
Combinations of Peptide-Protein Extracts from Native Probiotics Suppress the Growth of Multidrug-Resistant Staphylococcus aureus and Citrobacter freundii via Membrane Perturbation and Ultrastructural Changes


Abstract
:1. Introduction
2. Results and Discussion
2.1. Several PCs Inhibit Both Target S. aureus ATCC1026 and C. freundii UTNB3Sm1 Microorganisms
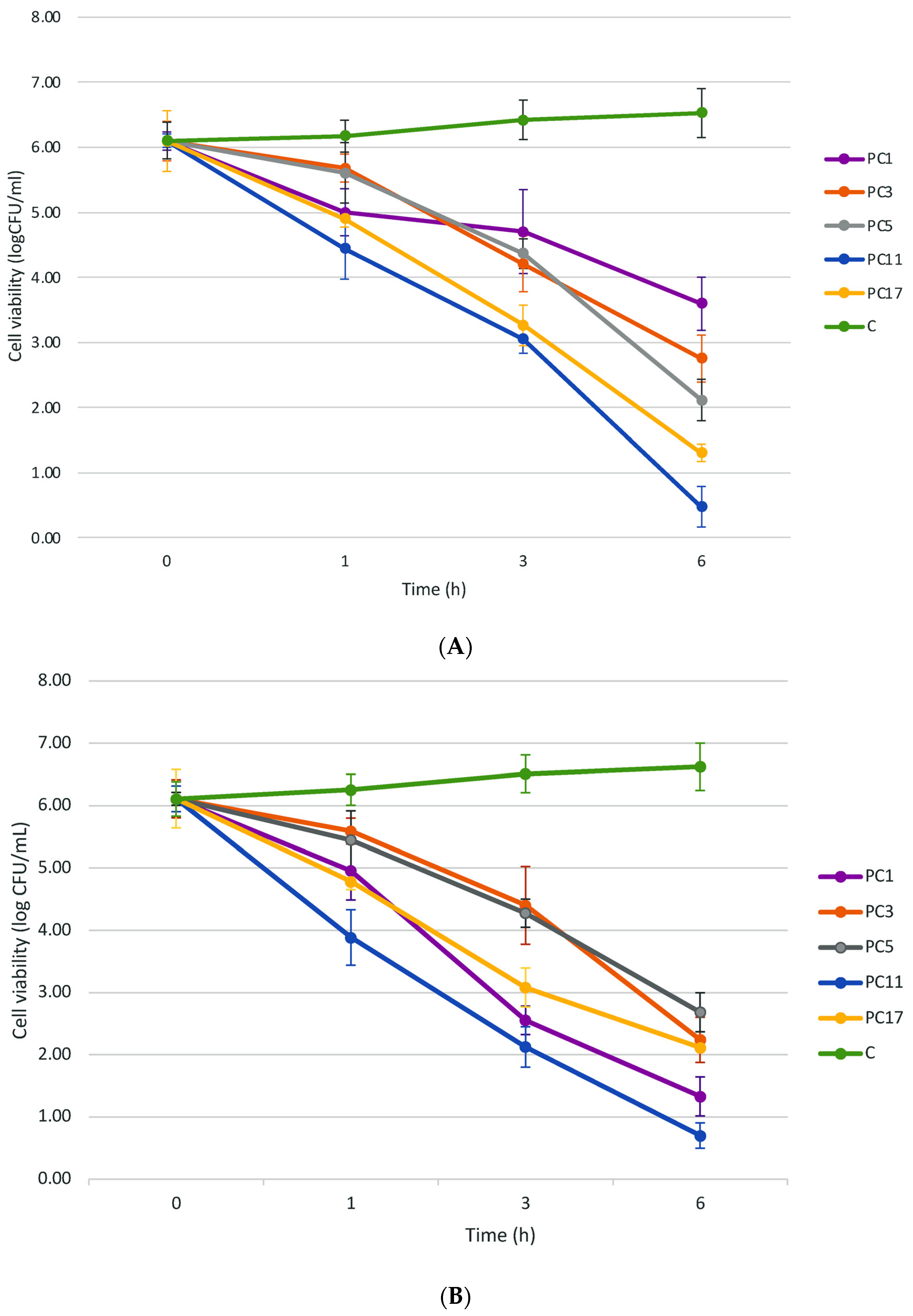
2.2. Overloading of the Peptide-Protein Extracts from UTNGt2 and/or UTNGt21A but Not UTNGt28 Strains Reduced the Target Cell Viability In Vitro
2.3. PC Treatment Compromised the Cell Membrane Integrity of Target Bacteria
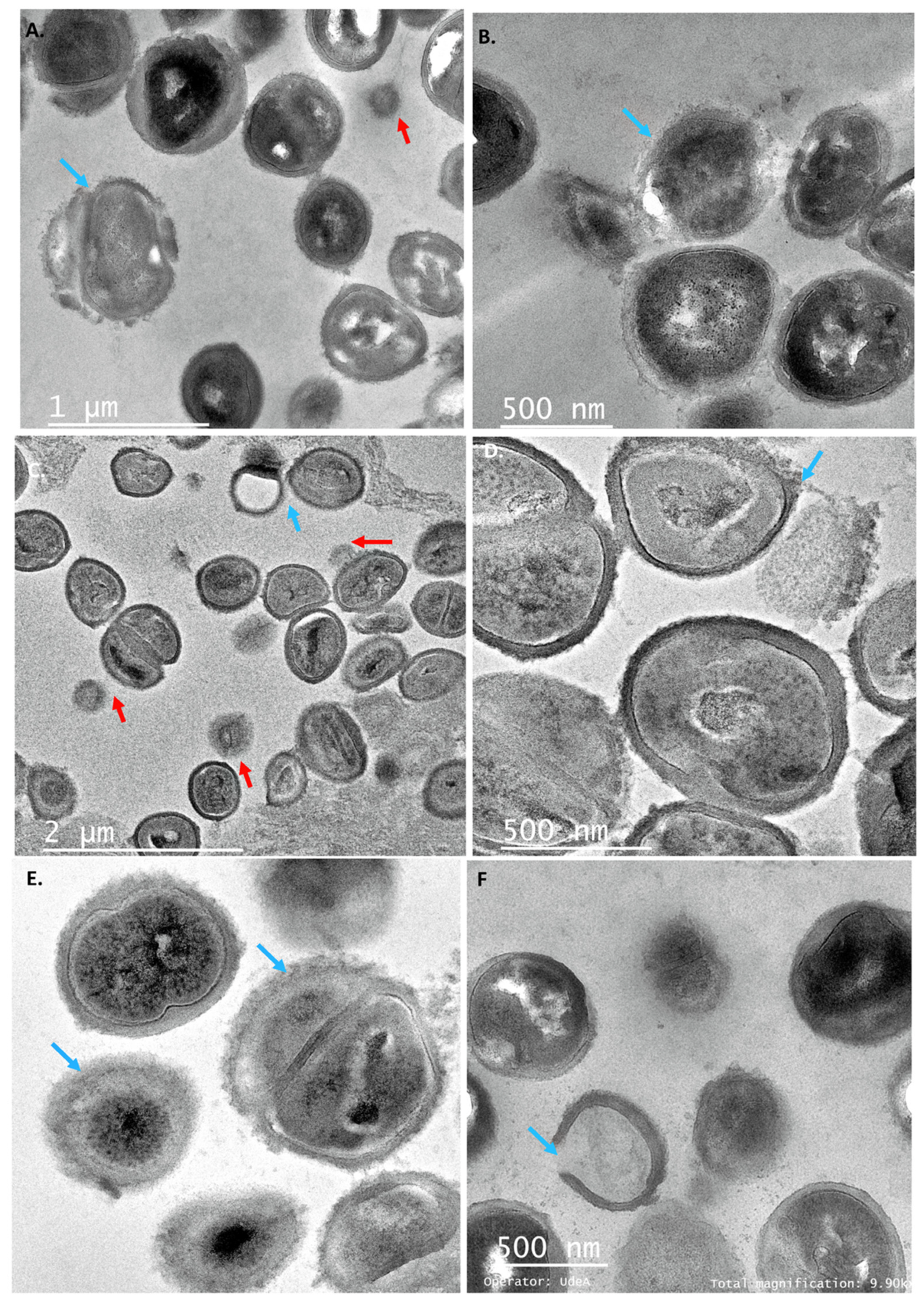
2.4. Individual PCs Induced Several Membrane Morphological Changes of the Target Strains
2.5. SEM Analysis Revealed Different Cell Shape Changes of Both Target Strains upon PC Treatment
3. Materials and Methods
3.1. Microorganisms
3.2. Peptide-Protein Extracts Preparation
3.3. Establish the Combination of the Peptide-Protein Extracts (PCs) and Antimicrobial Activity Evaluation
3.4. Time-Killing Assay of the Selected PCs
3.5. Leakage of Aromatic Molecules Assay
3.6. Transmission Electron Microscope (TEM) Examination of Target Cells Treated with PCs
3.7. Scanning Electron Microscope (SEM) Examination of Target Cells Treated with PCs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, Y.; Bai, J.; Zhong, K.; Huang, Y.; Qi, H.; Jiang, Y.; Gao, H. Antibacterial activity and membrane-disruptive mechanism of 3-p-trans-coumaroyl-2-hydroxyquinic acid, a novel phenolic compound from pine needles of Cedrus deodara, against Staphylococcus aureus. Molecules 2016, 21, 1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozemeijer, W.; Fink, P.; Rojas, E.; Jones, C.H.; Pavliakova, D.; Giardina, P.; Murphy, E.; Liberator, P.; Jiang, Q.; Girgenti, D. Evaluation of approaches to monitor Staphylococcus aureus virulence factor expression during human disease. PLoS ONE 2015, 10, e0116945. [Google Scholar] [CrossRef] [PubMed]
- Jeyanthy, V.; Velusamy, P.; Kumar, G.V.; Kiruba, K. Effect of naturally isolated hydroquinone in disturbing the cell membrane integrity of Pseudomonas aeruginosa MTCC 741 and Staphylococcus aureus MTCC 740. Heliyon 2021, 7, e07021. [Google Scholar] [CrossRef] [PubMed]
- Kadariya, J.; Smith, T.C.; Thapaliya, D. Staphylococcus aureus and staphylococcal food-borne disease: An ongoing challenge in public health. BioMed Res. Int. 2014, 2014, 827965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aminharati, F.; Ehrampoush, M.H.; Soltan Dallal, M.M.; Yaseri, M.; Dehghani Tafti, A.A.; Rajabi, Z. Citrobacter freundii foodborne disease outbreaks related to environmental conditions in Yazd Province, Iran. Iran. J. Public Health 2019, 48, 1099–1105. [Google Scholar] [CrossRef]
- Liu, L.; Qin, L.; Hao, S.; Lan, R.; Xu, B.; Guo, Y.; Jiang, R.; Sun, H.; Chen, X.; Lv, X.; et al. Lineage, Antimicrobial resistance and virulence of Citrobacter spp. Pathogens 2020, 9, 195. [Google Scholar] [CrossRef] [Green Version]
- Joaquin, A.; Khan, S.; Russel, N.; al Fayez, N. Neonatal meningitis and bilateral cerebellar abscesses due to Citrobacter freundii. Pediatr. Neurosurg. 1991, 17, 23–24. [Google Scholar] [CrossRef]
- Tschape, H.; Prager, R.; Streckel, W.; Fruth, A.; Tietze, E.; Böhme, G. Verotoxinogenic Citrobacter freundii associated with severe gastroenteritis and cases of haemolytic uraemic syndrome in a nursery school: Green butter as the infection source. Epidemiol. Infect. 1995, 114, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Xia, S.; Lan, R.; Liu, L.; Ye, C.; Wang, Y.; Jin, N.; Cui, Z.; Jing, H.; Xiong, Y.; et al. Isolation and characterization of cytotoxic, aggregative Citrobacter freundii. PLoS ONE 2012, 7, e33054. [Google Scholar] [CrossRef] [Green Version]
- Pletz, M.W.; Wollny, A.; Dobermann, U.-H.; Rödel, J.; Neubauer, S.; Stein, C.; Brandt, C.; Hartung, A.; Mellmann, A.; Trommer, S.; et al. A nosocomial foodborne outbreak of a VIM carbapenemase-expressing Citrobacter freundii. Clin. Infect. Dis. 2018, 67, 58–64. [Google Scholar] [CrossRef]
- Liu, L.; Lan, R.; Liu, L.; Wang, Y.; Zhang, Y.; Wang, Y.; Xu, J. Antimicrobial resistance and cytotoxicity of Citrobacter spp. in Maanshan Anhui Province, China. Front. Microbiol. 2017, 8, 1357. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Xiong, Z.; Li, X.; Hu, L.; Shen, J.; Li, T.; Hu, F.; Chen, S. Prevalence of plasmid-mediated quinolone resistance determinants in Citrobacter freundii isolates from Anhui province, PR China. J. Med. Microbiol. 2011, 60, 1801–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, H.; Pinto, G.; Oliveira, A.; Oliveira, C.; Faustino, M.A.; Briers, Y.; Domingues, L.; Azeredo, J. Characterization and genome sequencing of a Citrobacter freundii phage CfP1 harboring a lysin active against multidrug-resistant isolates. Appl. Microbiol. Biotechnol. 2016, 100, 10543–10553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, H.; Shen, S.; Xu, J.; Lin, S.; Yuan, Y.; Jones, G.S. Synergistic interactions of cinnamaldehyde in combination with carvacrol against food-borne bacteria. Food Control 2013, 34, 619–623. [Google Scholar] [CrossRef]
- Silva, D.R.; Orlandi Sardi, J.C.; Souza Pitangui, N.; Roque, S.M.; da Silva, A.C.B.; Rosalen, P.L. Probiotics as an alternative antimicrobial therapy: Current reality and future directions. J. Funct. Foods 2020, 73, 104080. [Google Scholar] [CrossRef]
- Abdelhamid, A.G.; Esaam, A.; Hazaa, M.M. Cell free preparations of probiotics exerted antibacterial and antibiofilm activities against multidrug resistant E. Coli. Saudi Pharm. J. 2018, 26, 603–607. [Google Scholar] [CrossRef]
- Tumbarski, Y.; Nikolova, R.; Petkova, N.; Ivanov, I.; Lante, A. Biopreservation of fresh strawberries by carboxymethyl cellulose edible coatings enriched with a bacteriocin from Bacillus methylotrophicus BM47. Food Technol. Biotechnol. 2019, 57, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, antimicrobial peptides from bacterial origin: Overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef]
- Pérez-Ramos, A.; Madi-Moussa, D.; Coucheney, F.; Drider, D. Current knowledge of the mode of action and immunity mechanisms of lab-bacteriocins. Microorganisms 2021, 9, 2107. [Google Scholar] [CrossRef]
- Tenea, G.N.; Olmedo, D.; Ortega, C. Peptide-based formulation from lactic acid bacteria impairs the pathogen growth in Ananas comosus (Pineapple). Coatings 2020, 10, 457. [Google Scholar] [CrossRef]
- Tenea, G.N.; Hurtado, P.; Ortega, C. Inhibitory effect of substances produced by native Lactococcus lactis strains of tropical fruits towards food pathogens. Prev. Nutr. Food Sci. 2018, 23, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Tenea, G.N.; Pozo Delgado, T. Antimicrobial peptides from Lactobacillus plantarum UTNGt2 prevent harmful bacteria growth on fresh tomatoes. J. Microbiol. Biotechnol. 2019, 29, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Tenea, G.N.; Ortega, C. Genome Characterization of Lactiplantibacillus plantarum strain UTNGt2 originated from Theobroma grandiflorum (White Cacao) of Ecuadorian Amazon: Antimicrobial peptides from safety to potential applications. Antibiotics 2021, 10, 383. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Zhou, J.; Liu, G.; Chen, F.; Chen, Y.; Gao, X.; Dixon, W.; Song, M.; Xiao, H.; Cao, Y. Membrane disruption and DNA binding of Staphylococcus aureus cell induced by a novel antimicrobial peptide produced by Lactobacillus paracasei subsp. tolerans FX-6. Food Control 2016, 59, 609–613. [Google Scholar] [CrossRef]
- Vidhyasagar, V.; Jeevaratnam, K. Bacteriocin activity against various pathogens produced by Pediococcus pentosaceus VJ13 isolated from Idly batter. Biomed. Chromatogr. 2013, 27, 1497–1502. [Google Scholar] [CrossRef]
- FAO. 21.CFR.172.120. Code of Federal Regulation—Title 21. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr=172.120&SearchTerm=calciumdisodiumedta (accessed on 10 May 2021).
- Li, L.; Shi, Y.; Cheserek, M.J.; Su, G.; Le, G. Antibacterial activity and dual mechanisms of peptide analog derived from cell penetrating peptide against Salmonella typhimurium and Streptococcus pyogenes. Appl. Microbiol. Biotechnol. 2013, 97, 1711–1723. [Google Scholar] [CrossRef]
- Juda, M.; Paprota, K.; Malm, A. EDTA as a potential agent preventing formation of Staphylococcus epidermidis biofilm on polichloride vinyl biomaterials. Ann. Agric. Environ. Med. 2008, 15, 237–241. [Google Scholar]
- Kahl, B.C.; Belling, G.; Reichelt, R.; Herrmann, M.; Proctor, R.A.; Peters, G. Thymidine-dependent small-colony variants of Staphylococcus aureus exhibit gross morphological and ultrastructural changes consistent with impaired cell separation. J. Clin. Microbiol. 2003, 41, 410–413. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, M.; Zhou, C.; Kallenbach, N.R.; Ren, D. Control of bacterial persister cells by Trp/Arg-containing antimicrobial peptides. Appl. Environ. Microbiol. 2011, 77, 4878–4885. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Brul, S.; Zaat, S.A.J. Bacterial persister-cells and spores in the food chain: Their potential inactivation by antimicrobial peptides (AMPs). Int. J. Mol. Sci. 2020, 21, 8967. [Google Scholar] [CrossRef]
- Mandal, S.M.; Pati, B.R.; Chakraborty, R.; Franco, O.L. New insights into the bioactivity of peptides from probiotics. Front. Biosci. Elite 2016, 8, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Selegård, R.; Musa, A.; Nyström, P.; Aili, D.; Bengtsson, T.; Khalaf, H. Plantaricins markedly enhance the effects of traditional antibiotics against Staphylococcus epidermidis. Future Microbiol. 2019, 14, 195–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Farrah, S.; Baney, R.H. Membrane damage of bacteria by silanols treatment. Electron. J. Biotechnol. 2007, 10, 252–259. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef]
- Xue, R.; Liu, Y.; Zhang, Q.; Liang, C.; Qin, H.; Liu, P.; Wang, K.; Zhang, X.; Chen, L.; Wei, Y. Shape changes and interaction mechanism of Escherichia coli cells treated with sericin and use of a sericin-based hydrogel for wound healing. Appl. Environ. Microbiol. 2016, 82, 4663–4672. [Google Scholar] [CrossRef] [Green Version]
- Grigor’eva, A.; Bardasheva, A.; Tupitsyna, A.; Amirkhanov, N.; Tikunova, N.; Pyshnyi, D.; Ryabchikova, E. Changes in the ultrastructure of Staphylococcus aureus treated with cationic peptides and chlorhexidine. Microorganisms 2020, 8, 1991. [Google Scholar] [CrossRef]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Mode of action of the antimicrobial peptide Mel4 is independent of Staphylococcus aureus cell membrane permeability. PLoS ONE 2019, 14, e0215703. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, R.; Chalasani, A.G.; Chattopadhyay, D.; Roy, U. Ultrastructural changes in methicillin-resistant Staphylococcus aureus (MRSA) induced by a novel cyclic peptide ASP-1 from Bacillus subtilis: A scanning electron microscopy (SEM) study. Rev. Argent. Microbiol. 2021, 53, 281–286. [Google Scholar] [CrossRef]
- Cushnie, T.P.; O’Driscoll, N.H.; Lamb, A.J. Morphological and ultrastructural changes in bacterial cells as an indicator of antibacterial mechanism of action. Cell Mol. Life Sci. 2016, 73, 4471–4492. [Google Scholar] [CrossRef]
- Yang, L.; Li, P.; Liang, B.; Hu, X.; Li, J.; Xie, J.; Yang, C.; Hao, R.; Wang, L.; Jia, L.; et al. Multidrug-resistant Citrobacter freundii ST139 co-producing NDM-1 and CMY-152 from China. Sci. Rep. 2018, 8, 10653. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feeds. Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 2740. [Google Scholar] [CrossRef]
- Cheikhyoussef, A.; Pogori, N.; Chen, H.; Tian, F.; Chen, W.; Tang, J.; Zhang, H. Antimicrobial activity and partial characterization of bacteriocin-like inhibitory substances (BLIS) produced by Bifidobacterium infantis BCRC 14602. Food Control 2009, 20, 553–559. [Google Scholar] [CrossRef]
- Ge, J.; Sun, Y.; Xin, X.; Wang, Y.; Ping, W. Purification and partial characterization of a novel bacteriocin synthesized by Lactobacillus paracasei HD1–7 isolated from Chinese sauerkraut juice. Sci. Rep. 2016, 6, 19366. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Combination/Concentration | Inhibition Zone (mm) * | |
|---|---|---|---|
| S. aureus ATCC1026 | C. freundii UTNB3Sm1 | ||
| PC1 | UTNGt2 (1 × MIC) | 9.17 ± 0.29 | 11.03 ± 0.06 |
| PC2 | UTNGt2 (1 × MIC) + EDTA (0.1 mg/mL) | 9.17 ± 0.29 | 9.33 ± 0.58 |
| PC3 | UTNGt28 (1 × MIC) | 9.67 ± 0.58 | 9.17 ± 0.29 |
| PC4 | UTNGt28 (1 × MIC) + EDTA (0.1 mg/mL) | 9.67 ± 0.58 | 9.33 ± 0.58 |
| PC5 | UTNGt21A (1 × MIC) | 9.10 ± 0.17 | 9.33 ± 0.58 |
| PC6 | UTNGt21A (1 × MIC) + EDTA (0.1 mg/mL) | 9.10 ± 0.17 | 9.10 ± 0.17 |
| PC7 | (UTNGt2: UTNGt28) (1 × MIC: 1 × MIC) | 9.10 ± 0.17 | 10.17 ± 0.29 |
| PC8 | (UTNGt2: UTNGt28) (1 × MIC: 1 × MIC) + EDTA (0.1 mg/mL) | 9.33 ± 0.58 | 10.17 ± 0.29 |
| PC9 | (UTNGt2: UTNGt28) (1 × MIC: 3 × MIC) | 9.10 ± 0.17 | 9.67 ± 0.58 |
| PC10 | (UTNGt2: UTNGt28) (1 × MIC: 3 × MIC) + EDTA (0.1 mg/mL) | 9.10 ± 0.17 | 9.10 ± 0.17 |
| PC11 | (UTNGt2: UTNGt28) (3 × MIC: 1 × MIC) | 12.17 ± 0.29 | 13.67 ± 0.58 |
| PC12 | (UTNGt2: UTNGt28) (3 × MIC: 1 × MIC) + EDTA (0.1 mg/mL) | 9.10 ± 0.17 | 10.17 ± 0.29 |
| PC13 | (UTNGt21A: UTNGt28) (1 × MIC: 1 × MIC) | 9.10 ± 0.17 | 9.10 ± 0.17 |
| PC14 | (UTNGt21A: UTNGt28) (1 × MIC: 1 × MIC) + EDTA (0.1 mg/mL) | 9.10 ± 0.17 | 9.10 ± 0.17 |
| PC15 | (UTNGt21A: UTNGt28) (1 × MIC: 3 × MIC) | 9.67 ± 0.58 | 9.10 ± 0.17 |
| PC16 | (UTNGt21A: UTNGt28) (1 × MIC: 3 × MIC) + EDTA (0.1 mg/mL) | 9.67 ± 0.58 | 9.33 ± 0.58 |
| PC17 | (UTNGt21A: UTNGt28) (3 × MIC: 1 × MIC) | 11.67 ± 0.58 | 11.10 ± 0.17 |
| PC18 | (UTNGt21A: UTNGt28) (3 × MIC: 1 × MIC) + EDTA (0.1 mg/mL) | 9.17 ± 0.29 | 9.17 ± 0.29 |
| PC19 | EDTA (0.1 mg/mL) | 7.27 ± 0.64 | 6.10 ± 0.17 |
| MRS broth | Negative control | 6.00 ± 0.00 | 6.00 ± 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tenea, G.N.; Angamarca, E.; Olmedo, D. Combinations of Peptide-Protein Extracts from Native Probiotics Suppress the Growth of Multidrug-Resistant Staphylococcus aureus and Citrobacter freundii via Membrane Perturbation and Ultrastructural Changes. Antibiotics 2022, 11, 154. https://doi.org/10.3390/antibiotics11020154
Tenea GN, Angamarca E, Olmedo D. Combinations of Peptide-Protein Extracts from Native Probiotics Suppress the Growth of Multidrug-Resistant Staphylococcus aureus and Citrobacter freundii via Membrane Perturbation and Ultrastructural Changes. Antibiotics. 2022; 11(2):154. https://doi.org/10.3390/antibiotics11020154
Chicago/Turabian StyleTenea, Gabriela N., Evelyn Angamarca, and Daniela Olmedo. 2022. "Combinations of Peptide-Protein Extracts from Native Probiotics Suppress the Growth of Multidrug-Resistant Staphylococcus aureus and Citrobacter freundii via Membrane Perturbation and Ultrastructural Changes" Antibiotics 11, no. 2: 154. https://doi.org/10.3390/antibiotics11020154
APA StyleTenea, G. N., Angamarca, E., & Olmedo, D. (2022). Combinations of Peptide-Protein Extracts from Native Probiotics Suppress the Growth of Multidrug-Resistant Staphylococcus aureus and Citrobacter freundii via Membrane Perturbation and Ultrastructural Changes. Antibiotics, 11(2), 154. https://doi.org/10.3390/antibiotics11020154