. https://doi.org/10.3390/antibiotics10070879