
Antifungal Nanoformulation for Biocontrol of Tomato Root and Crown Rot Caused by Fusarium oxysporum f. sp. radicis-lycopersici

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtaining and Identifying the Isolates
2.2. Nanoemulsion Preparation
In Vitro Evaluation of Plant Extracts
2.3. Tomato Plant Trials
2.4. Statistical Analysis
3. Results
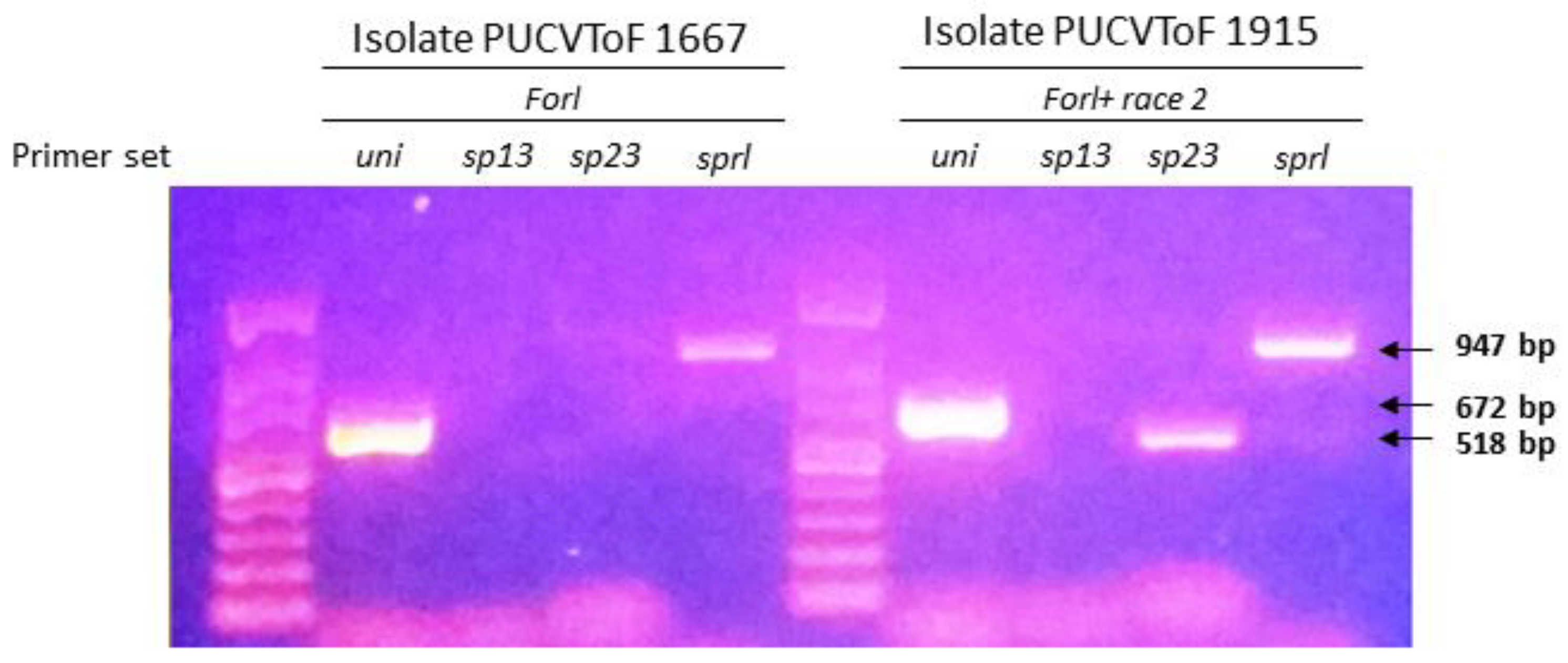
3.1. Identification of Isolates
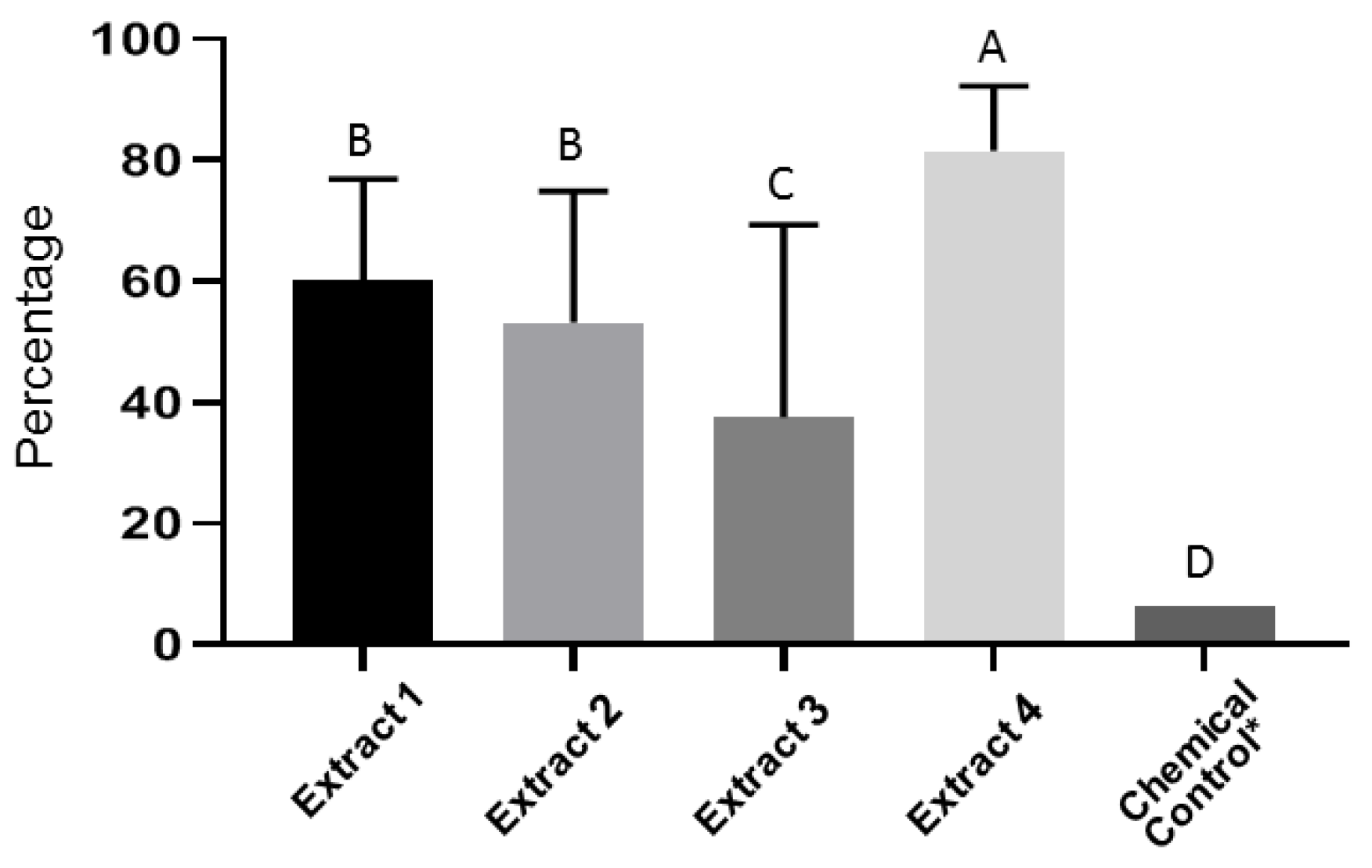
3.2. In Vitro Evaluation of Plant Extracts on Fusarium Oxysporum Spore Germination
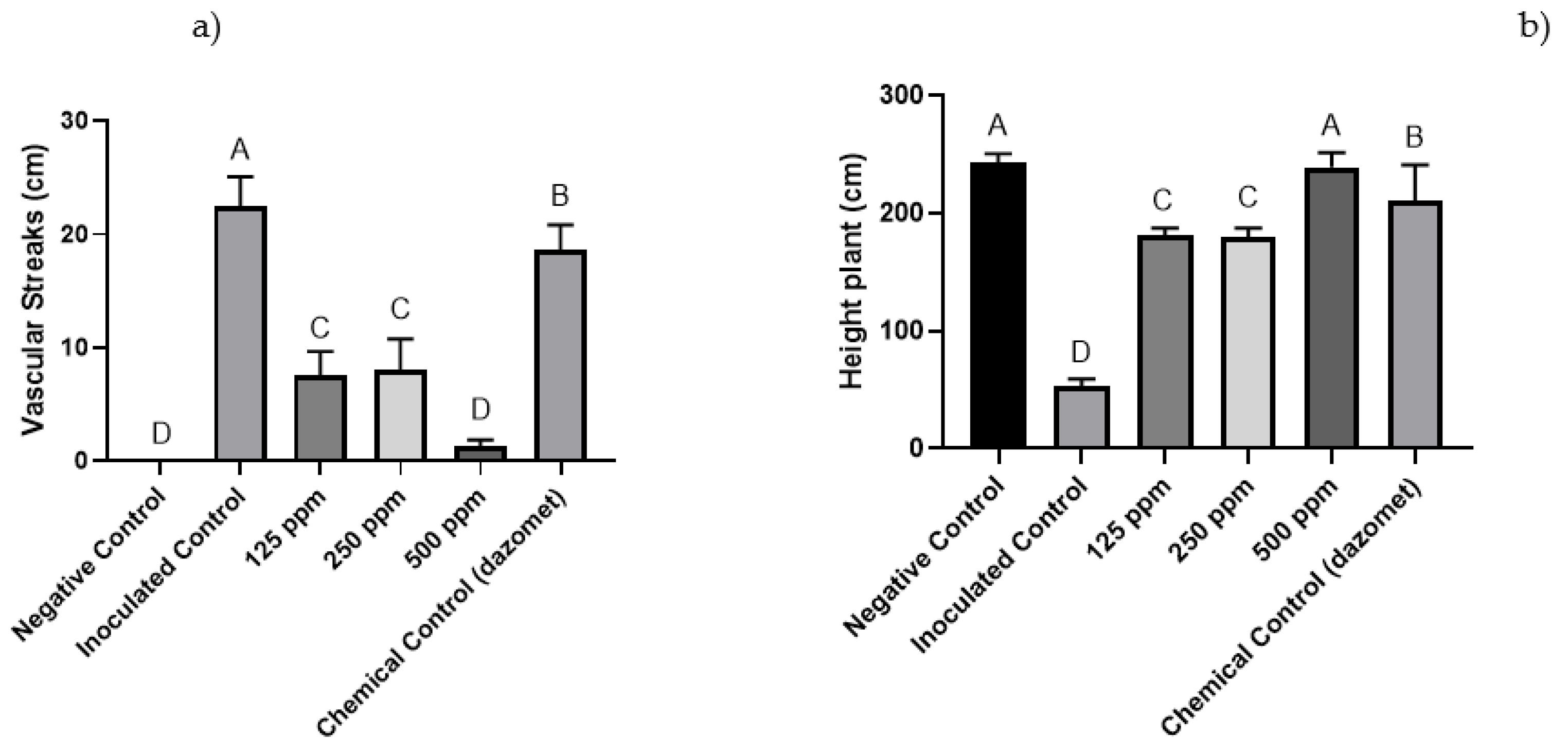
3.3. Greenhouse Trials
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caracterización Económica de la Cadena Agroalimentaria del Tomate de Uso Industrial. Available online: https://www.odepa.gob.cl/wp-content/uploads/2019/02/TomateIndustrial_2018.pdf (accessed on 4 February 2020).
- The World′s Leading Producers of Tomatoes. Available online: https://www.worldatlas.com/articles/which-are-the-world-s-leading-tomato-producing-countries.html (accessed on 22 January 2020).
- Carbonell, C.; Cifuentes, D.; Tello, J.; Cenis, J. Fusarium oxysporum f. sp. lycopersici and F. o. F. sp. radicis-lycopersici and its detection in plant using RAPD markers. Bol. San. Veg. Plagas 1994, 20, 919–926. [Google Scholar]
- Attitalla, H.; Fatehi, J.; Levenfors, J.; Brishammar, S. A rapid molecular method for differentiating two special forms (lycopersici and radicis-lycopersici) of Fusarium oxysporum. Mycol. Res. 2004, 108, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Ozbay, N.; Newman, S. Fusarium crown and root rot of tomato and control methods. Plant Pathol. J. 2004, 3, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Enespa, S.; Dwivedi, S.K. Effectiveness of some Antagonistic fungi and botanicals against Fusarium solani and Fusarium oxysporum f. sp. lycopersici infecting brinjal and tomato plants. Asian J. Plant Pathol. 2014, 8, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Singh, J.; Ray, S.; Rajput, R.S.; Vaishnav, A.; Singh, R.K.; Singh, H.B. Seed biopriming with antagonistic microbes and ascorbic acid induce resistance in tomato against fusarium wilt. Microbiol. Res. 2020, 237, 126482. [Google Scholar] [CrossRef] [PubMed]
- Apodaca, M.; Zavaleta, E.; Osada, S.; García, R.; Valenzuela, J. Hospedantes asintomáticos de Fusarium oxysporum Schlechtend. f. sp. radicis lycopersici W.R. Jarvis y Shoemaker en Sinaloa, México. Rev. Mex. Fitopatol. 2004, 22, 7–13. [Google Scholar]
- Pérez-Leal, R.; Villa-Martinez, A.; Morales, H.; Basurto-Sotelo, M.; Soto-Parra, J.; Martínez-Escudero, E. Current situation of Fusarium spp. in the control and evaluation of the antifungal activity on vegetables extracts. Acta Agron. 2014, 64, 194–205. [Google Scholar]
- Lagopodi, A.; Ram, A.; Lamers, G.; Punt, P.; Van den Hodel, C.; Lugtenberg, B.; Bloemberg, G. Novel aspects of tomato root colonization and infection by Fusarium oxysporum f. sp. radicis-lycopersici revealed by confocal laser scanning microscopic analysis using the green fluorescent protein as a marker. Mol. Plant-Microbe Interact. 2001, 15, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Gastélum, I.; Martínez Téllez, M.; Corrales-Maldonado, C.; Vargas-Arispuro, I.; Arellano-Gil, M. Induction of defense response in tomato plants against Forl by garlic extract. Rev. Mex. Fitopatol. 2018, 36, 394–413. [Google Scholar] [CrossRef]
- Stanghellini, M.; Rasmussen, S. Hydroponics: A solution for zoosporic pathogens. Plant Dis. 1994, 78, 1129–1138. [Google Scholar] [CrossRef]
- Ramyabharathi, S.A.; Meena, B.; Raguchander, T. Induction of chitinase and β-1, 3-glucanase PR proteins in tomato through liquid formulated Bacillus subtilis EPCO 16 against fusarium wilt. J. Today Biol. Sci. Res. Rev. 2012, 1, 50–60. [Google Scholar]
- García, C.; Ruíz, C.; Palmero, D.; de Cara, M.; Díaz, M.; Camacho, F.; Tello, J. Comportamiento de patrones de tomate frente a la patogenicidad de Fusarium oxysporum f. sp. radicis-lycopersici. In Proceedings of the XIII Congreso de ciencias Hortícolas, Almería, Spain, 16–20 April 2012. [Google Scholar]
- Manual de Cultivo del Tomate al Aire Libre. Available online: http://www.inia.cl/wpcontent/uploads/ManualesdeProduccion/11%20Manual%20Tomate%20Aire%20Libre.pdf (accessed on 4 February 2020).
- Herrera, R. Estudio Etiológico de Fusarium Oxysporum en Tomate en la Zona Central de Chile. Ph.D. Thesis, Universidad de Chile, Santiago, Chile, 2017. [Google Scholar]
- Sepúlveda-Chavera, G.; Huanca, W. First report of Fusarium oxysporum f. sp. lycopersici race 3 and F. oxysporum f. sp. radicis-lycopersici in tomatoes in the Azapa valley of Chile. Plant Dis. 2014, 98, 1432. [Google Scholar] [CrossRef] [PubMed]
- Salvador, O.; Staltari, S.; Chorzempa, S.E.; Astiz, M.; Molina, M. Biological control of Fusarium graminearum: Use of Trichoderma spp. and biofumigation with aerial part of Brassica juncea. Rev. Fac. Cienc. Agrar. Univ. Nac. Cuyo. 2014, 46, 45–57. [Google Scholar]
- Van den Bosch, F.; Paveley, N.; Van den Berg, F.; Hobbelen, P.; Oliver, R. Mixtures as a Fungicide resistance management tactic. Phytopathology 2014, 104, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- McGovern, R. Management of tomato diseases caused by Fusarium oxysporum. Crop Prot. 2015, 73, 78–92. [Google Scholar] [CrossRef]
- Louws, F.J.; Suchoff, D.; Kressin, J.; Panthee, D.; Driver, J.; Gunter, C. Integrating grafting and emerging products to manage soilborne diseases of tomato. Acta Hort. 2018, 1207, 249–254. [Google Scholar] [CrossRef]
- Kim, J.; Jiypung, K. Inhibitory effect of algal extracts on mycelial growth of the tomato-wilt pathogen, Fusarium oxysporum f. sp. lycopersici. Mycobiology 2008, 36, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Avendaño, C.; Arbeláez, G.; Rondón, G. Biological control of Phaseolus vulgaris bean vascular wilt caused by Fusarium oxysporum f. sp. phaseoli with combined Entrophospora colombiana, Trichoderma sp. and Pseudomonas fluorescens action. Rev. Agron. Colom. 2006, 24, 62–67. [Google Scholar]
- Montenegro, I.; Madrid, A.; Cuellar, M.; Seeger, M.; Alfaro, J.; Besoain, X.; Martínez, J.; Ramirez, I.; Olguín, Y.; Valenzuela, M. Biopesticide activity from drimanic compounds to control tomato pathogens. Molecules 2018, 23, 2053. [Google Scholar] [CrossRef] [Green Version]
- Alzate, D.; Mier, G.; Afanador, L.; Durango, D.; García, C. Evaluation of phytotoxicity and antifungal activity against Colletotrichum acutatum of essential oils of thyme (Thymus vulgaris), lemongrass (Cymbopogon citratus), and its main constituents. Rev. Qui. Farm. 2008, 16, 116–125. [Google Scholar]
- Alanazi, S.; Alnoman, M.; Banawas, S.; Saito, R.; Sarker, M.R. The inhibitory effects of essential oil constituents against germination, outgrowth and vegetative growth of spores of Clostridium perfringens type A in laboratory medium and chicken meat. Food Microbiol. 2018, 73, 311–318. [Google Scholar] [CrossRef]
- López-Benítez, A.; López-Betancourt, S.; Vázquez-Badillo, M.; Rodríguez-Herrera, S. Inhibición del Crecimiento Micelial de Fusarium oxysporum Schlechtend. f. sp. lycopersici (Sacc.) Snyder y Hansen, Rhizoctonia solani Kühn y Verticilllium dahliae Kleb. Mediante Extractos Vegetales Acuosos. Rev. Mex. Fitopatol. 2005, 23, 183. [Google Scholar]
- Pal, K.K.; McSpadden, B.G. Biological Control of Plant Pathogens. Plant Health Instr. 2006, 5. [Google Scholar] [CrossRef] [Green Version]
- Alamri, S.; Hashem, M.; Mostafa, Y.S.; Nafady, N.; Abo-Elyours, K. Biological control of root rot in lettuce caused by Exserohilum rostratum and Fusarium oxysporum via induction of the defense mechanism. Biol. Control 2019, 128, 76–84. [Google Scholar] [CrossRef]
- Shen, Z.; Xue, C.; Penton, C.R.; Thomashow, L.; Zhang, N.; Wang, B.; Ruan, Y.; Li, R.; Shen, Q. Suppression of banana Panama disease induced by soil microbiome reconstruction through an integrated agricultural strategy. Soil Biol. Biochem. 2019, 128, 164–174. [Google Scholar] [CrossRef]
- Gogoi, R.; Dureja, P.; Singh, P.K. Nanoformulations-A safer and effective option for agrochemicals. Indian Farming 2009, 59, 7–12. [Google Scholar]
- Sharma, A.; Sharma, N.K.; Srivastava, A.; Kataria, A.; Dubey, S.; Sharma, S.; Kundu, B. Clove and lemongrass oil based non-ionic nanoemulsión for suppressing the growth of plant pathogenic Fusarium oxysporum f. sp. lycopersici. Ind. Crops Prod. 2018, 123, 353–362. [Google Scholar] [CrossRef]
- Santos-Ordoñez, N.; Reyes-Munguía, A.; Hernández-Fuentes, A.D.; Almaraz-Buendía, I.; Campos-Montiel, R.G.; Cenobio-Galindo, A. Efecto de compuestos bioactivos nanoencapsulados contra hongos de interés agroalimentario. Inv. Des. Cien. Tec. Alim. 2019, 4, 118–124. [Google Scholar]
- Park, J.Y.; Kim, S.H.; Kim, N.H.; Lee, S.W.; Jeun, Y.C.; Hong, J.K. Differential inhibitory activities of four plant essential oils on in vitro growth of Fusarium oxysporum f. sp. Fragariae causing fusarium wilt in strawberry plants. Plant Pathol. J. 2017, 33, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elsalam, K.A.; Khokhlov, A.R. Eugenol oil nanoemulsion: Antifungal activity against Fusarium oxysporum f. sp. vasinfectum and phytotoxicity on cottonseeds. Appl. Nanosci. 2015, 5, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Nash, S.M.; Snyder, W.C. Quantitative estimations by plate counts of propagules of the bean root rot Fusarium in field soils. Phytopathology 1962, 52, 567–572. [Google Scholar]
- Hirano, Y.; Arie, T. PCR-based differentiation of Fusarium oxysporum f. sp. lycopersici and radicis-lycopersici and races of F. oxysporum f. sp. lycopersici. J. Gen. Plant Pathol. 2006, 72, 273–283. [Google Scholar] [CrossRef]
- Sonneveld, C. Fertigation in the greenhouse industry. In Proceedings of the Dahlia Greidinger International Symposium on Fertigation, Technion Institute of Technology, Haifa, Israel, 25 March–1 April 1995. [Google Scholar]
- Paulus, O. Fusarium crown and root rot. In Compendium of Tomato Diseases; Jones, J.B., Jones, J.P., Stall, R.E., Zitter, T.A., Eds.; APS Press: Eagan, MN, USA, 1997; p. 14. [Google Scholar]
- Bautista, S.; Hernández, M.L.; Bosquez, E.M.; Wilson, C.L. Effects of chitosan and plant extracts on growth of Colletotrichum gloeosporioides, anthracnose levels and quality of papaya fruti. Crop Prot. 2003, 22, 1087–1092. [Google Scholar] [CrossRef]
- Abou-Jawdah, Y.; Sobh, H.; Salameh, A. Antimycotic activities of selected plant flora, growing wild in Lebanon, against phytopathogenic fungi. J. Agric. Food Chem. 2002, 50, 3208–3213. [Google Scholar] [CrossRef] [PubMed]
- Kliebenstein, D.J. Secondary metabolites and plant/environment interactions: A view through Arabidopsis thaliana tinged glasses. Plant Cell Environ. 2004, 27, 675–684. [Google Scholar] [CrossRef]
- Jasso de Rodríguez, D.; Hernández-Castillo, D.; Angulo-Sánchez, J.L.; Rodríguez-García, R.; Villarreal-Quintanilla, J.A.; Lira-Saldivar, R.H. Antifungal activity in vitro of Flourensia spp. Extracts on Alternaria sp., Rhizoctonia solani, and Fusarium oxysporum. Ind. Crops Prod. 2007, 25, 111–116. [Google Scholar] [CrossRef]
- Bowers, J.; Locke, J.C. Effect of botanical extracts on the population density of Fusarium oxysporum in soil and control of fusarium wilt in the greenhouse. Plant Dis. 2000, 84, 300–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Set | Uni | Sp13 | Sp23 | Sprl | Result |
|---|---|---|---|---|---|
| PUCVToF 1667 | + 1 | − | − | + | Forl |
| PUCVToF 1915 | + | − | + | + | Forl + race 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aravena, R.; Besoain, X.; Riquelme, N.; Salinas, A.; Valenzuela, M.; Oyanedel, E.; Barros, W.; Olguin, Y.; Madrid, A.; Alvear, M.; et al. Antifungal Nanoformulation for Biocontrol of Tomato Root and Crown Rot Caused by Fusarium oxysporum f. sp. radicis-lycopersici. Antibiotics 2021, 10, 1132. https://doi.org/10.3390/antibiotics10091132
Aravena R, Besoain X, Riquelme N, Salinas A, Valenzuela M, Oyanedel E, Barros W, Olguin Y, Madrid A, Alvear M, et al. Antifungal Nanoformulation for Biocontrol of Tomato Root and Crown Rot Caused by Fusarium oxysporum f. sp. radicis-lycopersici. Antibiotics. 2021; 10(9):1132. https://doi.org/10.3390/antibiotics10091132
Chicago/Turabian StyleAravena, Ricardo, Ximena Besoain, Natalia Riquelme, Aldo Salinas, Miryam Valenzuela, Eduardo Oyanedel, Wilson Barros, Yusser Olguin, Alejandro Madrid, Matias Alvear, and et al. 2021. "Antifungal Nanoformulation for Biocontrol of Tomato Root and Crown Rot Caused by Fusarium oxysporum f. sp. radicis-lycopersici" Antibiotics 10, no. 9: 1132. https://doi.org/10.3390/antibiotics10091132
APA StyleAravena, R., Besoain, X., Riquelme, N., Salinas, A., Valenzuela, M., Oyanedel, E., Barros, W., Olguin, Y., Madrid, A., Alvear, M., & Montenegro, I. (2021). Antifungal Nanoformulation for Biocontrol of Tomato Root and Crown Rot Caused by Fusarium oxysporum f. sp. radicis-lycopersici. Antibiotics, 10(9), 1132. https://doi.org/10.3390/antibiotics10091132