
Activity of Pyrazinamide against Mycobacterium tuberculosis at Neutral pH in PZA-S1 Minimal Medium

Abstract
:1. Introduction
2. Results
2.1. Determination of PZA Activity at Neutral pH in Minimal Medium with Amino Acid as the Nitrogen Source
2.2. Effect of Activity of PZA by pH in PZA-S1 Minimal Medium
2.3. Identification of PZA-Resistant M. tuberculosis at Neutral pH 6.8 in PZA-S1 Minimal Medium
2.4. Susceptibility Testing of First-Line TB Drugs and Determination of pH during Growth of M. tuberculosis at Neutral pH 6.8 in PZA-S1 Minimal Medium
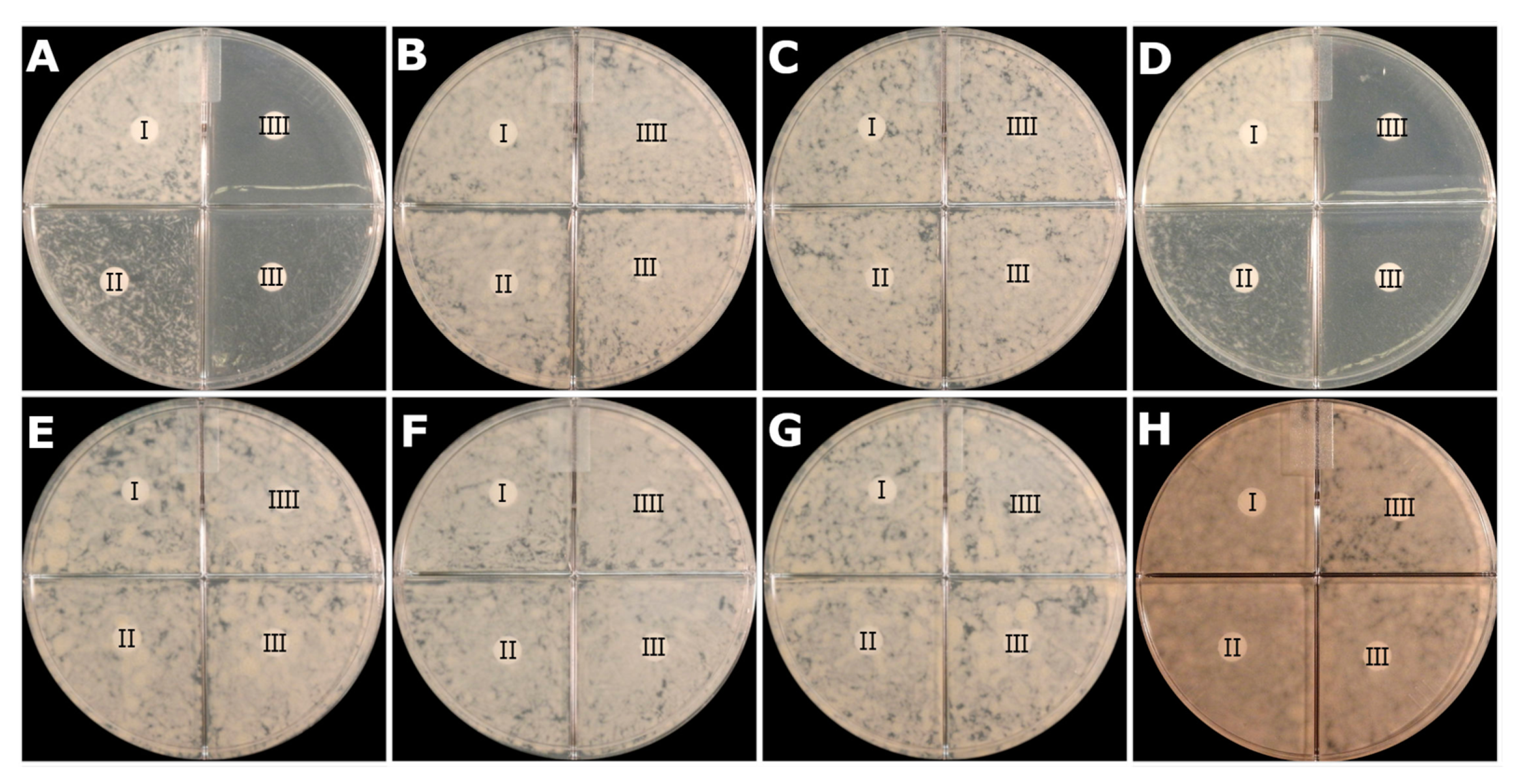
2.5. Activity of PZA at Neutral pH on PZA-S1 Agar Plate
3. Discussion
4. Materials and Methods
4.1. Culture Media
4.2. Bacterial Strain and Culture Conditions
4.3. Determination of MICs of PZA Using Broth Microdilution Method
4.4. Determination of Activity of PZA at Neutral pH on Agar Plate
4.5. Determination of the pH of Cultures during Growth of M. tuberculosis in PZA-S1 Media
5. Conclusions
6. Patents
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roberts, L. How COVID hurt the fight against other dangerous diseases. Nature 2021, 592, 502–504. [Google Scholar] [CrossRef]
- World Health Organization. Global Tuberculosis Report 2020; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Zhang, Y.; Shi, W.; Zhang, W.; Mitchison, D. Mechanisms of Pyrazinamide Action and Resistance. Microbiol. Spectr. 2014, 2, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Dorman, S.E.; Nahid, P.; Kurbatova, E.V.; Phillips, P.P.J.; Bryant, K.; Dooley, K.E.; Engle, M.; Goldberg, S.V.; Phan, H.T.T.; Hakim, J.; et al. Four-Month Rifapentine Regimens with or without Moxifloxacin for Tuberculosis. N. Engl. J. Med. 2021, 384, 1705–1718. [Google Scholar] [CrossRef]
- McDermott, W.; Tompsett, R. Activation of pyrazinamide and nicotinamide in acidic environments in vitro. Am. Rev. Tuberc. 1954, 70, 748–754. [Google Scholar]
- Scorpio, A.; Zhang, Y. Mutations in pncA, a gene encoding pyrazinamidase/nicotinamidase, cause resistance to the antituberculous drug pyrazinamide in tubercle bacillus. Nat. Med. 1996, 2, 662–667. [Google Scholar] [CrossRef]
- Konno, K.; Feldmann, F.M.; McDermott, W. Pyrazinamide susceptibility and amidase activity of tubercle bacilli. Am. Rev. Respir. Dis. 1967, 95, 461–469. [Google Scholar] [CrossRef]
- Zimhony, O.; Cox, J.S.; Welch, J.T.; Vilcheze, C.; Jacobs, W.R., Jr. Pyrazinamide inhibits the eukaryotic-like fatty acid synthetase I (FASI) of Mycobacterium tuberculosis. Nat. Med. 2000, 6, 1043–1047. [Google Scholar] [CrossRef]
- Shi, W.; Zhang, X.; Jiang, X.; Yuan, H.; Lee, J.S.; Barry, C.E., 3rd; Wang, H.; Zhang, W.; Zhang, Y. Pyrazinamide inhibits trans-translation in Mycobacterium tuberculosis. Science 2011, 333, 1630–1632. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, Y.; Bi, J.; Cai, Q.; Liao, X.; Li, W.; Guo, C.; Zhang, Q.; Lin, T.; Zhao, Y.; et al. Structural basis for targeting the ribosomal protein S1 of Mycobacterium tuberculosis by pyrazinamide. Mol. Microbiol. 2015, 95, 791–803. [Google Scholar] [CrossRef]
- Keiler, K.C. Biology of trans-translation. Annu. Rev. Microbiol. 2008, 62, 133–151. [Google Scholar] [CrossRef]
- Shi, W.; Chen, J.; Feng, J.; Cui, P.; Zhang, S.; Weng, X.; Zhang, W.; Zhang, Y. Aspartate decarboxylase (PanD) as a new target of pyrazinamide in Mycobacterium tuberculosis. Emerg. Microbes. Infect. 2014, 3, e58. [Google Scholar] [CrossRef]
- Gopal, P.; Yee, M.; Sarathy, J.; Low, J.L.; Sarathy, J.P.; Kaya, F.; Dartois, V.; Gengenbacher, M.; Dick, T. Pyrazinamide Resistance Is Caused by Two Distinct Mechanisms: Prevention of Coenzyme A Depletion and Loss of Virulence Factor Synthesis. ACS Infect. Dis. 2016, 2, 616–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopal, P.; Nartey, W.; Ragunathan, P.; Sarathy, J.; Kaya, F.; Yee, M.; Setzer, C.; Manimekalai, M.S.S.; Dartois, V.; Gruber, G.; et al. Pyrazinoic Acid Inhibits Mycobacterial Coenzyme A Biosynthesis by Binding to Aspartate Decarboxylase PanD. ACS Infect. Dis. 2017, 3, 807–819. [Google Scholar] [CrossRef]
- Gopal, P.; Tasneen, R.; Yee, M.; Lanoix, J.P.; Sarathy, J.; Rasic, G.; Li, L.; Dartois, V.; Nuermberger, E.; Dick, T. In Vivo-Selected Pyrazinoic Acid-Resistant Mycobacterium tuberculosis Strains Harbor Missense Mutations in the Aspartate Decarboxylase PanD and the Unfoldase ClpC1. ACS Infect. Dis. 2017, 3, 492–501. [Google Scholar] [CrossRef]
- Njire, M.; Wang, N.; Wang, B.; Tan, Y.; Cai, X.; Liu, Y.; Mugweru, J.; Guo, J.; Hameed, H.M.A.; Tan, S.; et al. Pyrazinoic Acid Inhibits a Bifunctional Enzyme in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2017, 61, e00070-17. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Cui, P.; Shi, W.; Li, Q.; Zhang, W.; Li, M.; Zhang, Y. Pyrazinoic Acid Inhibits the Bifunctional Enzyme (Rv2783) in Mycobacterium tuberculosis by Competing with tmRNA. Pathogens 2019, 8, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boshoff, H.I.; Mizrahi, V.; Barry, C.E., 3rd. Effects of pyrazinamide on fatty acid synthesis by whole mycobacterial cells and purified fatty acid synthase I. J. Bacteriol. 2002, 184, 2167–2172. [Google Scholar] [CrossRef] [Green Version]
- Dillon, N.A.; Peterson, N.D.; Feaga, H.A.; Keiler, K.C.; Baughn, A.D. Anti-tubercular Activity of Pyrazinamide is Independent of trans-Translation and RpsA. Sci. Rep. 2017, 7, 6135. [Google Scholar] [CrossRef]
- Dillon, N.A.; Peterson, N.D.; Rosen, B.C.; Baughn, A.D. Pantothenate and pantetheine antagonize the antitubercular activity of pyrazinamide. Antimicrob. Agents Chemother. 2014, 58, 7258–7263. [Google Scholar] [CrossRef] [Green Version]
- APHL. Issues in Mycobacterium tuberculosis Complex (MTBC) Drug Susceptibility Testing: Pyrazinamide (PZA); Association of Public Health Laboratories: Washington, DC, USA, 2016. [Google Scholar]
- Köser, C.U.; Cirillo, D.M.; Miotto, P. How to optimally combine genotypic and phenotypic drug susceptibility testing methods for pyrazinamide. Antimicrob. Agents Chemother. 2020, 64, e01003-20. [Google Scholar] [CrossRef]
- Salfinger, M. Drug-Resistant Tuberculosis: Inconsistent Results of Pyrazinamide Susceptibility Testing-Reply. JAMA 1995, 273, 917. [Google Scholar] [CrossRef]
- Zhang, Y.; Permar, S.; Sun, Z. Conditions that may affect the results of susceptibility testing of Mycobacterium tuberculosis to pyrazinamide. J. Med. Microbiol. 2002, 51, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.P.; Billington, O.J.; McHugh, T.D.; Mitchison, D.A.; Gillespie, S.H. Comparison of phenotypic and genotypic methods for pyrazinamide susceptibility testing with Mycobacterium tuberculosis. J. Clin. Microbiol. 2000, 38, 3686–3688. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, M.G.; Soeters, H.M.; Warren, R.M.; York, T.; Sampson, S.L.; Streicher, E.M.; van Helden, P.D.; van Rie, A. A Global Perspective on Pyrazinamide Resistance: Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0133869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitfield, M.G.; Warren, R.M.; Streicher, E.M.; Sampson, S.L.; Sirgel, F.A.; van Helden, P.D.; Mercante, A.; Willby, M.; Hughes, K.; Birkness, K.; et al. Mycobacterium tuberculosis pncA Polymorphisms That Do Not Confer Pyrazinamide Resistance at a Breakpoint Concentration of 100 Micrograms per Milliliter in MGIT. J. Clin. Microbiol. 2015, 53, 3633–3635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naluyange, R.; Mboowa, G.; Komakech, K.; Semugenze, D.; Kateete, D.P.; Ssengooba, W. High prevalence of phenotypic pyrazinamide resistance and its association with pncA gene mutations in Mycobacterium tuberculosis isolates from Uganda. PLoS ONE 2020, 15, e0232543. [Google Scholar] [CrossRef]
- Pang, Y.; Zhang, Z.; Wang, Y.; Wang, S.; Song, Y.; Zhao, B.; Zhou, Y.; Ou, X.; Li, Q.; Xia, H.; et al. Genotyping and Prevalence of Pyrazinamide- and Moxifloxacin-Resistant Tuberculosis in China, 2000 to 2010. Antimicrob. Agents Chemother. 2017, 61, e02170-16. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Consultation on the Translation of Tuberculosis Research into Global Policy Guidelines: 2–4 March 2021: Meeting Report; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- World Health Organization. Policy Guidance on Drug-Susceptibility Testing (DST) of Second-Line Antituberculosis Drugs; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Steenken, W., Jr.; Wolinsky, E. The antituberculous activity of pyrazinamide in vitro and in the guinea pig. Am. Rev. Tuberc. 1954, 70, 367–369. [Google Scholar] [CrossRef]
- Salfinger, M.; Heifets, L.B. Determination of pyrazinamide MICs for Mycobacterium tuberculosis at different pHs by the radiometric method. Antimicrob. Agents Chemother. 1988, 32, 1002–1004. [Google Scholar] [CrossRef] [Green Version]
- den Hertog, A.L.; Menting, S.; Pfeltz, R.; Warns, M.; Siddiqi, S.H.; Anthony, R.M. Pyrazinamide Is Active against Mycobacterium tuberculosis Cultures at Neutral pH and Low Temperature. Antimicrob. Agents Chemother. 2016, 60, 4956–4960. [Google Scholar] [CrossRef] [Green Version]
- Gu, P.; Constantino, L.; Zhang, Y. Enhancement of the antituberculosis activity of weak acids by inhibitors of energy metabolism but not by anaerobiosis suggests that weak acids act differently from the front-line tuberculosis drug pyrazinamide. J. Med. Microbiol. 2008, 57, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Peterson, N.D.; Rosen, B.C.; Dillon, N.A.; Baughn, A.D. Uncoupling Environmental pH and Intrabacterial Acidification from Pyrazinamide Susceptibility in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2015, 59, 7320–7326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Q.; Chen, Z.F.; Li, Y.Y.; Zhang, Y.; Ren, Y.; Fu, Z.; Xu, S.Q. Nutrient-starved incubation conditions enhance pyrazinamide activity against Mycobacterium tuberculosis. Chemotherapy 2007, 53, 338–343. [Google Scholar] [CrossRef]
- Beck, B.P.a.M. Beiträge zur ernährungsphysiologie des tuberkelbacillus. Z. Für Hyg. Und Infekt. 1894, 18, 128–152. [Google Scholar]
- Long, E.R. The nutrition of acid-fast bacteria. Am. Rev. Tuberc. 1922, 5, 857–869. [Google Scholar]
- Dubos, R.J.; Middlebrook, G. Media for tubercle bacilli. Am. Rev. Tuberc. 1947, 56, 334–345. [Google Scholar] [PubMed]
- Lyon, R.H.; Hall, W.H.; Costas-Martinez, C. Utilization of Amino Acids During Growth of Mycobacterium tuberculosis in Rotary Cultures. Infect. Immun. 1970, 1, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, E.R. A Study in Fundamentals of the Nutrition of the Tubercle Bacillus: The Utilization of Some Amino-Acids and Ammonium Salts. Am. Rev. Tuberc. 1919, 3, 86–108. [Google Scholar]
- Dubos, R.J.; Davis, B.D. Factors affecting the growth of tubercle bacilli in liquid media. J. Exp. Med. 1946, 83, 409–423. [Google Scholar] [CrossRef] [Green Version]
- Sauton, B. Sur la nutrition minérale du bacille tuberculeux. CR Acad. Sci. 1912, 155, 860–861. [Google Scholar]
- Tarshis, M.S.; Weed, W.A., Jr. Lack of significant in vitro sensitivity of Mycobacterium tuberculosis to pyrazinamide on three different solid media. Am. Rev. Tuberc. 1953, 67, 391–395. [Google Scholar]
- Marks, J. A ‘Stepped Ph’ Technique for the Estimation of Pyrazinamide Sensitivity. Tubercle 1964, 45, 47–50. [Google Scholar] [CrossRef]
- Zwolska-Opalanska, Z. Difficulties in the determination of resistance of tubercle bacilli to pyrazinamide in vitro. Gruzlica 1966, 34, 95–99. [Google Scholar]
- Hall, L.; Jude, K.P.; Clark, S.L.; Wengenack, N.L. Antimicrobial susceptibility testing of Mycobacterium tuberculosis complex for first and second line drugs by broth dilution in a microtiter plate format. J. Vis. Exp. 2011, 52, e3094. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ma, Y.F.; Jiang, G.L.; Chen, S.T.; Wang, G.R.; Huang, H.R. Sensititre(R) MYCOTB MIC plate for drug susceptibility testing of Mycobacterium tuberculosis complex isolates. Int. J. Tuberc. Lung Dis. 2016, 20, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Held, P. Using phenol red to assess pH in tissue culture media. BioTek Appl. Note1 2018, 1–7. Available online: https://www.biotek.com/resources/application-notes/using-phenol-red-to-assess-ph-in-tissue-culture-media/ (accessed on 22 March 2018).
- Whalen, J.W.; Mallmann, W. Charcoal agar media for the cultivation of Mycobacterium tuberculosis. Am. Rev. Tuberc. Pulm. Dis. 1955, 71, 382–389. [Google Scholar]
- Wayne, L.G.; Krasnow, I. Preparation of tuberculosis susceptibility testing mediums by means of impregnated disks. Tech. Bull. Regist. Med. Technol. 1966, 36, 115–117. [Google Scholar] [CrossRef]
- Wade, M.M.; Zhang, Y. Anaerobic incubation conditions enhance pyrazinamide activity against Mycobacterium tuberculosis. J. Med. Microbiol. 2004, 53, 769–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, D.; Waddell, S.J.; Mitchison, D.A. Effects of low incubation temperatures on the bactericidal activity of anti-tuberculosis drugs. J. Antimicrob. Chemother. 2011, 66, 146–150. [Google Scholar] [CrossRef]
- Zhang, Y.; Wade, M.M.; Scorpio, A.; Zhang, H.; Sun, Z. Mode of action of pyrazinamide: Disruption of Mycobacterium tuberculosis membrane transport and energetics by pyrazinoic acid. J. Antimicrob. Chemother. 2003, 52, 790–795. [Google Scholar] [CrossRef]
- Wade, M.M.; Zhang, Y. Effects of weak acids, UV and proton motive force inhibitors on pyrazinamide activity against Mycobacterium tuberculosis in vitro. J. Antimicrob. Chemother. 2006, 58, 936–941. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Scorpio, A.; Nikaido, H.; Sun, Z. Role of acid pH and deficient efflux of pyrazinoic acid in unique susceptibility of Mycobacterium tuberculosis to pyrazinamide. J. Bacteriol. 1999, 181, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Cole, S.T.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry, C.E., 3rd; et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 1998, 393, 537–544. [Google Scholar] [CrossRef]
- Borah, K.; Beyss, M.; Theorell, A.; Wu, H.; Basu, P.; Mendum, T.A.; Nh, K.; Beste, D.J.V.; McFadden, J. Intracellular Mycobacterium tuberculosis Exploits Multiple Host Nitrogen Sources during Growth in Human Macrophages. Cell Rep. 2019, 29, 3580–3591.e3584. [Google Scholar] [CrossRef] [PubMed]
- Youmans, A.S.; Youmans, G.P. Studies on the metabolism of Mycobacterium tuberculosis. V. The effect of amino acids on the growth of M. tuberculosis var. hominis. J. Bacteriol. 1954, 67, 734–737. [Google Scholar] [CrossRef] [Green Version]
- Lyon, R.H.; Hall, W.H.; Costas-Martinez, C. Effect of L-asparagine on growth of Mycobacterium tuberculosis and on utilization of other amino acids. J. Bacteriol. 1974, 117, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Dingle, J.H.; Weinzirl, J. The Biology of the Tubercle Bacillus: II. The Asparagin and Glycerin Metabolism of the Tubercle Bacillus. J. Bacteriol. 1932, 23, 281. [Google Scholar] [CrossRef] [Green Version]
- Steele, M.A.; Des Prez, R.M. The role of pyrazinamide in tuberculosis chemotherapy. Chest 1988, 94, 845–850. [Google Scholar] [CrossRef]
- Heifets, L.; Lindholm-Levy, P. Pyrazinamide sterilizing activity in vitro against semidormant Mycobacterium tuberculosis bacterial populations. Am. Rev. Respir. Dis. 1992, 145, 1223–1225. [Google Scholar] [CrossRef]
- Zhang, Y.; Mitchison, D. The curious characteristics of pyrazinamide: A review. Int. J. Tuberc. Lung Dis. 2003, 7, 6–21. [Google Scholar] [PubMed]
- Rohde, K.; Yates, R.M.; Purdy, G.E.; Russell, D.G. Mycobacterium tuberculosis and the environment within the phagosome. Immunol. Rev. 2007, 219, 37–54. [Google Scholar] [CrossRef] [PubMed]
- Sarathy, J.P.; Dartois, V. Caseum: A Niche for Mycobacterium tuberculosis Drug-Tolerant Persisters. Clin. Microbiol. Rev. 2020, 33, e00159-19. [Google Scholar] [CrossRef] [PubMed]
- Smith, T. The Reaction Curve Of the Human and the Bovine type of the Tubercle Bacillus in Glycerine Bouillon. J. Med. Res. 1910, 23, 185–204. [Google Scholar]
- Long, E.R.; Major, A.L. A Method of Following Reaction Changes in Cultures of Acid-fast Bacteria. Am. Rev. Tuberc. 1921, 5, 715–722. [Google Scholar]
- Mok, S.; Roycroft, E.; Flanagan, P.R.; Montgomery, L.; Borroni, E.; Rogers, T.R.; Fitzgibbon, M.M. Overcoming the Challenges of Pyrazinamide Susceptibility Testing in Clinical Mycobacterium tuberculosis isolates. Antimicrob. Agents Chemother. 2021. [Google Scholar] [CrossRef] [PubMed]
- Stottmeier, K.D.; Beam, R.E.; Kubica, G.P. Determination of drug susceptibility of mycobacteria to pyrazinamide in 7H10 agar. Am. Rev. Respir. Dis. 1967, 96, 1072–1075. [Google Scholar] [CrossRef]
- World Health Organization. Technical Manual for Drug Susceptibility Testing of Medicines Used in the Treatment of Tuberculosis; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Brander, E. A simple way of detecting pyrazinamide resistance. Tubercle 1972, 53, 128–131. [Google Scholar] [CrossRef]
- Hall, L.; Jude, K.P.; Clark, S.L.; Dionne, K.; Merson, R.; Boyer, A.; Parrish, N.M.; Wengenack, N.L. Evaluation of the Sensititre MycoTB plate for susceptibility testing of the Mycobacterium tuberculosis complex against first- and second-line agents. J. Clin. Microbiol. 2012, 50, 3732–3734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rancoita, P.M.V.; Cugnata, F.; Gibertoni Cruz, A.L.; Borroni, E.; Hoosdally, S.J.; Walker, T.M.; Grazian, C.; Davies, T.J.; Peto, T.E.A.; Crook, D.W.; et al. Validating a 14-Drug Microtiter Plate Containing Bedaquiline and Delamanid for Large-Scale Research Susceptibility Testing of Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2018, 62, e00344-18. [Google Scholar] [CrossRef] [Green Version]
- Humphries, R.M.; Kircher, S.; Ferrell, A.; Krause, K.M.; Malherbe, R.; Hsiung, A.; Burnham, C.A. The Continued Value of Disk Diffusion for Assessing Antimicrobial Susceptibility in Clinical Laboratories: Report from the Clinical and Laboratory Standards Institute Methods Development and Standardization Working Group. J. Clin. Microbiol. 2018, 56, e00437-18. [Google Scholar] [CrossRef] [Green Version]
- Afanas’eva Iu, P.; Dorozhkova, I.R. Paper disk methods for determination of the drug sensitivity of Mycobacterium tuberculosis to the 2d order preparations. II. Lab. Delo 1969, 5, 296–298. [Google Scholar] [PubMed]
- Heinrichs, M.T.; May, R.J.; Heider, F.; Reimers, T.; SK, B.S.; Peloquin, C.A.; Derendorf, H. Mycobacterium tuberculosis Strains H37ra and H37rv have equivalent minimum inhibitory concentrations to most antituberculosis drugs. Int. J. Mycobacteriol. 2018, 7, 156–161. [Google Scholar] [CrossRef]
- Shi, W.; Chen, J.; Zhang, S.; Zhang, W.; Zhang, Y. Identification of Novel Mutations in LprG (rv1411c), rv0521, rv3630, rv0010c, ppsC, and cyp128 Associated with Pyrazinoic Acid/Pyrazinamide Resistance in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2018, 62, e00430-18. [Google Scholar] [CrossRef] [Green Version]
- Woods, G.L.; Brown-Elliott, B.A.; Conville, P.S.; Desmond, E.P.; Hall, G.S.; Lin, G.; Pfyffer, G.E.; Ridderhof, J.C.; Siddiqi, S.H.; Wallace, R.J., Jr. Susceptibility Testing of Mycobacteria, Nocardiae, and Other Aerobic Actinomycetes; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011. [Google Scholar]
- Kaniga, K.; Cirillo, D.M.; Hoffner, S.; Ismail, N.A.; Kaur, D.; Lounis, N.; Metchock, B.; Pfyffer, G.E.; Venter, A. A Multilaboratory, Multicountry Study To Determine MIC Quality Control Ranges for Phenotypic Drug Susceptibility Testing of Selected First-Line Antituberculosis Drugs, Second-Line Injectables, Fluoroquinolones, Clofazimine, and Linezolid. J. Clin. Microbiol. 2016, 54, 2963–2968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esser, V.M.; Elefson, D.E. Experiences with the Kirby-Bauer method of antibiotic susceptibility testing. Am. J. Clin. Pathol. 1970, 54, 193–198. [Google Scholar] [CrossRef]
- Hudzicki, J. Kirby-Bauer Disk Diffusion Susceptibility Test Protocol. Available online: https://asm.org/Protocols/Kirby-Bauer-Disk-Diffusion-Susceptibility-Test-Pro (accessed on 8 December 2009).
- ISO. ISO 20776-1: 2006 Clinical Laboratory Testing and In Vitro Diagnostic Test Systems–Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test Devices–Part 1: Reference Method for Testing the In Vitro Activity of Antimicrobial Agents against Rapidly Growing Aerobic Bacteria Involved in Infectious Diseases; International Organization for Standardization: Geneva, Switzerland, 2006; p. 19. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nitrogen Source and Other Ingredients @ | Concentration (g/L) | MIC of PZA (mg/L) | Status of Growth c |
|---|---|---|---|
| L-Aspartic acid (L-Asp) | 2.0 | >1000 | +++ |
| L-Glutamic acid (L-Glu) | 2.0 | >1000 | +++ |
| L-Asparagine (L-Asn) | 2.0 | >1000 | +++ |
| 0.5 | >1000 | +++ | |
| 0.1 | >1000 | +++ | |
| L-Glutamine (L-Gln) | 2.0 | >1000 | +++ |
| L-Cysteine (L-Cys) | 2.0 | >1000 | ++ |
| L-Histidine (L-His) | 2.0 | 500 | ++ |
| L-Proline (L-Pro) | 2.0 | 500 | ++ |
| L-Valine (L-Val) * | 2.0 | 500 | +++ |
| L-Leucine (L-Leu) | 2.0 | 500 | ++ |
| L-Arginine (L-Arg) | 2.0 | 250 | + |
| L-Lysine (L-Lys) | 2.0 | 250 | + |
| L-Serine (L-Ser) | 2.0 | 250 | + |
| L-Threonine (L-Thr) | 2.0 | 250 | + |
| L-Isoleucine (L-Ile) | 2.0 | 250 | ++ |
| Glycine (Gly) | 2.0 | 250 | +++ |
| L-Alanine (L-Ala) | 4.0 | 125 | +++ |
| 2.0 | 125 | +++ | |
| 1.0 | 125 | +++ | |
| 0.5 | 125 | +++ | |
| L-Methionine (L-Met) | 2.0 | ND | - |
| L-Phenylalanine (L-Phe) | 2.0 | ND | - |
| L-Tryptophan (L-Trp) | 2.0 | ND | - |
| L-Tyrosine * (L-Tyr) | 2.0 | ND | - |
| pH | Visible Bacterial Growth with PZA (mg/L) | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 12.5 | 25 | 50 | 100 | 200 | 400 | 800 | |
| 8.5 * | + | + | + | + | - | - | - | - |
| 8.0 | + | + | + | + | + | - | - | - |
| 7.5 | + | + | + | + | + | - | - | - |
| 7.0 | + | + | + | + | + | - | - | - |
| 6.5 | + | + | + | + | + | - | - | - |
| 6.0 | + | + | + | + | - | - | - | - |
| 5.5 * | + | + | - | - | - | - | - | - |
| 5.0 | - | - | - | - | - | - | - | - |
| Day of Incubation at 37 °C | |||||
|---|---|---|---|---|---|
| 0 | 9 | 15 | 18 | 30 | |
| pH of culture | 6.8 | ~6.9 | ~7.0 | ~7.2 | ~6.5 |
| pH of control * | 6.8 | 6.8 | 6.8 | 6.8 | 6.8 |
| M. tuberculosis Strain | PZA Susceptibility * | pncA Mutation | PncA Amino Acid Substitution |
|---|---|---|---|
| M. tuberculosis H37Ra (ATCC 25177) | S | Wild type | |
| M. tuberculosis H37Ra (NR-122) | S | Wild type | |
| M. tuberculosis H37Ra PZAR #6 | R | T476C | L159P |
| M. tuberculosis H37Ra PZAR #78 | R | T412C | C138R |
| M. tuberculosis H37Ra PZAR #98 | R | T202C | W68R |
| M. tuberculosis H37Ra PZAR #106 | R | A146G | D49G |
| M. tuberculosis H37Ra PZAR #186 | R | G395C | G132A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, W. Activity of Pyrazinamide against Mycobacterium tuberculosis at Neutral pH in PZA-S1 Minimal Medium. Antibiotics 2021, 10, 909. https://doi.org/10.3390/antibiotics10080909
Shi W. Activity of Pyrazinamide against Mycobacterium tuberculosis at Neutral pH in PZA-S1 Minimal Medium. Antibiotics. 2021; 10(8):909. https://doi.org/10.3390/antibiotics10080909
Chicago/Turabian StyleShi, Wanliang. 2021. "Activity of Pyrazinamide against Mycobacterium tuberculosis at Neutral pH in PZA-S1 Minimal Medium" Antibiotics 10, no. 8: 909. https://doi.org/10.3390/antibiotics10080909
APA StyleShi, W. (2021). Activity of Pyrazinamide against Mycobacterium tuberculosis at Neutral pH in PZA-S1 Minimal Medium. Antibiotics, 10(8), 909. https://doi.org/10.3390/antibiotics10080909