
Characterization and Clonal Diffusion of Ceftaroline Non-Susceptible MRSA in Two Hospitals in Central Italy

 , , ,
, , , 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Susceptibility to Ceftaroline
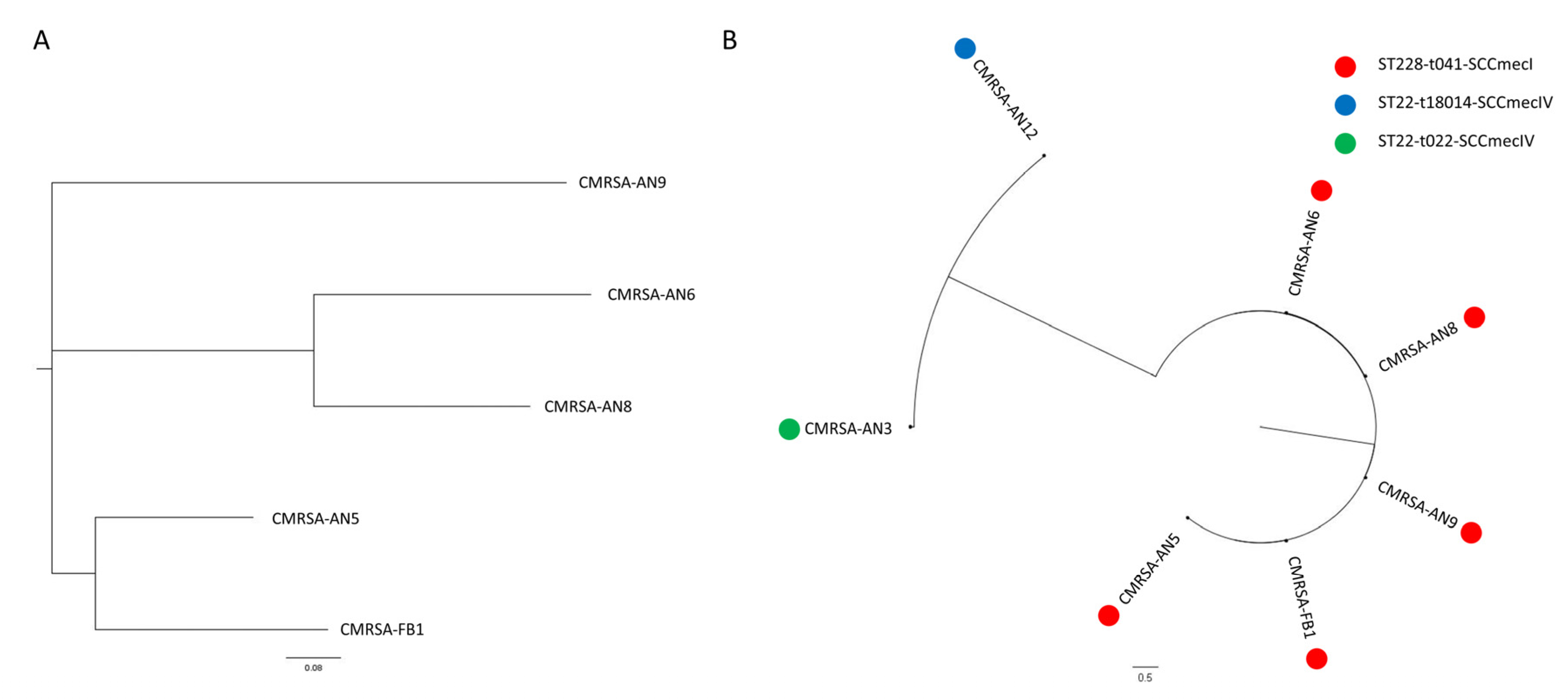
2.2. Epidemiology of Ceftaroline Non-Susceptible MRSA
2.3. Analysis of PBP and GDPP Mutations
2.4. Resistome and Virulence Analysis
3. Materials and Methods
3.1. Strain
3.2. Susceptibility to Ceftaroline and MIC Determination
3.3. Typing, WGS, and Genome Analysis
3.4. Analysis of PBP Mutations
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati., M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef]
- Otto, M. MRSA virulence and spread. Cell Microbiol. 2012, 14, 1513–1521. [Google Scholar] [CrossRef] [Green Version]
- Chambers, H.F.; Deleo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, O.; Lucarini, G.; Orlando, F.; Pierpaoli, E.; Ghiselli, R.; Provinciali, M.; Castelli, P.; Guerrieri, M.; Di Primio, R.; Offidani, A.; et al. Role of Daptomycin on Burn Wound Healing in an Animal Methicillin-Resistant Staphylococcus aureus Infection Model. Antimicrob. Agents Chemother. 2017, 24, e00606-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonetti, O.; Lucarini, G.; Morroni, G.; Orlando, F.; Lazzarini, R.; Zizzi, A.; Brescini, L.; Provinciali, M.; Giacometti, A.; Offidani, A.; et al. New Evidence and Insights on Dalbavancin and Wound Healing in a Mouse Model of Skin Infection. Antimicrob. Agents Chemother. 2020, 24, e02062-19. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, O.; Rizzetto, G.; Molinelli, E.; Cirioni, O.; Offidani, A. Review: A Safety Profile of Dalbavancin for On- and Off-Label Utilization. Ther. Clin. Risk Manag. 2021, 22, 223–232. [Google Scholar] [CrossRef]
- Silvestri, C.; Cirioni, O.; Arzeni, D.; Ghiselli, R.; Simonetti, O.; Orlando, F.; Ganzetti, G.; Staffolani, S.; Brescini, L.; Provinciali, M.; et al. In vitro activity and in vivo efficacy of tigecycline alone and in combination with daptomycin and rifampin against Gram-positive cocci isolated from surgical wound infection. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1759–1764. [Google Scholar] [CrossRef]
- David, M.Z.; Dryden, M.; Gottlieb, T.; Tattevin, P.; Gould, I.M. Recently approved antibacterials for methicillin-resistant Staphylococcus aureus (MRSA) and other Gram-positive pathogens: The shock of the new. Int. J. Antimicrob. Agents 2017, 50, 303–307. [Google Scholar] [CrossRef]
- Abbas, M.; Paul, M.; Huttner, A. New and improved? A review of novel antibiotics for Gram-positive bacteria. Clin. Microbiol. Infect. 2017, 23, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Laudano, J.B. Ceftaroline fosamil: A new broad-spectrum cephalosporin. J. Antimicrob. Chemother. 2011, 66 (Suppl. 3), iii11–iii18. [Google Scholar] [CrossRef]
- Sader, H.S.; Flamm, R.K.; Streit, J.M.; Carvalhaes, C.G.; Mendes, R.E. Antimicrobial activity of ceftaroline and comparator agents tested against organisms isolated from patients with community-acquired bacterial pneumonia in Europe, Asia, and Latin America. Int. J. Infect. Dis. 2018, 77, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Bae, I.G.; Stone, G.G. Activity of ceftaroline against pathogens associated with community-acquired pneumonia collected as part of the AWARE surveillance program, 2015–2016. Diagn Microbiol. Infect. Dis. 2019, 95, 114843. [Google Scholar] [CrossRef] [PubMed]
- Horner, C.; Mushtaq, S.; Livermore, D.M. BSAC Resistance Surveillance Standing Committee. Activity of ceftaroline versus ceftobiprole against staphylococci and pneumococci in the UK and Ireland: Analysis of BSAC surveillance data. J. Antimicrob. Chemother. 2020, 75, 3239–3243. [Google Scholar] [PubMed]
- Lahiri, S.D.; McLaughlin, R.E.; Whiteaker, J.D.; Ambler, J.E.; Alm, R.A. Molecular characterization of MRSA isolates bracketing the current EUCAST ceftaroline-susceptible breakpoint for Staphylococcus aureus: The role of PBP2a in the activity of ceftaroline. J. Antimicrob. Chemother. 2015, 70, 2488–2498. [Google Scholar] [CrossRef] [Green Version]
- Kelley, W.L.; Jousselin, A.; Barras, C.; Lelong, E.; Renzoni, A. Missense mutations in PBP2A Affecting ceftaroline susceptibility detected in epidemic hospital-acquired methicillin-resistant Staphylococcus aureus clonotypes ST228 and ST247 in Western Switzerland archived since 1998. Antimicrob. Agents Chemother. 2015, 59, 1922–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greninger, A.L.; Chatterjee, S.S.; Chan, L.C.; Hamilton, S.M.; Chambers, H.F.; Chiu, C.Y. Whole-Genome Sequencing of Methicillin-Resistant Staphylococcus aureus Resistant to Fifth-Generation Cephalosporins Reveals Potential Non-mecA Mechanisms of Resistance. PLoS ONE 2016, 18, e0149541. [Google Scholar] [CrossRef]
- Bongiorno, D.; Mongelli, G.; Stefania, S.; Campanile, F. Genotypic analysis of Italian MRSA strains exhibiting low-level ceftaroline and ceftobiprole resistance. Diagn Microbiol. Infect. Dis. 2019, 95, 114852, Erratum in Diagn. Microbiol. Infect. Dis. 2020, 96, 115000. [Google Scholar]
- Sader, H.S.; Rhomberg, P.R.; Doyle, T.B.; Flamm, R.K.; Mendes, R.E. Evaluation of the Revised Ceftaroline Disk Diffusion Breakpoints When Testing a Challenge Collection of Methicillin-Resistant Staphylococcus aureus Isolates. J. Clin. Microbiol. 2018, 27, e00777-18. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Rivas, L.M.; Spencer, M.; Martinez, R.; Lam, M.; Rojas, P.; Porte, L.; Silva, F.; Braun, S.; Valdivieso, F.; et al. A Multicenter Study to Evaluate Ceftaroline Breakpoints: Performance in an Area with High Prevalence of Methicillin-Resistant Staphylococcus aureus Sequence Type 5 Lineage. J. Clin. Microbiol. 2019, 57, e00798-19, Erratum in J. Clin. Microbiol. 2021, 19, 59. [Google Scholar] [CrossRef] [Green Version]
- Soler-Gómez, A.; Arca-Suárez, J.; González-Gómez, M.C.; Galán-Sánchez, F.; Rodríguez-Iglesias, M. Antimicrobial activity of ceftaroline against methicillin-resistant Staphylococcus aureus (MRSA) isolates from Spain and molecular characterisation of borderline susceptible and resistant strains. J. Glob. Antimicrob. Resist. 2018, 15, 171–172. [Google Scholar] [CrossRef]
- Sader, H.S.; Jones, R.N. Impact of EUCAST, CLSI and USCAST ceftaroline breakpoint changes on the susceptibility of methicillin-resistant Staphylococcus aureus isolates collected from US medical centres (2015–2018). Clin. Microbiol. Infect. 2020, 26, 658–659. [Google Scholar] [CrossRef]
- Urbán, E.; Stone, G.G. Impact of EUCAST ceftaroline breakpoint change on the susceptibility of methicillin-resistant Staphylococcus aureus isolates collected from patients with complicated skin and soft-tissue infections. Clin. Microbiol. Infect. 2019, 25, 1429.e1–1429.e4. [Google Scholar] [CrossRef] [Green Version]
- Morroni, G.; Brenciani, A.; Vincenzi, C.; Barocci, S.; Tili, E.; Manso, E.; Mingoia, M.; Menzo, S.; Varaldo, P.E.; Giovanetti, E. A clone of linezolid-resistant Staphylococcus epidermidis bearing the G2576T mutation is endemic in an Italian hospital. J. Hosp. Infect. 2016, 94, 203–206. [Google Scholar] [CrossRef]
- Alm, R.A.; McLaughlin, R.E.; Kos, V.N.; Sader, H.S.; Iaconis, J.P.; Lahiri, S.D. Analysis of Staphylococcus aureus clinical isolates with reduced susceptibility to ceftaroline: An epidemiological and structural perspective. J. Antimicrob. Chemother. 2014, 69, 2065–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argudín, M.A.; Dodémont, M.; Taguemount, M.; Roisin, S.; de Mendonça, R.; Deplano, A.; Nonhoff, C.; Denis, O. In vitro activity of ceftaroline against clinical Staphylococcus aureus isolates collected during a national survey conducted in Belgian hospitals. J. Antimicrob. Chemother. 2017, 72, 56–59, Erratum in J. Antimicrob. Chemother. 2018, 1, 2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonelli, A.; Giani, T.; Coppi, M.; Di Pilato, V.; Arena, F.; Colavecchio, O.L.; Conte, V.; Santerre Henriksen, A.; Rossolini, G.M. MRSA-HAP Study Group. Staphylococcus aureus from hospital-acquired pneumonia from an Italian nationwide survey: Activity of ceftobiprole and other anti-staphylococcal agents, and molecular epidemiology of methicillin-resistant isolates. J. Antimicrob. Chemother. 2019, 1, 3453–3461. [Google Scholar] [CrossRef] [PubMed]
- Morroni, G.; Brenciani, A.; Brescini, L.; Fioriti, S.; Simoni, S.; Pocognoli, A.; Mingoia, M.; Giovanetti, E.; Barchiesi, F.; Giacometti, A.; et al. High Rate of Ceftobiprole Resistance among Clinical Methicillin-Resistant Staphylococcus aureus Isolates from a Hospital in Central Italy. Antimicrob. Agents Chemother. 2018, 26, e01663-18. [Google Scholar] [CrossRef] [Green Version]
- Ba, X.; Kalmar., L.; Hadjirin, N.F.; Kerschner, H.; Apfalter, P.; Morgan, F.J.; Paterson, G.K.; Girvan, S.L.; Zhou, R.; Harrison, E.M.; et al. Truncation of GdpP mediates β-lactam resistance in clinical isolates of Staphylococcus aureus. J. Antimicrob. Chemother. 2019, 1, 1182–1191. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, J.M.; O’Neill, A.J. Loss of function of the gdpP protein leads to joint β-lactam/glycopeptide tolerance in Staphylococcus aureus. Antimicrob. Agents Chemother. 2012, 56, 579–581. [Google Scholar] [CrossRef] [Green Version]
- Schaumburg, F.; Peters, G.; Alabi, A.; Becker, K.; Idelevich, E.A. Missense mutations of PBP2a are associated with reduced susceptibility to ceftaroline and ceftobiprole in African MRSA. J. Antimicrob. Chemother. 2016, 71, 41–44. [Google Scholar] [CrossRef]
- Paulsen, I.T.; Brown, M.H.; Littlejohn, T.G.; Mitchell, B.A.; Skurray, R.A. Multidrug resistance proteins QacA and QacB from Staphylococcus aureus: Membrane topology and identification of residues involved in substrate specificity. Proc. Natl. Acad. Sci. USA 1996, 16, 3630–3635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gravet, A.; Colin, D.A.; Keller, D.; Girardot, R.; Monteil, H.; Prévost, G. Characterization of a novel structural member, LukE-LukD, of the bi-component staphylococcal leucotoxins family. FEBS Lett. 1998, 436, 202–208, Erratum in FEBS Lett. 1998, 18, 342. [Google Scholar] [CrossRef] [Green Version]
- Baldan, R.; Rancoita, P.M.; Di Serio, C.; Mazzotti, M.; Cichero, P.; Ossi, C.; Biancardi, A.; Nizzero, P.; Saracco, A.; Scarpellini, P.; et al. Epidemic MRSA clone ST22-IV is more resistant to multiple host- and environment-related stresses compared with ST228-I. J. Antimicrob. Chemother. 2015, 70, 757–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 11.0. 2021. Available online: http://www.eucast.org (accessed on 1 February 2021).
- CLSI. Performance Strandards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplemente M100; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Brenciani, A.; Morroni, G.; Pollini, S.; Tiberi, E.; Mingoia, M.; Varaldo, P.E.; Rossolini, G.M.; Giovanetti, E. Characterization of novel conjugative multiresistance plasmids carrying cfr from linezolid-resistant Staphylococcus epidermidis clinical isolates from Italy. J. Antimicrob. Chemother. 2016, 71, 307–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 24, 124. [Google Scholar] [CrossRef]


{kind=link}
| Isolate Category | Breakpoint Diameters (mm) | No. of Isolates Tested (% on Total Isolates of the Study) | ||||
|---|---|---|---|---|---|---|
| EUCAST | CLSI | DD a (EUCAST) | DD a (CLSI) | MIC b (EUCAST) | MIC b (CLSI) | |
| Susceptible | ≥20 | ≥25 | 186 (77.8%) | 228 (95.4%) | 39 (94.1%) | 39 (94.1%) |
| Susceptible, increased exposure | 17–19 | NA | 41 (17.2%) | NA | 7 (2.9%) | NA |
| Susceptible-dose dependent | NA | 20–24 | NA | 10 (4.2%) | NA | 14 (5.8%) |
| Resistant | <17 | ≤19 | 12 (5.0%) | 1 (0.4%) | 7 (2.9%) | 0 (0%) |
| Strain | MIC (mg/L) | Amino Acid Substitutions | PFGE Pulsotype | Spa Type | MLST | SCCmec | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PBP2a | PBP1 | PBP2 | PBP3 | PBP4 | GDPP | ||||||
| CMRSA-AN3 | 2 | S225R | S629-S664T | C197Y-L246V-P285A-T439V-T691A | R504K-K584N | D98E-S189T-E398A | I52V-N105D-S391P | C | t18014 | ST22 | IV |
| CMRSA-AN5 | 2 | N146K | WT | C197Y | T92X | WT | WT | A | t041 | ST228 | I |
| CMRSA-AN6 | 4 | N146K | S194N | C197Y | WT | N337D | WT | D | t041 | ST228 | I |
| CMRSA-AN8 | 2 | N146K | WT | C197Y | WT | L354X | Truncated | E | t041 | ST228 | I |
| CMRSA-AN9 | 4 | N146K | WT | WT | WT | WT | WT | F | t041 | ST228 | I |
| CMRSA-AN12 | 2 | WT | S629T-S664T | C197Y-L246T-P285A-T439V-T691A | R504K-K584N | D98E-S189T-E398A | I52V-N105D-S391P | G | t022 | ST22 | IV |
| CMRSA-FB1 | 2 | N146K | WT | WT | WT | WT | WT | B | t041 | ST228 | I |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morroni, G.; Fioriti, S.; Salari, F.; Brenciani, A.; Brescini, L.; Mingoia, M.; Giovanetti, E.; Pocognoli, A.; Giacometti, A.; Molinelli, E.; et al. Characterization and Clonal Diffusion of Ceftaroline Non-Susceptible MRSA in Two Hospitals in Central Italy. Antibiotics 2021, 10, 1026. https://doi.org/10.3390/antibiotics10081026
Morroni G, Fioriti S, Salari F, Brenciani A, Brescini L, Mingoia M, Giovanetti E, Pocognoli A, Giacometti A, Molinelli E, et al. Characterization and Clonal Diffusion of Ceftaroline Non-Susceptible MRSA in Two Hospitals in Central Italy. Antibiotics. 2021; 10(8):1026. https://doi.org/10.3390/antibiotics10081026
Chicago/Turabian StyleMorroni, Gianluca, Simona Fioriti, Federica Salari, Andrea Brenciani, Lucia Brescini, Marina Mingoia, Eleonora Giovanetti, Antonella Pocognoli, Andrea Giacometti, Elisa Molinelli, and et al. 2021. "Characterization and Clonal Diffusion of Ceftaroline Non-Susceptible MRSA in Two Hospitals in Central Italy" Antibiotics 10, no. 8: 1026. https://doi.org/10.3390/antibiotics10081026
APA StyleMorroni, G., Fioriti, S., Salari, F., Brenciani, A., Brescini, L., Mingoia, M., Giovanetti, E., Pocognoli, A., Giacometti, A., Molinelli, E., Offidani, A., Simonetti, O., & Cirioni, O. (2021). Characterization and Clonal Diffusion of Ceftaroline Non-Susceptible MRSA in Two Hospitals in Central Italy. Antibiotics, 10(8), 1026. https://doi.org/10.3390/antibiotics10081026