
Impact of Chronic Tetracycline Exposure on Human Intestinal Microbiota in a Continuous Flow Bioreactor Model

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Structural Change of the Microbial Community
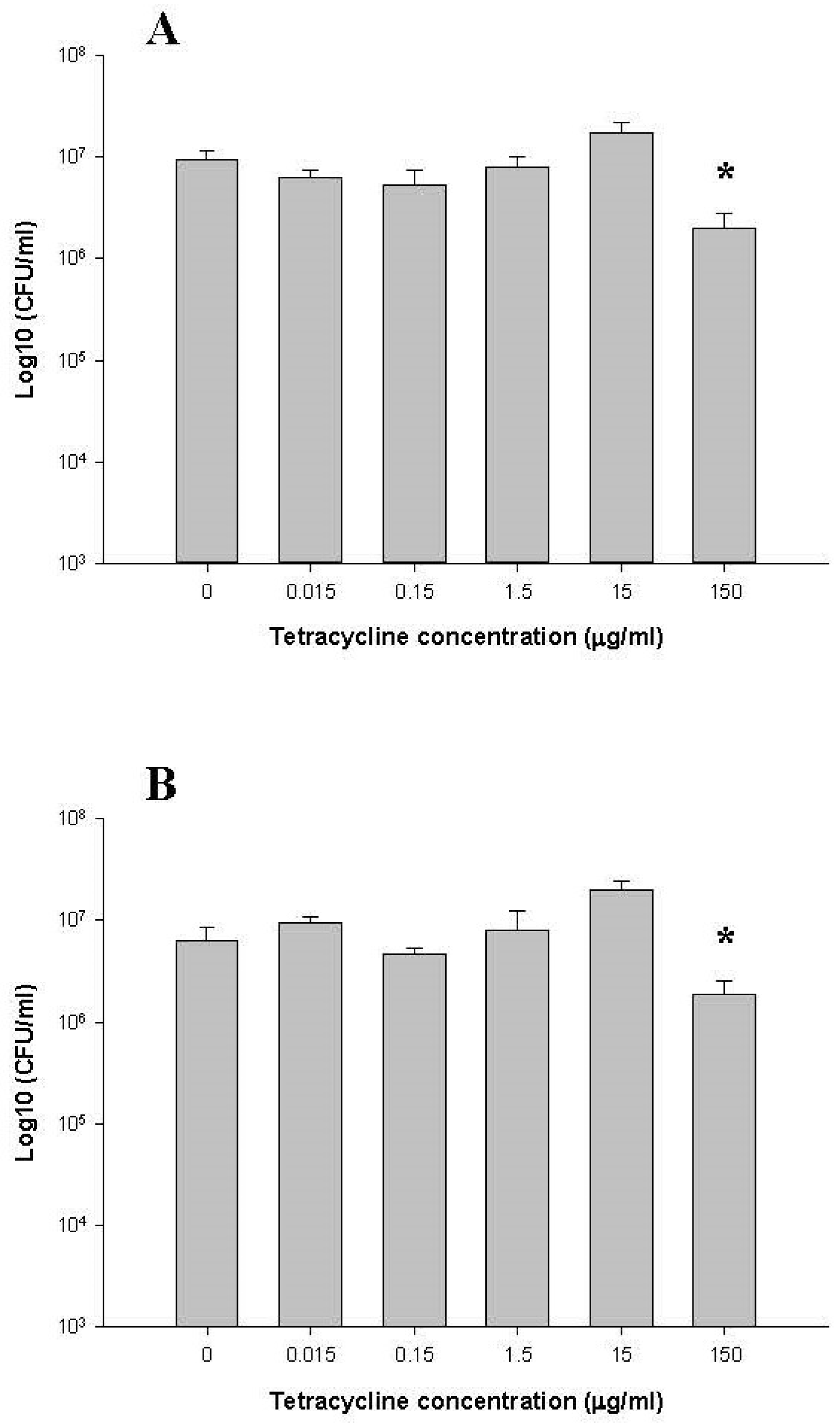
2.1.1. Total Counts of Anaerobic Bacteria
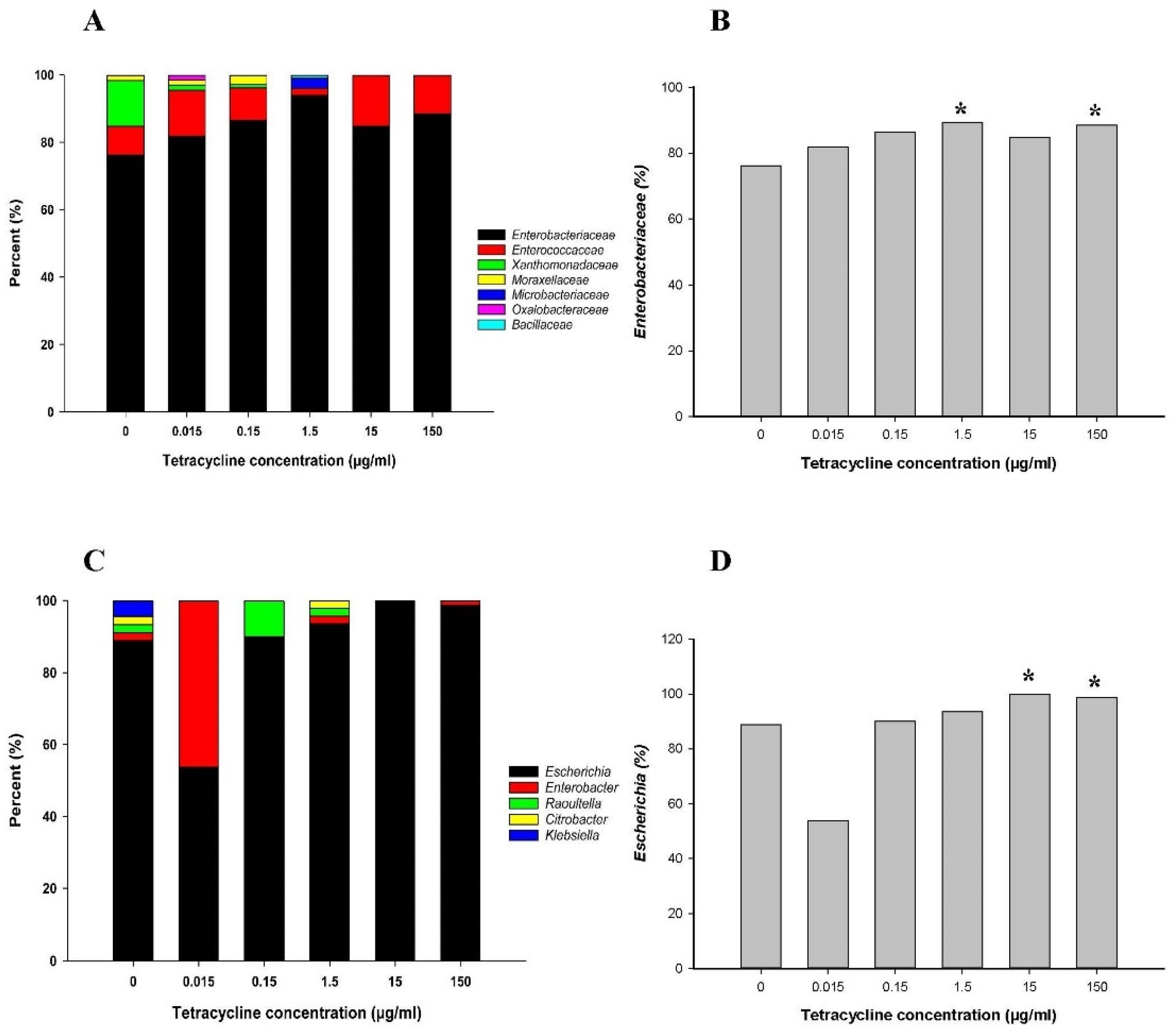
2.1.2. 16S Metagenomic Sequencing Analysis
2.1.3. Abundance of the Family Enterobacteriaceae and the Genus Escherichia
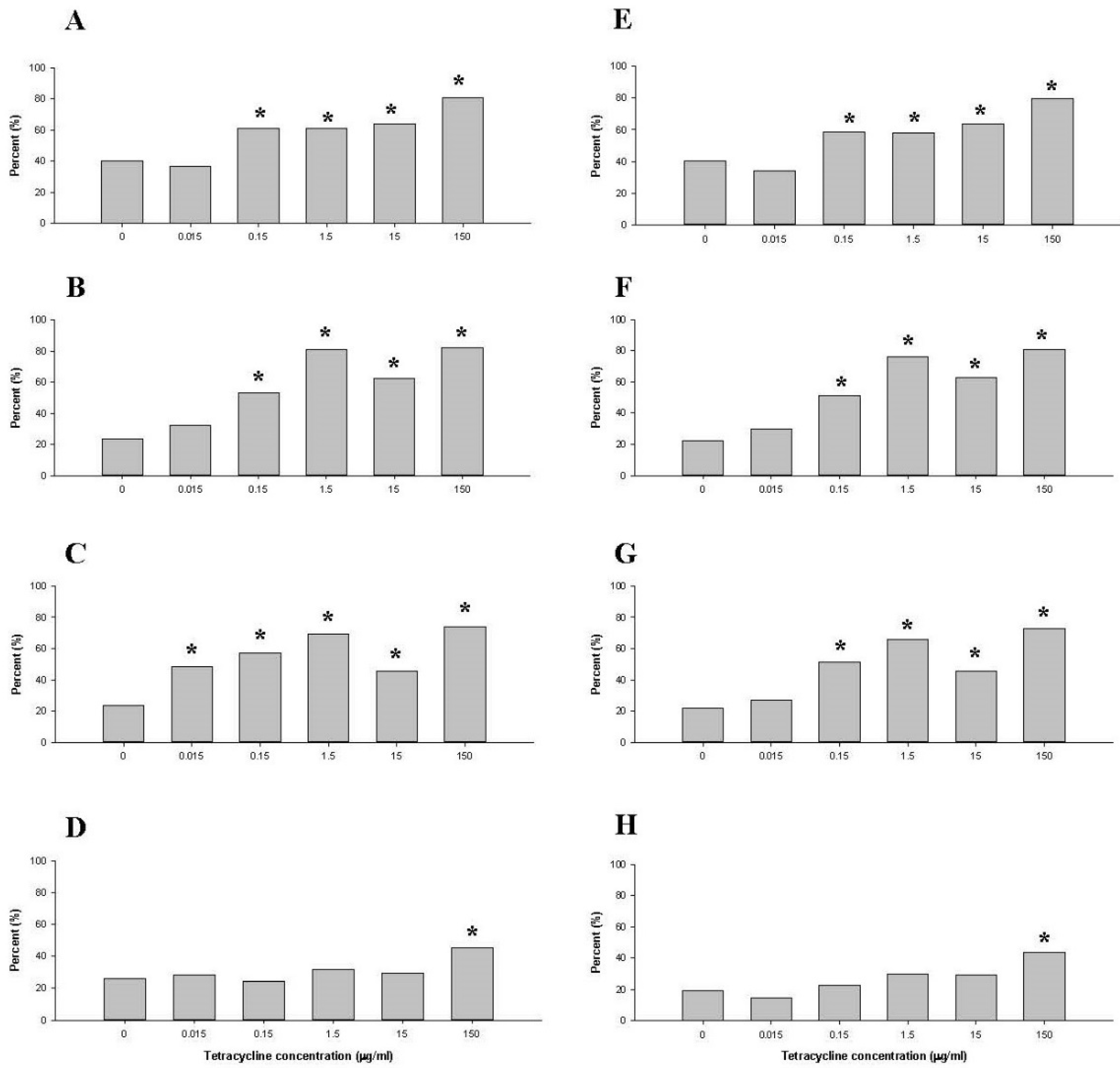
2.1.4. Comparison of Tetracycline Resistance Genes in the Family Enterobacteriaceae and the Genus Escherichia
2.2. Functional Change of the Microbial Community
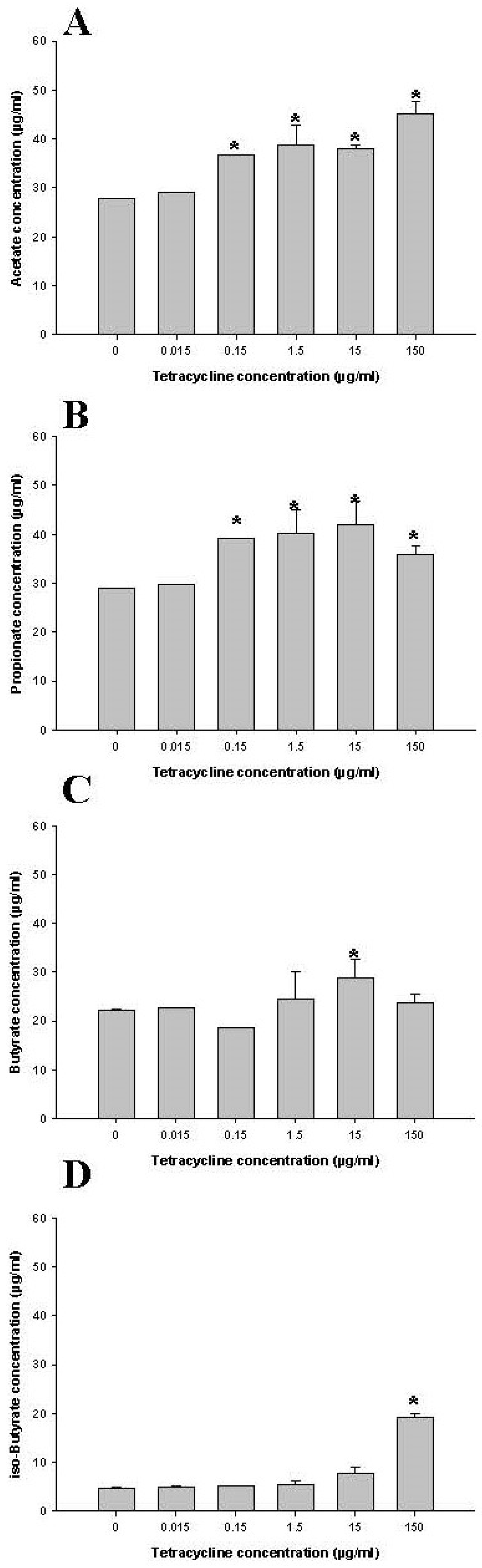
2.2.1. Short Chain Fatty Acid (SCFA) Concentration Changes
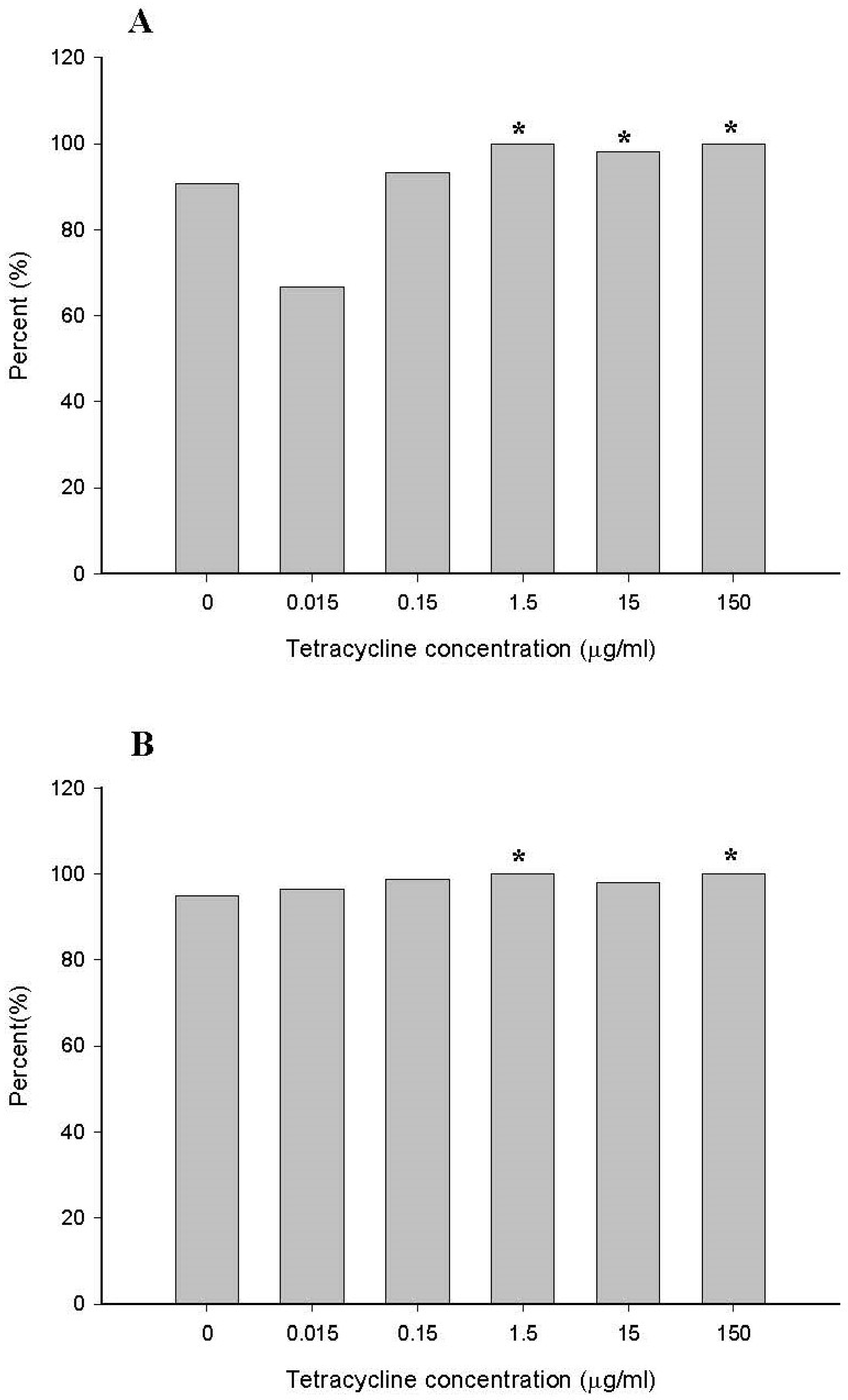
2.2.2. Comparison of Minimum Inhibitory Concentration (MIC) for the Family Enterobacteriaceae and the Genus Escherichia
3. Discussion
4. Materials and Methods
4.1. Human Fecal Samples
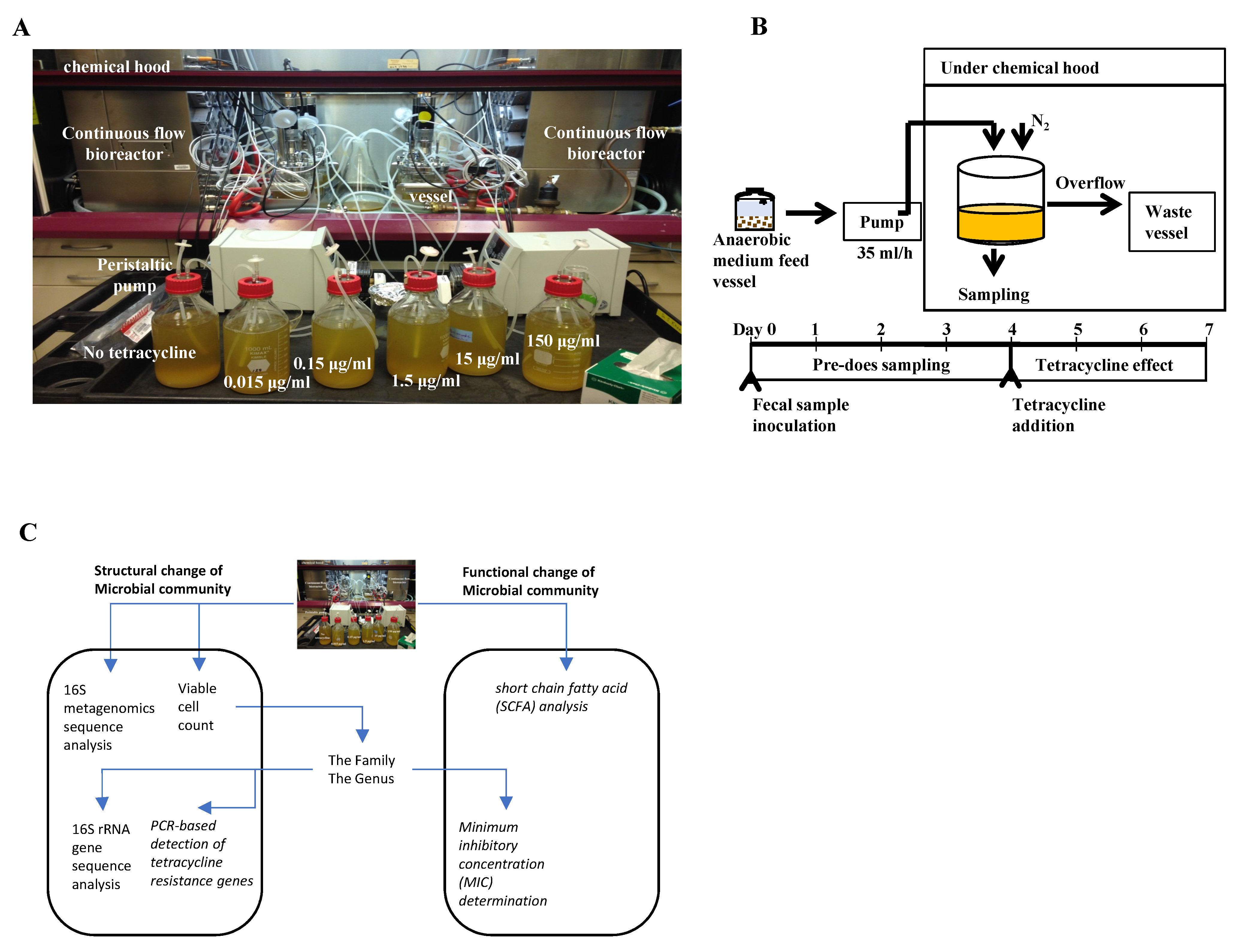
4.2. Establishment of Continuous Flow Bioreactor Cultures
4.3. Tetracycline Analysis by HPLC and LC-MS/MS
4.4. Structural Assessment of Microbiological Endpoints
4.4.1. Viable Bacterial Counting at the Microbial Community-Level
4.4.2. 16S rRNA Metagenomics Sequencing Analysis at the Microbial Community Level
4.4.3. Isolation and 16S rRNA Sequencing Analysis at the Family Level
4.4.4. PCR-Based Detection of Tetracycline Resistance Genes at the Family Level
4.5. Functional Assessment of Microbiological Endpoints
4.5.1. SCFA Analysis at the Microbial Community-Level
4.5.2. MIC Determination at the Family Level
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Sommer, F.; Backhed, F. The gut microbiota-masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Card, R.M.; Warburton, P.J.; MacLaren, N.; Mullany, P.; Allan, E.; Anjum, M.F. Application of microarray and functional-based screening methods for the detection of antimicrobial resistance genes in the microbiomes of healthy humans. PLoS ONE 2014, 9, e86428. [Google Scholar] [CrossRef] [Green Version]
- Francino, M.P. Antibiotics and the human gut microbiome: Dysbioses and accumulation of resistances. Front. Microbiol. 2015, 6, 1543. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Yang, X.; Qin, J.; Lu, N.; Cheng, G.; Wu, N.; Pan, Y.; Li, J.; Zhu, L.; Wang, X.; et al. Metagenome-wide analysis of antibiotic resistance genes in a large cohort of human gut microbiota. Nat. Commun. 2013, 4, 2151. [Google Scholar] [CrossRef] [Green Version]
- Pineiro, S.A.; Cerniglia, C.E. Antimicrobial drug residues in animal-derived foods: Potential impact on the human intestinal microbiome. J. Vet. Pharm. Ther. 2021, 44, 215–222. [Google Scholar] [CrossRef]
- Boobis, A.; Cerniglia, C.; Chicoine, A.; Fattori, V.; Lipp, M.; Reuss, R.; Verger, P.; Tritscher, A. Characterizing chronic and acute health risks of residues of veterinary drugs in food: Latest methodological developments by the joint FAO/WHO expert committee on food additives. Crit. Rev. Toxicol. 2017, 47, 885–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerniglia, C.E.; Pineiro, S.A.; Kotarski, S.F. An update discussion on the current assessment of the safety of veterinary antimicrobial drug residues in food with regard to their impact on the human intestinal microbiome. Drug Test. Anal. 2016, 8, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.I. A genomic view of our symbiosis with members of the gut microbiota. J. Pediatr. Gastroenterol. Nutr. 2005, 40, S28. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef] [Green Version]
- Wagner, R.D.; Johnson, S.J.; Cerniglia, C.E. In vitro model of colonization resistance by the enteric microbiota: Effects of antimicrobial agents used in food-producing animals. Antimicrob. Agents Chemother. 2008, 52, 1230–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carman, R.J.; Simon, M.A.; Petzold, H.E., III; Wimmer, R.F.; Batra, M.R.; Fernandez, A.H.; Miller, M.A.; Bartholomew, M. Antibiotics in the human food chain: Establishing no effect levels of tetracycline, neomycin, and erythromycin using a chemostat model of the human colonic microflora. Regul. Toxicol. Pharmacol. RTP 2005, 43, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Cerniglia, C.E.; Kotarski, S. Evaluation of veterinary drug residues in food for their potential to affect human intestinal microflora. Regul. Toxicol. Pharmacol. 1999, 29, 238–261. [Google Scholar] [CrossRef] [PubMed]
- Corpet, D.E. Antibiotic residues and drug resistance in human intestinal flora. Antimicrob. Agents Chemother. 1987, 31, 587–593. [Google Scholar] [CrossRef] [Green Version]
- Hirsh, D.C.; Burton, G.C.; Blenden, D.C. The effect of tetracycline upon establishment of Escherichia coli of bovine origin in the enteric tract of man. J. Appl. Bacteriol. 1974, 37, 327–333. [Google Scholar] [CrossRef]
- Perrin-Guyomard, A.; Cottin, S.; Corpet, D.E.; Boisseau, J.; Poul, J.M. Evaluation of residual and therapeutic doses of tetracycline in the human-flora-associated (HFA) mice model. Regul. Toxicol. Pharm. 2001, 34, 125–136. [Google Scholar] [CrossRef]
- Jung, J.Y.; Ahn, Y.; Khare, S.; Gokulan, K.; Pineiro, S.A.; Cerniglia, C.E. An in vitro study to assess the impact of tetracycline on the human intestinal microbiome. Anaerobe 2018, 49, 85–94. [Google Scholar] [CrossRef]
- Cerniglia, C.E.; Kotarski, S. Approaches in the safety evaluations of veterinary antimicrobial agents in food to determine the effects on the human intestinal microflora. J. Vet. Pharm. Ther. 2005, 28, 3–20. [Google Scholar] [CrossRef]
- EMEA. Committee for Veterinary Medicinal Products-Oxytetracycline, Tetracycline, Chlortetracycline: Summary Report (3). 1995. Available online: https://www.ema.europa.eu/en/documents/mrl-report/oxytetracycline-tetracycline-chlortetracycline-summary-report-3-committee-veterinary-medicinal_en.pdf (accessed on 15 June 2021).
- CLSI. Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria; Approved Standard, 8th ed.; CLSI Document M11-a8; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2012. [Google Scholar]
- JECFA. Tetracyclines: Oxytetracycline, Chlortetracycline, and Tetracycline (Addendum). 1998. Available online: http://www.inchem.org/documents/jecfa/jecmono/v041je07.htm (accessed on 15 June 2021).
- Ahn, Y.; Jung, J.Y.; Veach, B.T.; Khare, S.; Gokulan, K.; Pineiro, S.A.; Cerniglia, C.E. In vitro test systems to determine tetracycline residue binding to human feces. Regul. Toxicol. Pharm. 2018, 99, 105–115. [Google Scholar] [CrossRef]
- Hao, H.; Guo, W.; Iqbal, Z.; Cheng, G.; Wang, X.; Dai, M.; Huang, L.; Wang, Y.; Peng, D.; Liu, Z.; et al. Impact of cyadox on human colonic microflora in chemostat models. Regul. Toxicol. Pharmacol. RTP 2013, 67, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Carman, R.J.; Woodburn, M.A. Effects of low levels of ciprofloxacin on a chemostat model of the human colonic microflora. Regul. Toxicol. Pharmacol. RTP 2001, 33, 276–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilback, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carman, R.J.; Simon, M.A.; Fernandez, H.; Miller, M.A.; Bartholomew, M.J. Ciprofloxacin at low levels disrupts colonization resistance of human fecal microflora growing in chemostats. Regul. Toxicol. Pharmacol. RTP 2004, 40, 319–326. [Google Scholar] [CrossRef]
- Hao, H.; Zhou, S.; Cheng, G.; Dai, M.; Wang, X.; Liu, Z.; Wang, Y.; Yuan, Z. Effect of tulathromycin on colonization resistance, antimicrobial resistance, and virulence of human gut microbiota in chemostats. Front. Microbiol. 2016, 7, 477. [Google Scholar] [CrossRef] [Green Version]
- Hao, H.; Yao, J.; Wu, Q.; Wei, Y.; Dai, M.; Iqbal, Z.; Wang, X.; Wang, Y.; Huang, L.; Chen, D.; et al. Microbiological toxicity of tilmicosin on human colonic microflora in chemostats. Regul. Toxicol. Pharmacol. RTP 2015, 73, 201–208. [Google Scholar] [CrossRef]
- Ahn, Y.; Linder, S.W.; Veach, B.T.; Yan, S.S.; Fernandez, A.H.; Pineiro, S.A.; Cerniglia, C.E. In vitro enrofloxacin binding in human fecal slurries. Regul. Toxicol. Pharmacol. 2012, 62, 74–84. [Google Scholar] [CrossRef]
- Kim, B.S.; Kim, J.N.; Yoon, S.H.; Chun, J.; Cerniglia, C.E. Impact of enrofloxacin on the human intestinal microbiota revealed by comparative molecular analysis. Anaerobe 2012, 18, 310–320. [Google Scholar] [CrossRef]
- Goodman, A.L.; Kallstrom, G.; Faith, J.J.; Reyes, A.; Moore, A.; Dantas, G.; Gordon, J.I. Extensive personal human gut microbiota culture collections characterized and manipulated in gnotobiotic mice. Proc. Natl. Acad. Sci. USA 2011, 108, 6252–6257. [Google Scholar] [CrossRef] [Green Version]
- Rettedal, E.A.; Gumpert, H.; Sommer, M.O. Cultivation-based multiplex phenotyping of human gut microbiota allows targeted recovery of previously uncultured bacteria. Nat. Commun. 2014, 5, 4714. [Google Scholar] [CrossRef] [PubMed]
- Lau, J.T.; Whelan, F.J.; Herath, I.; Lee, C.H.; Collins, S.M.; Bercik, P.; Surette, M.G. Capturing the diversity of the human gut microbiota through culture-enriched molecular profiling. Genome Med. 2016, 8, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiergeist, A.; Glasner, J.; Reischl, U.; Gessner, A. Analyses of Intestinal Microbiota: Culture versus Sequencing. ILAR J. 2015, 56, 228–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagier, J.C.; Hugon, P.; Khelaifia, S.; Fournier, P.E.; La Scola, B.; Raoult, D. The rebirth of culture in microbiology through the example of culturomics to study human gut microbiota. Clin. Microbiol. Rev. 2015, 28, 237–264. [Google Scholar] [CrossRef] [Green Version]
- Lagier, J.C.; Million, M.; Hugon, P.; Armougom, F.; Raoult, D. Human gut microbiota: Repertoire and variations. Front. Cell Infect. Microbiol. 2012, 2, 136. [Google Scholar] [CrossRef] [Green Version]
- Cangelosi, G.A.; Meschke, J.S. Dead or alive: Molecular assessment of microbial viability. Appl. Environ. Microbiol. 2014, 80, 5884–5891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, G.T.; Macfarlane, S. Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 2012, 95, 50–60. [Google Scholar] [CrossRef]
- Rumney, C.; Rowland, I. Microbiological endpoint testing for veterinary antimicrobials—Setting MRLs. Food Chem. Toxicol. 1995, 33, 331–333. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feichtinger, M.; Mayrhofer, S.; Kneifel, W.; Domig, K.J. Tetracycline resistance patterns of Lactobacillus buchneri group strains. J. Food Prot. 2016, 79, 1741–1747. [Google Scholar] [CrossRef]
- Wang, N.; Hang, X.; Zhang, M.; Liu, X.; Yang, H. Analysis of newly detected tetracycline resistance genes and their flanking sequences in human intestinal bifidobacteria. Sci. Rep. 2017, 7, 6267. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.; Chan, C.H. Antibiotic resistance of pathogenic bacteria from odontogenic infections in Taiwan. J. Microbiol. Immunol. Infect. 2003, 36, 105–110. [Google Scholar]
- Roberts, M.C. Tetracycline resistance determinants: Mechanisms of action, regulation of expression, genetic mobility, and distribution. FEMS Microbiol. Rev. 1996, 19, 1–24. [Google Scholar] [CrossRef]
- Speer, B.S.; Shoemaker, N.B.; Salyers, A.A. Bacterial resistance to tetracycline: Mechanisms, transfer, and clinical significance. Clin. Microbiol. Rev. 1992, 5, 387–399. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, V.; Nam, H.M.; Sawant, A.A.; Headrick, S.I.; Nguyen, L.T.; Oliver, S.P. Distribution of tetracycline and streptomycin resistance genes and class 1 integrons in Enterobacteriaceae isolated from dairy and nondairy farm soils. Microb. Ecol. 2008, 55, 184–193. [Google Scholar] [CrossRef]
- Schmidt, A.S.; Bruun, M.S.; Dalsgaard, I.; Larsen, J.L. Incidence, distribution, and spread of tetracycline resistance determinants and integron-associated antibiotic resistance genes among motile aeromonads from a fish farming environment. Appl. Environ. Microbiol. 2001, 67, 5675–5682. [Google Scholar] [CrossRef] [Green Version]
- Bryan, A.; Shapir, N.; Sadowsky, M.J. Frequency and distribution of tetracycline resistance genes in genetically diverse, nonselected, and nonclinical Escherichia coli strains isolated from diverse human and animal sources. Appl. Environ. Microbiol. 2004, 70, 2503–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VICH. Studies to Evaluate the Safety of Residues of Veterinary Drugs in Human Food: General Approach to Establish a Microbiological ADI. 2019. Available online: https://vichsec.org/en/guidelines/pharmaceuticals/pharma-safety/antimicrobial-safety.html (accessed on 15 June 2021).
- Kim, B.S.; Kim, J.N.; Cerniglia, C.E. In vitro culture conditions for maintaining a complex population of human gastrointestinal tract microbiota. J. Biomed. Biotechnol. 2011, 2011, 838040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. Tolerances for Residues of New Animal Drugs in Food; Oxytetracycline-21 CFR Part 556. 1996. Available online: https://www.gpo.gov/fdsys/pkg/FR-1996-12-23/pdf/96-32430.pdf (accessed on 15 June 2021).
- Illumina. 16S Metagenomic Sequencing Library Preparation: Preparing 16S Ribosomal RNA Gene Amplicons for the Illumina MiSeq System. 2013. Available online: https://www.illumina.com/content/dam/illumina-support/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 15 June 2021).
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Warnes, M.G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R. Gplots: Various R Programming Tools for Plotting Data. 2016. Available online: https://cran.r-project.org/web/packages/gplots/index.html (accessed on 15 June 2021).
- Lane, D.J. 16S/23S rRNA Sequencing; John Wiley & Sons, Ltd.: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Peng, S.; Wang, Y.; Zhou, B.; Lin, X. Long-term application of fresh and composted manure increase tetracycline resistance in the arable soil of eastern China. Sci. Total Environ. 2015, 506–507, 279–286. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methods | Strengths | Weaknesses | References | |
|---|---|---|---|---|
| Structural change | Viable cell count | Easy and rapid screening Cost effective | Laborious and time-consuming process Low-resolution techniques Lack of sensitivity | Carman et al. [14] Corpet [16] Hirsh et al. [17] Perrin-Guyomard et al. [18] |
| 16S metagenomics sequence analysis PCR-based detection of tetracycline resistance genes 16S rRNA gene sequence analysis | Reduce assay time High-resolution techniques Sensitive and specific techniques Indigenous intestinal bacteria | Expensive equipment and trained staff Data interpretation Extensive sample preparation prior to analysis | Jung et al. [19] Kim et al. [32] | |
| Functional change | Short chain fatty acid (SCFA) analysis | Accurate technique for quantitative analysis Does not require extraction steps Rapid for time course Indigenous intestinal bacteria | Expensive equipment and trained staff Data interpretation | Carman et al. [14] Perrin-Guyomard et al. [18] |
| Minimum inhibitory concentration (MIC) determination | Rapid screening techniques Standard procedures Designed and optimized for the evaluation of pathogenic clinical isolates | Laborious and time-consuming process Not commensal intestinal bacteria Does not take into account the ecological interactions of bacteria in gastrointestinal tract | Carman et al. [14] Wagner et al. [13] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, Y.; Jung, J.Y.; Kweon, O.; Veach, B.T.; Khare, S.; Gokulan, K.; Piñeiro, S.A.; Cerniglia, C.E. Impact of Chronic Tetracycline Exposure on Human Intestinal Microbiota in a Continuous Flow Bioreactor Model. Antibiotics 2021, 10, 886. https://doi.org/10.3390/antibiotics10080886
Ahn Y, Jung JY, Kweon O, Veach BT, Khare S, Gokulan K, Piñeiro SA, Cerniglia CE. Impact of Chronic Tetracycline Exposure on Human Intestinal Microbiota in a Continuous Flow Bioreactor Model. Antibiotics. 2021; 10(8):886. https://doi.org/10.3390/antibiotics10080886
Chicago/Turabian StyleAhn, Youngbeom, Ji Young Jung, Ohgew Kweon, Brian T. Veach, Sangeeta Khare, Kuppan Gokulan, Silvia A. Piñeiro, and Carl E. Cerniglia. 2021. "Impact of Chronic Tetracycline Exposure on Human Intestinal Microbiota in a Continuous Flow Bioreactor Model" Antibiotics 10, no. 8: 886. https://doi.org/10.3390/antibiotics10080886
APA StyleAhn, Y., Jung, J. Y., Kweon, O., Veach, B. T., Khare, S., Gokulan, K., Piñeiro, S. A., & Cerniglia, C. E. (2021). Impact of Chronic Tetracycline Exposure on Human Intestinal Microbiota in a Continuous Flow Bioreactor Model. Antibiotics, 10(8), 886. https://doi.org/10.3390/antibiotics10080886