
Syzygium aromaticum Extracts as a Potential Antibacterial Inhibitors against Clinical Isolates of Acinetobacter baumannii: An In-Silico-Supported In-Vitro Study


 and
and
Abstract
:1. Introduction
2. Results
2.1. Patients and Clinical Characteristics
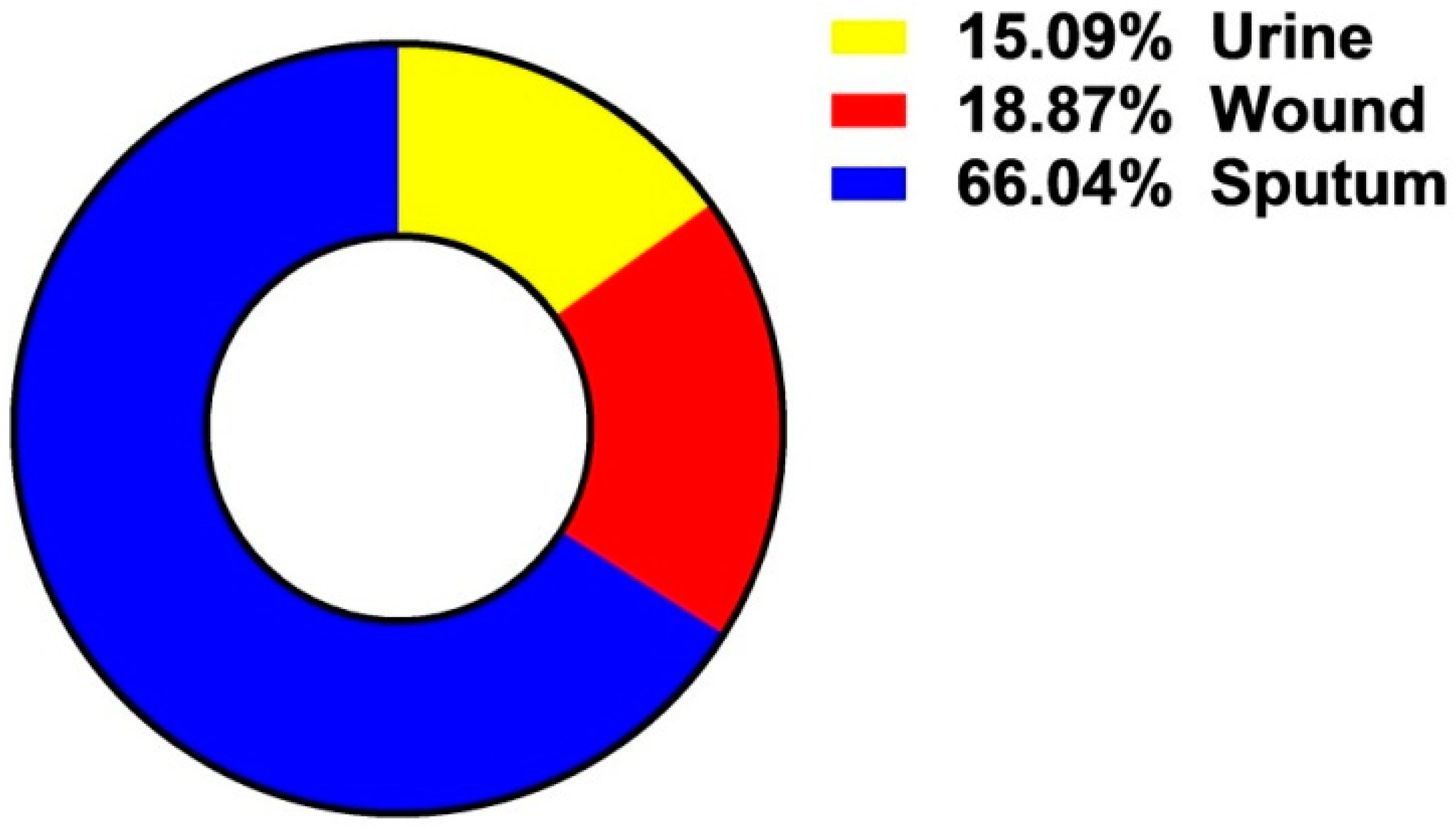
The Incidence of A. baumannii Isolates among Examined Clinical Samples
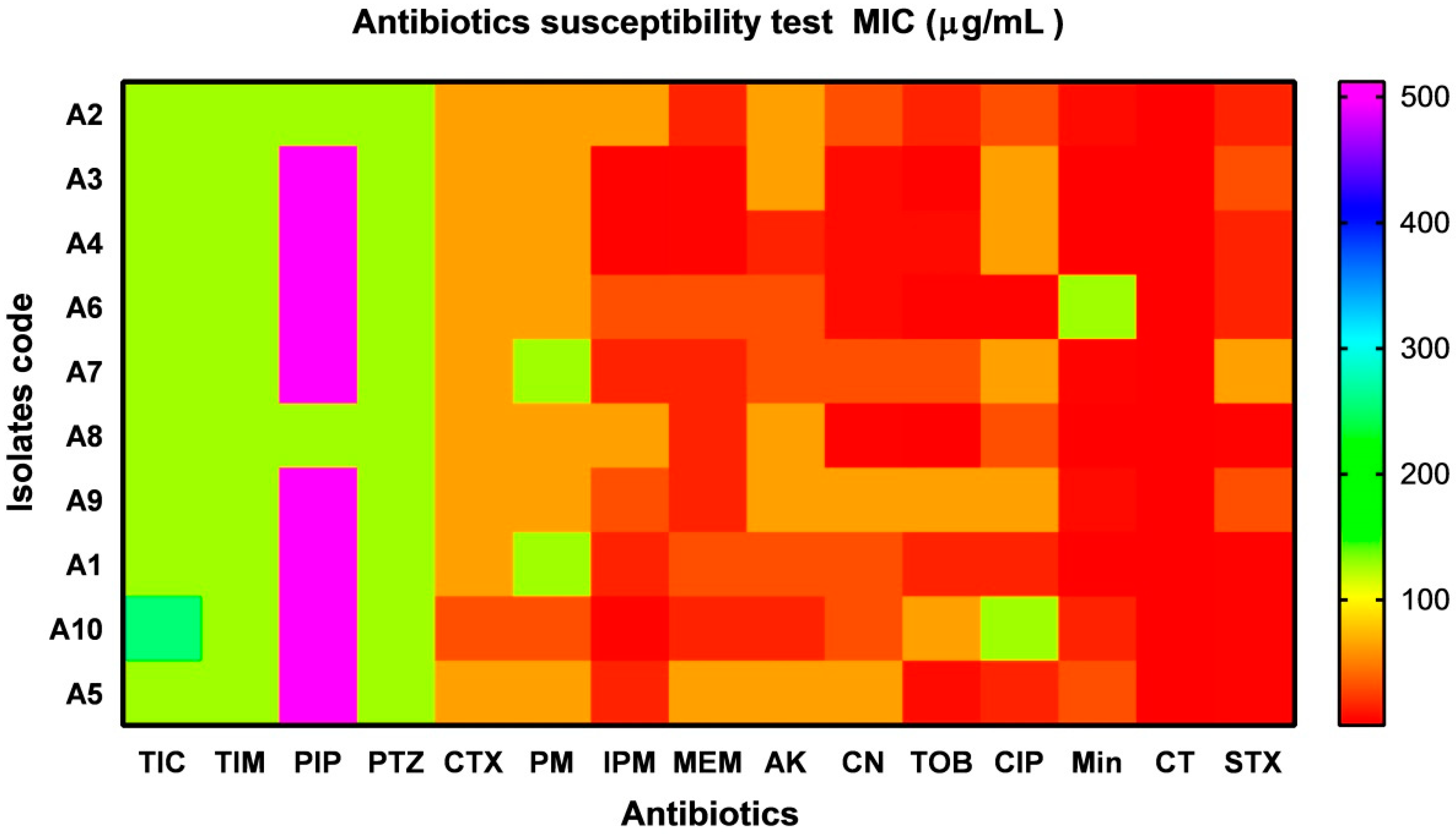
2.2. Antimicrobial Susceptibility Testing
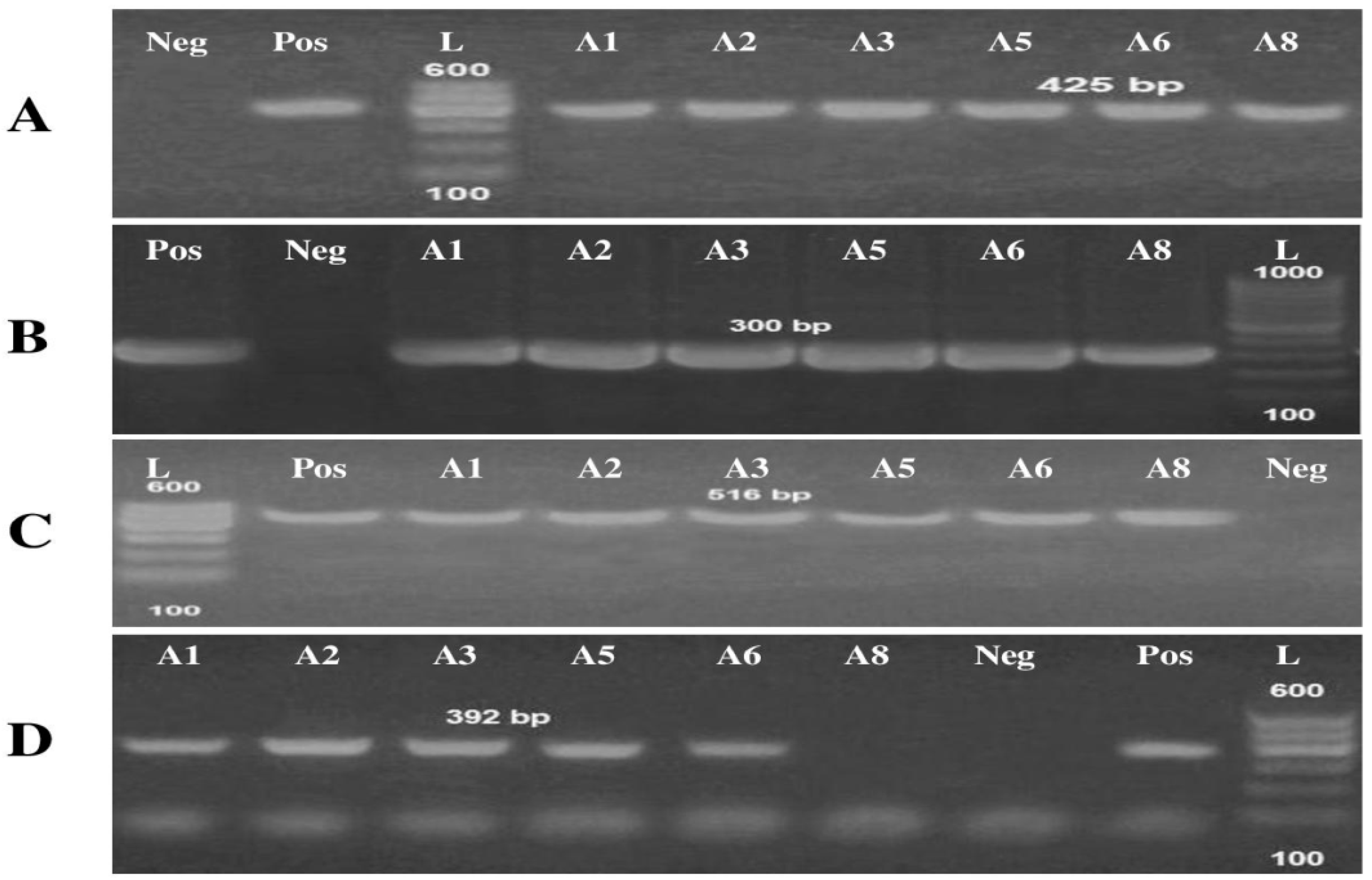
Detection of Virulence and Antibiotic-Resistant Genes
2.3. Gas Chromatography-Mass Spectrometry (GC/MS) Analysis
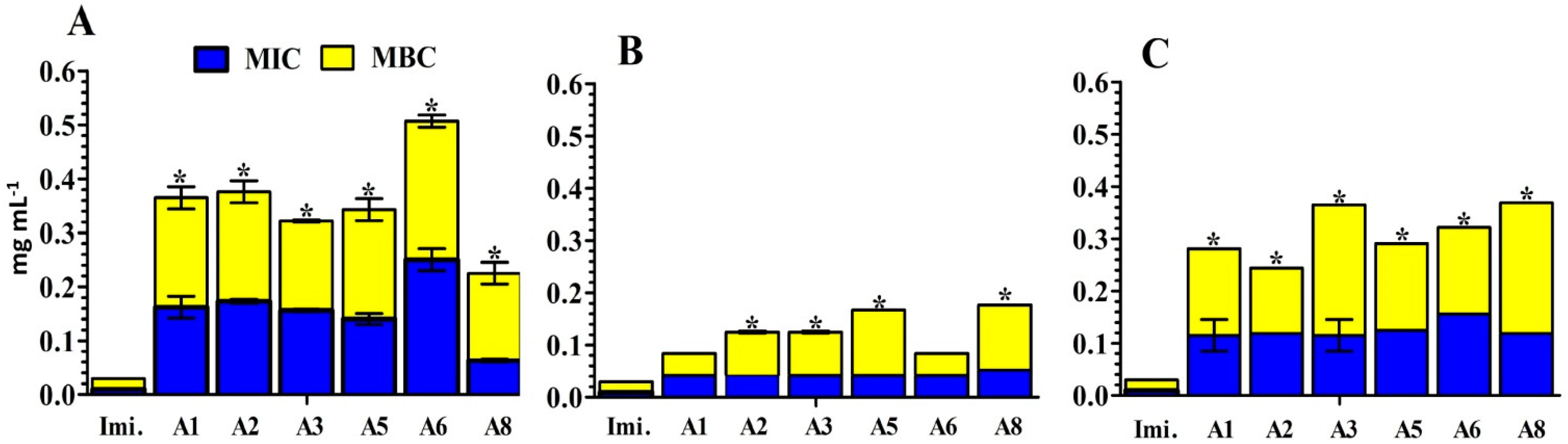
2.4. In Vitro Assay for the Antibacterial Activity of S. aromaticum Extracts
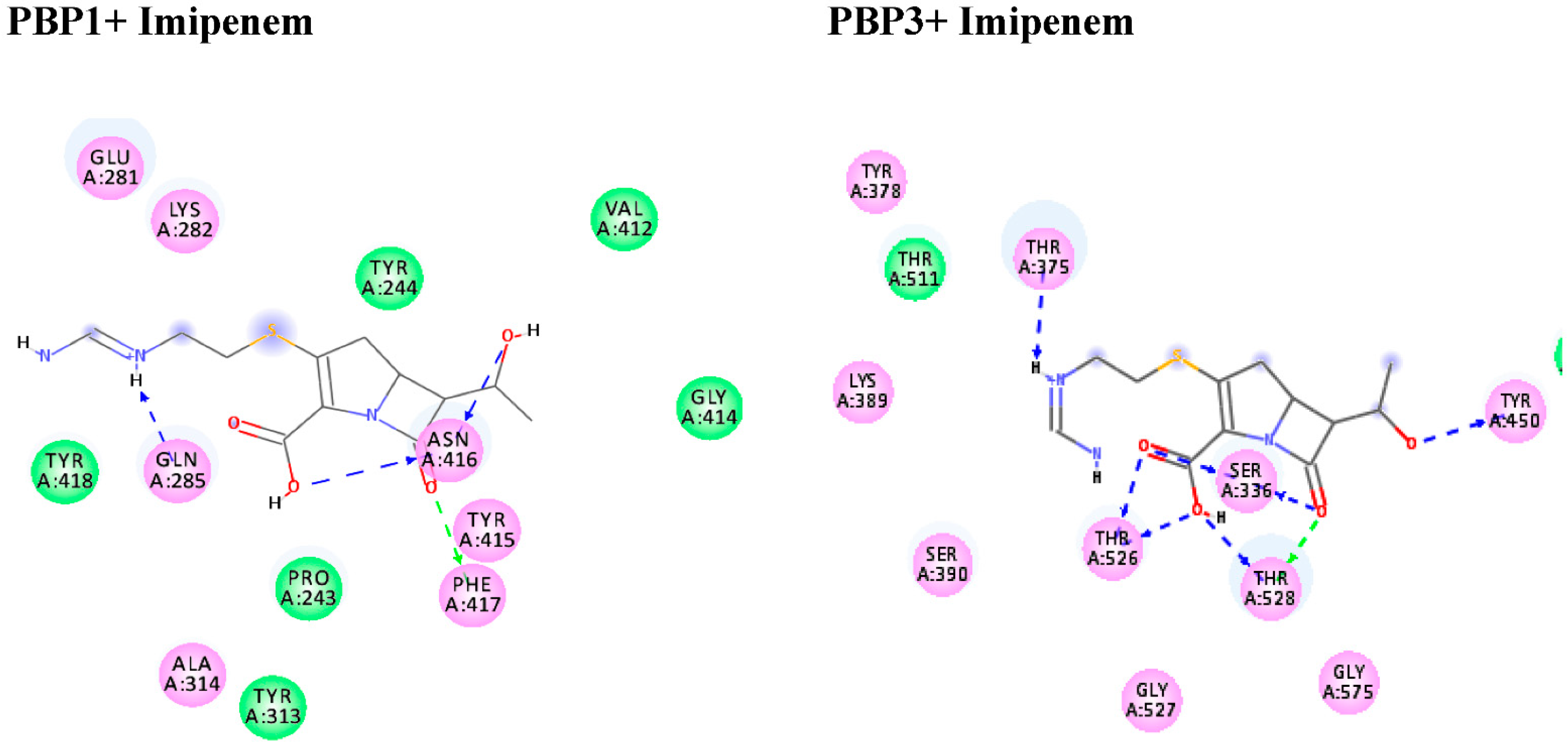
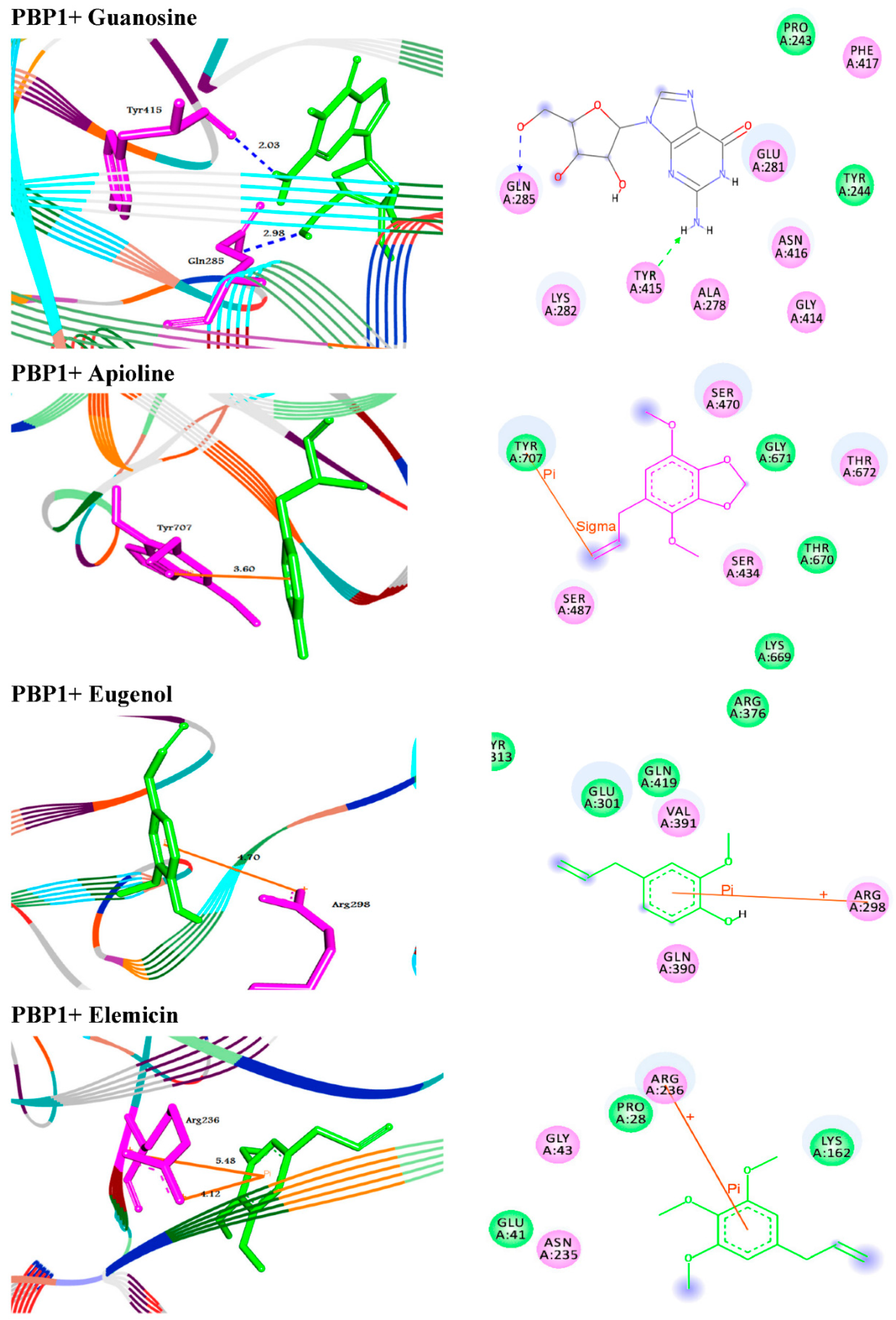
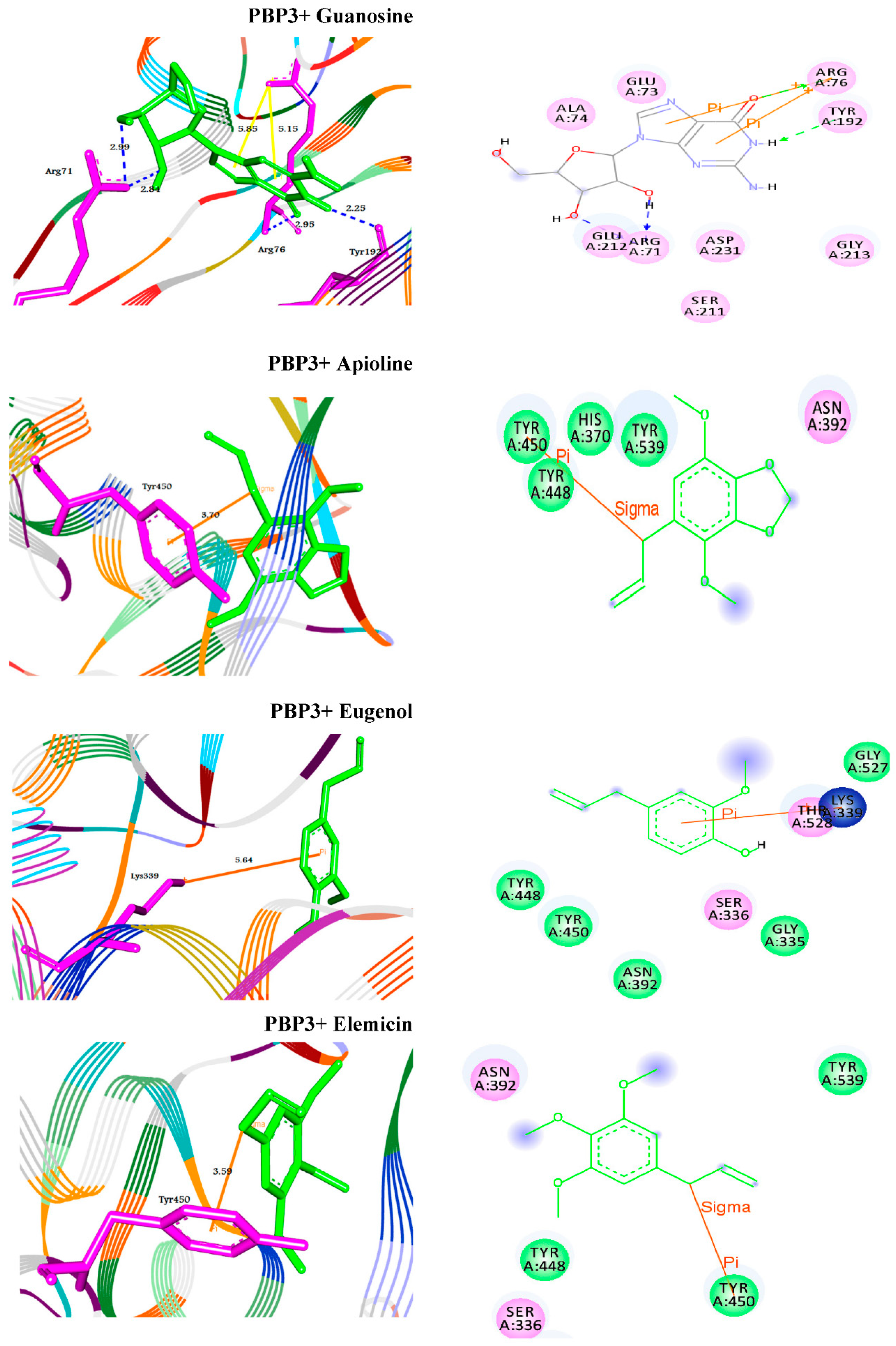
2.5. Molecular Docking Studies of Standard Antibiotic and Herbal Ligands
3. Discussion
4. Materials and Methods
4.1. Sampling Collection, Isolation, and Processing
4.2. Phenotypic Identification of the Isolates by Vitek-2 Systems
4.3. Antimicrobial Susceptibility Testing
4.4. Recognition of Virulence and Antibiotic-Resistant Genes of A. baumannii Isolates
4.5. DNA Amplification for the Selected Virulence and Antibiotic Resistance Genes of Isolates
4.6. Plant Material and Extraction
4.7. Determination of the Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) by INT Reduction Assay
4.8. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
4.9. Molecular Docking Studies of Standard Antibiotic and Herbal Ligands
4.10. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Algammal, A.M.; Hetta, H.F.; Batiha, G.E.; Hozzein, W.N.; El Kazzaz, W.M.; Hashem, H.R.; Tawfik, A.M.; El-Tarabili, R.M. Virulence-determinants and antibiotic-resistance genes of MDR- E. coli isolated from secondary infections following FMD-outbreak in cattle. Sci. Rep. 2020, 10, 19779. [Google Scholar] [CrossRef] [PubMed]
- Abouelfetouh, A.; Torky, A.S.; Aboulmagd, E. Phenotypic and genotypic characterization of carbapenem-resistant Acinetobacter baumannii isolates from Egypt. Antimicrob. Resist. Infect. Control 2019, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Pritsch, M.; Zeynudin, A.; Messerer, M.; Baumer, S.; Liegl, G.; Schubert, S.; Löscher, T.; Hoelscher, M.; Belachew, T.; Rachow, A.; et al. First report on bla NDM-1-producing Acinetobacter baumannii in three clinical isolates from Ethiopia. BMC Infect. Dis. 2017, 17, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. BioMed Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravarty, B. Genetic mechanisms of antibiotic resistance and virulence in Acinetobacter baumannii: Background, challenges and future prospects. Mol. Biol. Rep. 2020, 47, 4037–4046. [Google Scholar] [CrossRef] [PubMed]
- Motbainor, H.; Bereded, F.; Mulu, W. Multi-drug resistance of blood stream, urinary tract and surgical site nosocomial infections of Acinetobacter baumannii and Pseudomonas aeruginosa among patients hospitalized at Felegehiwot referral hospital, Northwest Ethiopia: A cross-sectional study. BMC Infect. Dis. 2020, 20, 92. [Google Scholar] [CrossRef] [Green Version]
- Cerezales, M.; Xanthopoulou, K.; Wille, J.; Krut, O.; Seifert, H.; Gallego, L.; Higgins, P.G. Mobile genetic elements harboring antibiotic resistance determinants in Acinetobacter baumannii isolates from Bolivia. Front. Microbiol. 2020, 11, 919. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.T.; Carmeli, Y.; Falagas, M.T.; Giske, C.T.; Harbarth, S.; Hindler, J.T.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Ghaima, K.K. Distribution of extended spectrum beta-lactamase (ESBL) genes among Acinetobacter baumannii isolated from burn infections. MOJ Cell Sci. Rep. 2018, 5, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Al-Agamy, M.H.; Khalaf, N.G.; Tawfick, M.M.; Shibl, A.M.; El Kholy, A. Molecular characterization of carbapenem-insensitive Acinetobacter baumannii in Egypt. Int. J. Infect. Dis. 2014, 22, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Beriş, F.Ş.; Budak, E.E.; Gülek, D.; Uzun, A.; Cizmeci, Z.; Mengeloğlu, F.Z.; Direkel, Ş.; Cetinkol, Y.; Altıntop, A.; Iraz, M.; et al. Investigation of the frequency and distribution of beta-lactamase genes in the clinical isolates of Acinetobacter baumannii collected from different regions of Turkey: A multicenter study. Mikrobiyoloji Bul. 2016, 50, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Smiline, A.S.G.; Vijayashree, J.P.; Paramasivam, A. Molecular characterization of plasmid-encoded blaTEM, blaSHV and blaCTX-M among extended spectrum β-lactamases [ESBLs] producing Acinetobacter baumannii. Br. J. Biomed. Sci. 2018, 75, 200–202. [Google Scholar] [CrossRef] [PubMed]
- Salih, T.; Salih, H.A. In Silico Design and Molecular Docking Studies of Carbapenem Analogues Targeting Acinetobacter baumannii PBP1A Receptor. Al Mustansiriyah J. Pharm. Sci. 2020, 20, 35–50. [Google Scholar]
- Skariyachan, S.; Manjunath, M.; Bachappanavar, N. Screening of potential lead molecules against prioritised targets of multi-drug-resistant-Acinetobacter baumannii–insights from molecular docking, molecular dynamic simulations and in vitro assays. J. Biomol. Struct. Dyn. 2019, 37, 1146–1169. [Google Scholar] [CrossRef]
- Penwell, W.F.; Shapiro, A.B.; Giacobbe, R.A.; Gu, R.F.; Gao, N.; Thresher, J.; McLaughlin, R.E.; Huband, M.D.; DeJonge, B.L.; Ehmann, D.E.; et al. Molecular mechanisms of sulbactam antibacterial activity and resistance determinants in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2015, 59, 1680–1689. [Google Scholar] [CrossRef] [Green Version]
- Cayô, R.; Rodríguez, M.C.; Espinal, P.; Fernández-Cuenca, F.; Ocampo-Sosa, A.A.; Pascual, Á.; Ayala, J.A.; Vila, J.; Martínez-Martínez, L. Analysis of genes encoding penicillin-binding proteins in clinical isolates of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 5907–5913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leungtongkam, U.; Thummeepak, R.; Kitti, T.; Tasanapak, K.; Wongwigkarn, J.; Styles, K.M.; Wellington, E.M.; Millard, A.D.; Sagona, A.P.; Sitthisak, S. Genomic analysis reveals high virulence and antibiotic resistance amongst phage susceptible Acinetobacter baumannii. Sci. Rep. 2020, 10, 16154. [Google Scholar] [CrossRef] [PubMed]
- Elamary, R.; Salem, W.M. Optimizing and purifying extracellular amylase from soil bacteria to inhibit clinical biofilm-forming bacteria. PeerJ 2020, 8, e10288. [Google Scholar] [CrossRef] [PubMed]
- Yassien, A.S.; Hassan, M.M.; Elamary, R.B. Prevalence of lipase producer Aspergillus niger in nuts and anti-biofilm efficacy of its crude lipase against some human pathogenic bacteria. Sci. Rep. 2021, 11, 7981. [Google Scholar] [CrossRef]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [Green Version]
- Batiha, G.E.S.; Alkazmi, L.M.; Wasef, L.G.; Beshbishy, A.M.; Nadwa, E.H.; Rashwan, E.K. Syzygium aromaticum L. (Myrtaceae): Traditional uses, bioactive chemical constituents, pharmacological and toxicological activities. Biomolecules 2020, 10, 2. [Google Scholar]
- Selles, S.M.A.; Kouidri, M.; Belhamiti, B.T.; Ait Amrane, A. Chemical composition, in-vitro antibacterial and antioxidant activities of Syzygium aromaticum essential oil. J. Food Meas. Charact. 2020, 14, 2352–2358. [Google Scholar] [CrossRef]
- Singh, R.; Lawrence, R.; Lawrence, K.; Agarwal, B.; Gupta, R.K.; Dar, S. Antioxidant and antibacterial activity of Syzigium aromaticum. Chem. Sci. Trans. 2015, 4, 239–245. [Google Scholar]
- Singh, J.; Baghotia, A.; Goel, S.P. Eugenia caryophyllata Thunberg (Family Myrtaceae): A review. Int. J. Res. Pharm. Biomed. Sci. 2012, 3, 1469–1475. [Google Scholar]
- Exner, M.; Bhattacharya, S.; Christiansen, B.; Gebel, J.; Goroncy-Bermes, P.; Hartemann, P.; Heeg, P.; Ilschner, C.; Kramer, A.; Larson, E.; et al. Antibiotic resistance: What is so special about multidrug-resistant Gram-negative bacteria? GMS Hyg. Infect. Control 2017, 12, Doc05. [Google Scholar] [CrossRef]
- Thomson, C.; Armitage, A. Urinary tract infection. In Oxford Textbook of Medicine; Warrell, D.A., Cox, T.M., Firth, J.D., Eds.; Oxford Uni Press: Oxford, UK, 2010; pp. 4103–4122. [Google Scholar]
- Elamary, R.B.; Albarakaty, F.M.; Salem, W.M. Efficacy of Acacia nilotica aqueous extract in treating biofilm-forming and multidrug resistant uropathogens isolated from patients with UTI syndrome. Sci. Rep. 2020, 10, 11125. [Google Scholar] [CrossRef]
- Abdulzahra, A.T.; Khalil, M.A.; Elkhatib, W.F. First report of colistin resistance among carbapenem-resistant Acinetobacter baumannii isolates recovered from hospitalized patients in Egypt. New Microbes New Infect. 2018, 26, 53–58. [Google Scholar] [CrossRef]
- Younis, R.I.; Nasef, S.A.; Salem, W.M. Detection of Multi-Drug Resistant Food-borne Bacteria in Ready-to-Eat Meat Products in Luxor City, Egypt. SVU-Int. J. Vet. Sci. 2019, 2, 20–35. [Google Scholar] [CrossRef] [Green Version]
- Eijkelkamp, B.A.; Stroeher, U.H.; Hassan, K.A.; Paulsen, I.T.; Brown, M.H. Comparative analysis of surface-exposed virulence factors of Acinetobacter baumannii. BMC Genom. 2014, 15, 1020. [Google Scholar] [CrossRef] [Green Version]
- Eraç, B.; Yılmaz, F.F.; Hoşgör Limoncu, M.; Oztürk, I.; Aydemir, S. Investigation of the virulence factors of multidrug-resistant Acinetobacter baumannii isolates. Mikrobiyoloji Bul. 2014, 48, 70–81. [Google Scholar] [CrossRef]
- Al-Kadmy, I.M.S.; Ali, A.N.M.; Salman, I.M.A.; Khazaal, S.S. Molecular characterization of Acinetobacter baumannii isolated from Iraqi hospital environment. New Microbes New Infect. 2018, 21, 51–57. [Google Scholar] [CrossRef]
- Moradi, J.; Hashemi, F.B.; Bahador, A. Antibiotic resistance of Acinetobacter baumannii in Iran: A systemic review of the published literature. Osong Public Health Res. Perspect. 2015, 6, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Tahir, H.U.; Sarfraz, R.A.; Ashraf, A.; Adil, S. Chemical composition and antidiabetic activity of essential oils obtained from two spices (Syzygium aromaticum and Cuminum cyminum). Int. J. Food Prop. 2016, 19, 2156–2164. [Google Scholar] [CrossRef]
- Mercier, B.; Prost, J.; Prost, M. The essential oil of turpentine and its major volatile fraction (α-and β-pinenes): A review. Int. J. Occup. Med. Environ. Health 2009, 22, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, R.E.; Parks, L.W.; Spence, K.D. Some effects of Douglas fir terpenes on certain microorganisms. Appl. Environ. Microbiol. 1980, 40, 301–304. [Google Scholar] [CrossRef] [Green Version]
- Hamad, A.; Mahardika, M.G.P.; Yuliani, I.; Hartanti, D. Chemical constituents and antimicrobial activities of essential oils of Syzygium polyanthum and Syzygium aromaticum. Rasayan J. Chem. 2017, 10, 564–569. [Google Scholar]
- Moo, C.L.; Yang, S.K.; Osman, M.A.; Yuswan, M.H.; Loh, J.Y.; Lim, W.M.; Lim, S.H.E.; Lai, K.S. Antibacterial Activity and Mode of Action of β-caryophyllene on Bacillus cereus. Pol. J. Microbiol. 2020, 69, 49. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.J.; Yoo, J.S.; Lee, T.G.; Cho, H.Y.; Kim, Y.H.; Kim, W.G. Fatty acid synthesis is a target for antibacterial activity of unsaturated fatty acids. FEBS Lett. 2005, 579, 5157–5162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Azeredo, G.A.; Stamford, T.L.M.; Nunes, P.C.; Neto, N.J.G.; De Oliveira, M.E.G.; De Souza, E.L. Combined application of essential oils from Origanum vulgare L. and Rosmarinus officinalis L. to inhibit bacteria and autochthonous microflora associated with minimally processed vegetables. Food Res. Int. 2011, 44, 1541–1548. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Zhou, L.; Huang, Y.; Wang, Y.; Hao, X.; Wang, J. Antimicrobial activity of globulol isolated from the fruits of Eucalyptus globulus Labill. Nat. Prod. Res. 2008, 22, 569–575. [Google Scholar] [CrossRef]
- Jimoh, S.O.; Arowolo, L.A.; Alabi, K.A. Phytochemical screening and antimicrobial evaluation of Syzygium aromaticum extract and essential oil. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 4557–4567. [Google Scholar] [CrossRef]
- Hu, J.; Hu, Q.; He, X.; Liu, C.; Kong, Y.; Cheng, Y.; Zhang, Y. Stimuli--responsive hydrogels with antibacterial activity assembled from guanosine, aminoglycoside, and a bifunctional anchor. Adv. Healthc. Mater. 2020, 9, 1901329. [Google Scholar] [CrossRef] [PubMed]
- El-Saghier, A.M.; El-Naggar, M.; Hussein, A.H.M.; El-Adasy, A.-B.A.; Olish, M.; Abdelmonsef, A.H. Eco-Friendly Synthesis, Biological Evaluation, and In Silico Molecular Docking Approach of Some New Quinoline Derivatives as Potential Antioxidant and Antibacterial Agents. Front. Chem. 2021, 9, 679967. [Google Scholar] [CrossRef] [PubMed]
- Gomha, S.M.; Abdelhady, H.A.; Hassain, D.Z.; Abdelmonsef, A.H.; El-Naggar, M.; Elaasser, M.M.; Mahmoud, H.K. Thiazole-Based Thiosemicarbazones: Synthesis, Cytotoxicity Evaluation and Molecular Docking Study. Drug Des. Dev. Ther. 2021, 15, 659–677. [Google Scholar] [CrossRef]
- Horchani, M.; Hajlaoui, A.; Harrath, A.H.; Mansour, L.; Jannet, H.B.; Romdhane, A. New pyrazolo-triazolo-pyrimidine derivatives as antibacterial agents: Design and synthesis, molecular docking and DFT studies. J. Mol. Struct. 2020, 1199, 127007. [Google Scholar] [CrossRef]
- Sharma, V.; Chitranshi, N.; Agarwal, A.K. Significance and biological importance of pyrimidine in the microbial world. Int. J. Med. Chem. 2014, 2014, 202784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, N.S.; Lindberg, R.B.; Mason, A.D., Jr.; Pruitt, B.A., Jr. The quantitative swab culture and smear: A quick, simple method for determining the number of viable aerobic bacteria on open wounds. J. Trauma 1976, 16, 89–94. [Google Scholar] [CrossRef]
- Joyanes, P.; del Carmen Conejo, M.; Martínez-Martínez, L.; Perea, E.J. Evaluation of the VITEK 2 system for the identification and susceptibility testing of three species of non-fermenting gram-negative rods frequently isolated from clinical samples. J. Clin. Microbiol. 2001, 39, 3247–3253. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standard Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI supplement M100; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Reller, L.B.; Weinstein, M.; Jorgensen, J.H.; Ferraro, M.J. Antimicrobial susceptibility testing: A review of general principles and contemporary practices. Clin. Infect. Dis. 2009, 49, 1749–1755. [Google Scholar]
- Prakash, D.; Saxena, R.S. Antimicrobial susceptibility pattern of human pathogenic bacteria related to Enterobacteriaceae family causing urinary tract infection. Adv. Appl. Sci. Res. 2013, 4, 98–104. [Google Scholar]
- Chiang, M.C.; Kuo, S.C.; Chen, Y.C.; Lee, Y.T.; Chen, T.L.; Fung, C.P. Polymerase chain reaction assay for the detection of Acinetobacter baumannii in endotracheal aspirates from patients in the intensive care unit. J. Microbiol. Immunol. Infect. 2011, 44, 106–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaguchi, K.; Ogitani, T.; Osawa, R.; Kawano, M.; Kokumai, N.; Kaneshige, T.; Noro, T.; Masubuchi, K.; Shimizu, Y. Virulence factors of avian pathogenic Escherichia coli strains isolated from chickens with coli septicemia in Japan. Avian Dis. 2007, 51, 656–662. [Google Scholar] [CrossRef]
- Colom, K.; Pérez, J.; Alonso, R.; Fernández-Aranguiz, A.; Lariño, E.; Cisterna, R. Simple and reliable multiplex PCR assay for detection of blaTEM, blaSHV and blaOXA–1 genes in Enterobacteriaceae. FEMS Microbiol. Lett. 2003, 223, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, O.; Mohamed, H.; Salem, W.; Afifi, M.; Song, Y. Efficacy of Ethanolic Extract of Syzygium aromaticum in the Treatment of Multidrug-Resistant Pseudomonas aeruginosa Clinical Isolates Associated with Urinary Tract Infections. Evid.-Based Complement. Altern. Med. 2021, 2021, 6612058. [Google Scholar] [CrossRef]
- Salem, W.; Shibat El-hamed, D.; Sayed, W.; Elamary, R. Alterations in virulence and antibiotic-resistant genes of multidrug-resistant Salmonella serovars isolated from poultry: The bactericidal efficacy of Allium sativum. Microb. Pathog. 2017, 108, 91–100. [Google Scholar] [CrossRef]
- Eloff, J.N.A. Sensitive and quick microplate method to determine the minimal inhibitory concentration of plant extracts for bacteria. Planta Med. 1998, 64, 711–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lall, N.; Henley-Smith, C.J.; De Canha, M.N.; Oosthuizen, C.B.; Berrington, D. Viability reagent, PrestoBlue, in comparison with other available reagents, utilized in cytotoxicity and antimicrobial assays. Int. J. Microbiol. 2013, 2013, 420601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The protein data bank. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002, 58, 899–907. [Google Scholar] [CrossRef]
- Haredi Abdelmonsef, A.; Eldeeb Mohamed, M.; El-Naggar, M.; Temairk, H.; Mohamed Mosallam, A. Novel Quinazolin-2,4-Dione Hybrid Molecules as Possible Inhibitors Against Malaria: Synthesis and in silico Molecular Docking Studies. Front. Mol. Biosci. 2020, 7, 105. [Google Scholar] [CrossRef]
- Rashdan, H.; Shehadi, I.; Abdelmonsef, A.H. Synthesis, Anticancer Evaluation, Computer-Aided Docking Studies, and ADMET Prediction of 1,2,3-Triazolyl-Pyridine Hybrids as Human Aurora B Kinase Inhibitors. ACS Omega 2021, 6, 1445–1455. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An Open chemical toolbox. J. Cheminformatics 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallakyan, S.; Olson, A.J. Small-Molecule Library Screening by Docking with PyRx. In Chemical Biology; Springer: Berlin/Heidelberg, Germany, 2015; Volume 1263, pp. 243–250. ISBN 9780123944474. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Female | Male | Patient Character | |||
|---|---|---|---|---|---|
| 56.88% | No = 63 | 43.12% | No = 47 | ||
| 28.5 | 18 | 12.77 | 6 | below (30) | Age (year) |
| 60.3 | 38 | 38.30 | 18 | between (30–60) | |
| 11.2 | 7 | 48.93 | 23 | above (60) | |
| Prevalence of underlying disease | |||||
| 20.63 | 9 | 6.38 | 3 | Respiratory disease. | |
| 16.1 | 7 | 10.63 | 5 | Diabetes mellitus. | |
| 27.0 | 17 | 21.3 | 10 | Diabetic and hypertension | |
| 15.87 | 10 | 10.63 | 5 | Diabetic and respiratory disease | |
| 11.11 | 7 | 17.02 | 8 | Diabetic (respiratory disease and hypertension) | |
| 9.51 | 6 | 10.63 | 5 | Patients with elevated liver function test. | |
| 11.1 | 7 | 19.16 | 9 | Patients with elevated kidney function test. | |
| 0.0 | 0 | 4.26 | 2 | Catheter presence | |
| Types of Samples (Isolate No.) | Virulence Genes | Antibiotic Resistance Genes | ||
|---|---|---|---|---|
| recA | iutA | blaTEM | BlaSHV | |
| Urine (A1) | 1 | 1 | 1 | 1 |
| Wound (A5) | 1 | 1 | 1 | 1 |
| Sputum (A2,A3,A6, A8 *) | 4 | 4 | 4 | 3 |
| Total | 6 | 6 | 6 | 5 |
| Aqueous Extracts | |||||
|---|---|---|---|---|---|
| No. | RT (min) | Compound Name | M. Formula | M.wt | Area (%) |
| 1 | 7.38 | Limonene | C10H16 | 136 | 1.56 |
| 2 | 8.61 | α-Pinene | C10H16 | 136 | 18.82 |
| 3 | 11.26 | (2E)-3,7-dimethylocta-2,6-dienal | C10H16O | 152 | 0.86 |
| 4 | 13.18 | Camphor | C10H16O | 152 | 11.75 |
| 5 | 14.33 | Cyclododecene | C12H22 | 166 | 0.76 |
| 6 | 14.80 | 2,4 Decadienal | C10H16O | 152 | 1.05 |
| 7 | 18.06 | α -Chamigrene | C15H24 | 204 | 0.52 |
| 8 | 18.21 | à-Guaiene | C15H24 | 204 | 0.59 |
| 9 | 18.57 | Beta-caryophyllene | C15H24 | 204 | 15.12 |
| 10 | 19.32 | Ethyl benzoylacetate | C11H12O3 | 192 | 1.05 |
| 11 | 19.52 | Globulol | C15H26O | 222 | 11.35 |
| 12 | 22.76 | Apioline | C12H14O4 | 222 | 5.45 |
| 13 | 29.05 | Hexadecanoic acid | C16H32O2 | 256 | 4.59 |
| 14 | 29.78 | Loganetin | C11H16O5 | 228 | 8.51 |
| 15 | 30.75 | Isobergapten | C12H8O4 | 216 | 1.09 |
| 16 | 32.66 | Oleic acid | C18H34O2 | 282 | 14.52 |
| 17 | 32.86 | Isochiapin B | C20H26N2O2 | 326 | 0.62 |
| 18 | 42.01 | Lucenin | C27H30O16 | 610 | 0.56 |
| Ethanolic extract | |||||
| 1 | 28.33 | Pentenenitrile | C5H7N | 81 | 4.37 |
| 2 | 29.66 | Ethyl oleate | C20H38O2 | 310 | 2.54 |
| 3 | 32.17 | cis-10-Nonadecenoic acid | C19H36O2 | 296 | 5.37 |
| 4 | 35.12 | Indole | C8H7N | 117 | 6.83 |
| 5 | 37.21 | Guanosine | C10H13N5O5 | 283 | 8.91 |
| 6 | 39.55 | Oleic acid | C18H34O2 | 282 | 27.22 |
| 7 | 39.75 | Chlorozotocin | C9H16ClN3O7 | 313 | 0.52 |
| 8 | 40.62 | 1-Eicosene | C20H40 | 280 | 6.30 |
| 9 | 46.77 | Nonadecene | C19H38 | 266 | 1.20 |
| 10 | 48.85 | 3-Hexacosanol | C26H54O | 382 | 2.21 |
| 11 | 51.96 | Nonacosane | C29H60 | 408 | 0.36 |
| 12 | 53.70 | Dodecanoic acid | C12H24O2 | 200 | 2.34 |
| Ethyl acetate extract | |||||
| 1 | 13.23 | Linalool | C10H18O | 154 | 1.24 |
| 2 | 13.94 | Carveol | C10H16O | 152 | 8.08 |
| 3 | 14.46 | Citral | C10H16O | 152 | 13.48 |
| 4 | 15.07 | Eugenol | C10H12O2 | 164 | 1.91 |
| 5 | 15.23 | 1-Hexadecene | C16H32 | 224 | 0.16 |
| 6 | 18.30 | Phenol, 2,4-bis(1,1-dimethylethyl)- | C14H22O | 206 | 0.13 |
| 7 | 20.62 | Elemicin | C12H16O3 | 208 | 3.32 |
| 8 | 21.73 | Farnesyl acetate | C17H28O2 | 264 | 1.44 |
| 9 | 22.42 | Heptadecane | C17H36 | 240 | 0.28 |
| 10 | 23.94 | Tetradecanoic acid | C14H28O2 | 228 | 0.73 |
| 11 | 27.11 | Hexadecanoic acid, methyl ester | C17H34O2 | 270 | 0.12 |
| 12 | 27.71 | Alantolactone | C15H20O2 | 232 | 6.36 |
| 13 | 28.16 | Hexadecanoic acid | C16H32O2 | 256 | 11.95 |
| 14 | 28.56 | Eremanthin | C15H18O2 | 230 | 2.76 |
| 15 | 30.32 | Linoleic acid methyl ester | C19H34O2 | 294 | 0.73 |
| 16 | 30.43 | Oleic acid methyl ester | C19H36O2 | 296 | 0.37 |
| 17 | 31.60 | Linoleic acid | C18H32O2 | 280 | 36.16 |
| 18 | 31.71 | Oleic Acid | C18H34O2 | 282 | 6.39 |
| 19 | 31.95 | Octadecanoic acid | C18H36O2 | 284 | 3.23 |
| 20 | 43.71 | Ethyl iso-allocholate | C26H44O5 | 436 | 0.22 |
| PBP1 (3udx) | PBP3 (3ue3) | |||||
|---|---|---|---|---|---|---|
| Binding Energy kcal/mol | Docked Complex (Amino Acid–Ligand) | Distance (Å) | Binding Energy kcal/mol | Docked Complex (Amino Acid–Ligand) | Distance (Å) | |
| Imipenem | −6.8 | H–bond | −6.5 | H–bond | ||
| GLN285:NE2–Imipenem | 2.15 | SER336–Imipenem | 1.94 | |||
| ASN416–Imipenem | 2.55 | SER336–Imipenem | 2.17 | |||
| PHE417–Imipenem | 2.96 | TYR450–Imipenem | 2.23 | |||
| THR526–Imipenem | 2.18 | |||||
| THR526–Imipenem | 1.99 | |||||
| THR528–Imipenem | 2.53 | |||||
| THR528–Imipenem | 2.97 | |||||
| Guanosine | −7.1 | H–bond | −8.1 | H–bond | ||
| GLN285:NE2–Guanosine | 2.98 | ARG71:NH1–Guanosine | 2.97 | |||
| TYR415:O–Guanosine | ARG71:NH1–Guanosine | 2.84 | ||||
| 2.03 | ARG76:N–Guanosine | 2.93 | ||||
| TYR192:O–Guanosine | 2.25 | |||||
| π–π interaction | ||||||
| ARG76:N–Guanosine | 4.14 | |||||
| ARG76:N–Guanosine | 5.06 | |||||
| Apioline | −5.6 | π–sigma interaction | −6.0 | π–sigma interaction | ||
| TYR707–Apioline | 3.60 | TYR450–Apioline | 3.70 | |||
| Eugenol | −5.4 | π–cation interaction | −5.8 | π–cation interaction | ||
| ARG298:NH2–Eugenol | 4.70 | LYS339:NZ–Eugenol | 5.64 | |||
| Elemicin | −5.3 | π–cation interaction | −5.2 | π–sigma interaction | ||
| ARG236:NH1–Elemicin | 5.48 | TYR450–Elemicin | 3.59 | |||
| ARG236:NH2–Elemicin | 4.12 | |||||
| Target Gene | Sequence | Amplified Segment (bp) | Primary Denaturation | Amplification (35 Cycles) | References | |||
|---|---|---|---|---|---|---|---|---|
| Secondary Denaturation | Annealing | Extension | Final Extension | |||||
| Virulence genes used for Acinetobacter baumannii isolates * | ||||||||
| recA | CCTGAATCTTCTGGTAAAAC GTTTCTGGGCTGCCAAACATTAC | 425 | 94 °C/5 min | 94 °C/30 s | 50 °C/45 s | 72 °C/30 s | 72 °C/10 min | [53] |
| iutA | GGCTGGACATGGGAACTGG CGTCGGGAACGGGTAGAATCG | 300 | 94 °C/5 min | 94 °C/30 s | 63 °C/30 s | 72 °C/45 s | 72 °C/7 min | [54] |
| Antibiotics resistance genes | ||||||||
| blaTEM | ATCAGCAATAAACCAGC CCCCGAAGAACGTTTTC | 516 | 94 °C/5 min | 94 °C/30 s | 54 °C/40 s | 72 °C/45 s | 72 °C/10 min | [55] |
| blaSHV | AGGATTGACTGCCTTTTTG ATTTGCTGATTTCGCTCG | 392 | 94 °C/5 min | 94 °C/30 s | 54 °C/40 s | 72 °C/45 s | 72 °C/10 min | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoud, A.; Afifi, M.M.; El Shenawy, F.; Salem, W.; Elesawy, B.H. Syzygium aromaticum Extracts as a Potential Antibacterial Inhibitors against Clinical Isolates of Acinetobacter baumannii: An In-Silico-Supported In-Vitro Study. Antibiotics 2021, 10, 1062. https://doi.org/10.3390/antibiotics10091062
Mahmoud A, Afifi MM, El Shenawy F, Salem W, Elesawy BH. Syzygium aromaticum Extracts as a Potential Antibacterial Inhibitors against Clinical Isolates of Acinetobacter baumannii: An In-Silico-Supported In-Vitro Study. Antibiotics. 2021; 10(9):1062. https://doi.org/10.3390/antibiotics10091062
Chicago/Turabian StyleMahmoud, Abdelhamed, Magdy M. Afifi, Fareed El Shenawy, Wesam Salem, and Basem H. Elesawy. 2021. "Syzygium aromaticum Extracts as a Potential Antibacterial Inhibitors against Clinical Isolates of Acinetobacter baumannii: An In-Silico-Supported In-Vitro Study" Antibiotics 10, no. 9: 1062. https://doi.org/10.3390/antibiotics10091062
APA StyleMahmoud, A., Afifi, M. M., El Shenawy, F., Salem, W., & Elesawy, B. H. (2021). Syzygium aromaticum Extracts as a Potential Antibacterial Inhibitors against Clinical Isolates of Acinetobacter baumannii: An In-Silico-Supported In-Vitro Study. Antibiotics, 10(9), 1062. https://doi.org/10.3390/antibiotics10091062