
Antibacterial Activity of the Novel Drug Gepotidacin against Stenotrophomonas maltophilia—An In Vitro and In Vivo Study

 ,
, 
Abstract
:1. Introduction
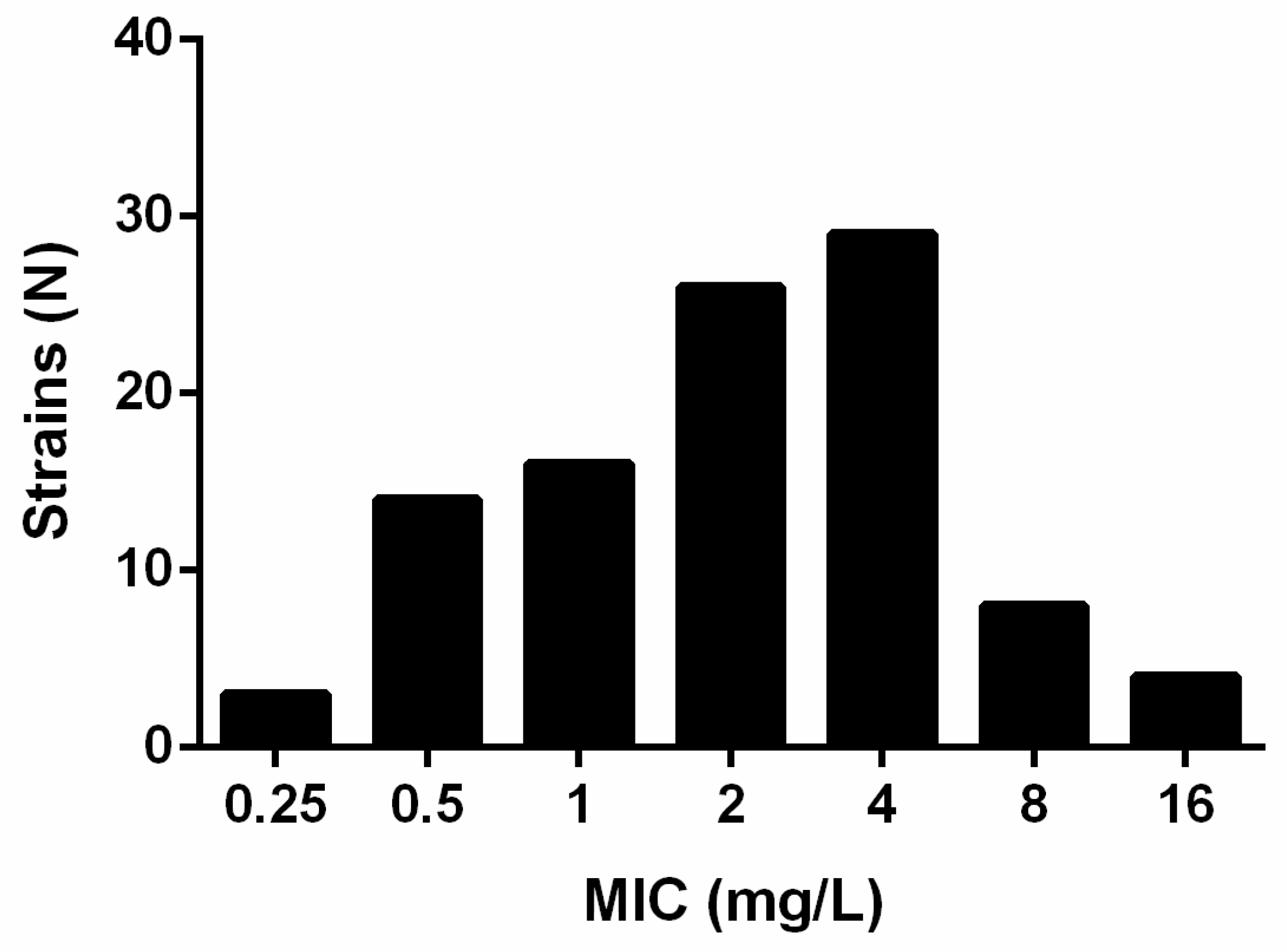
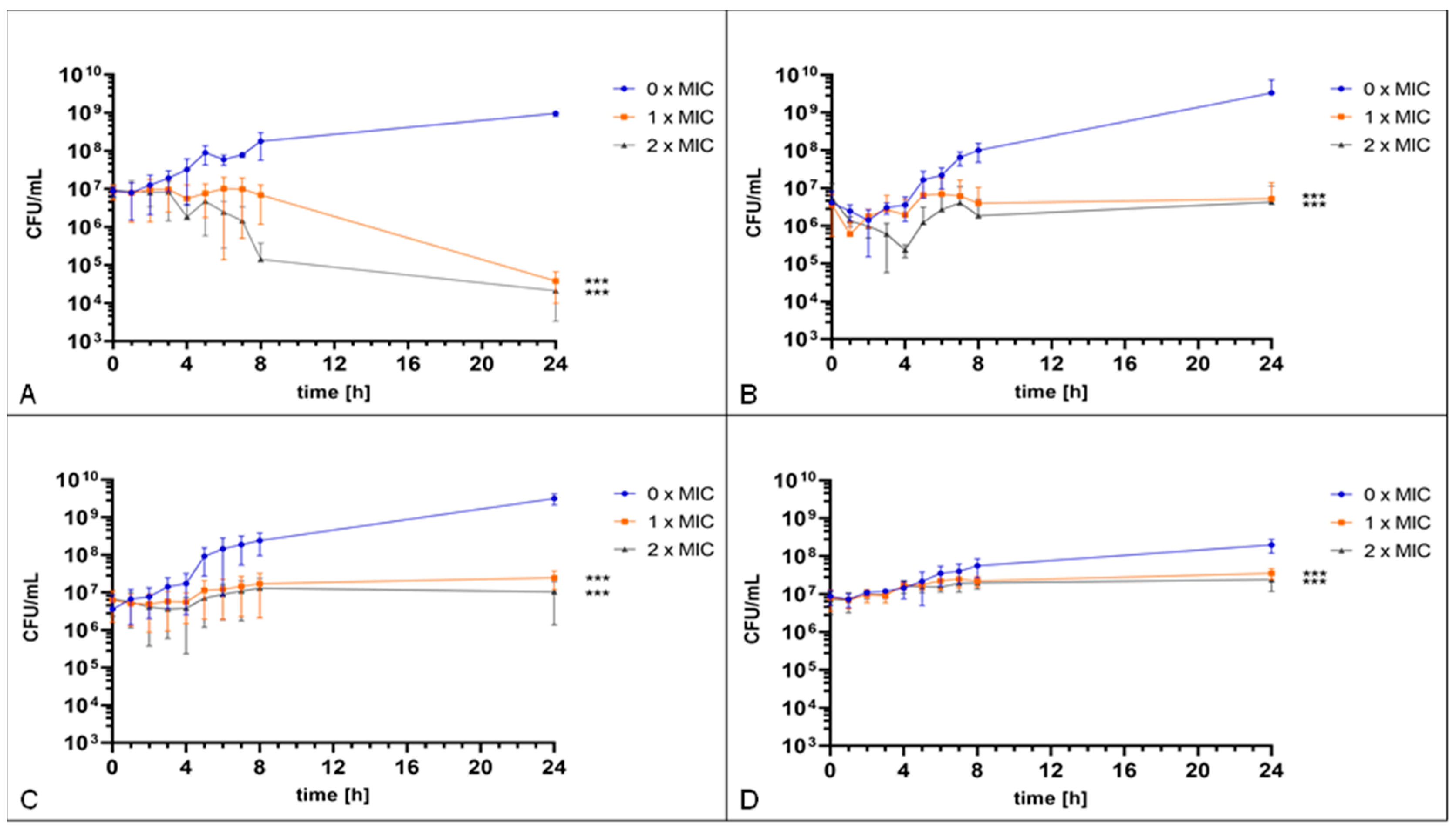
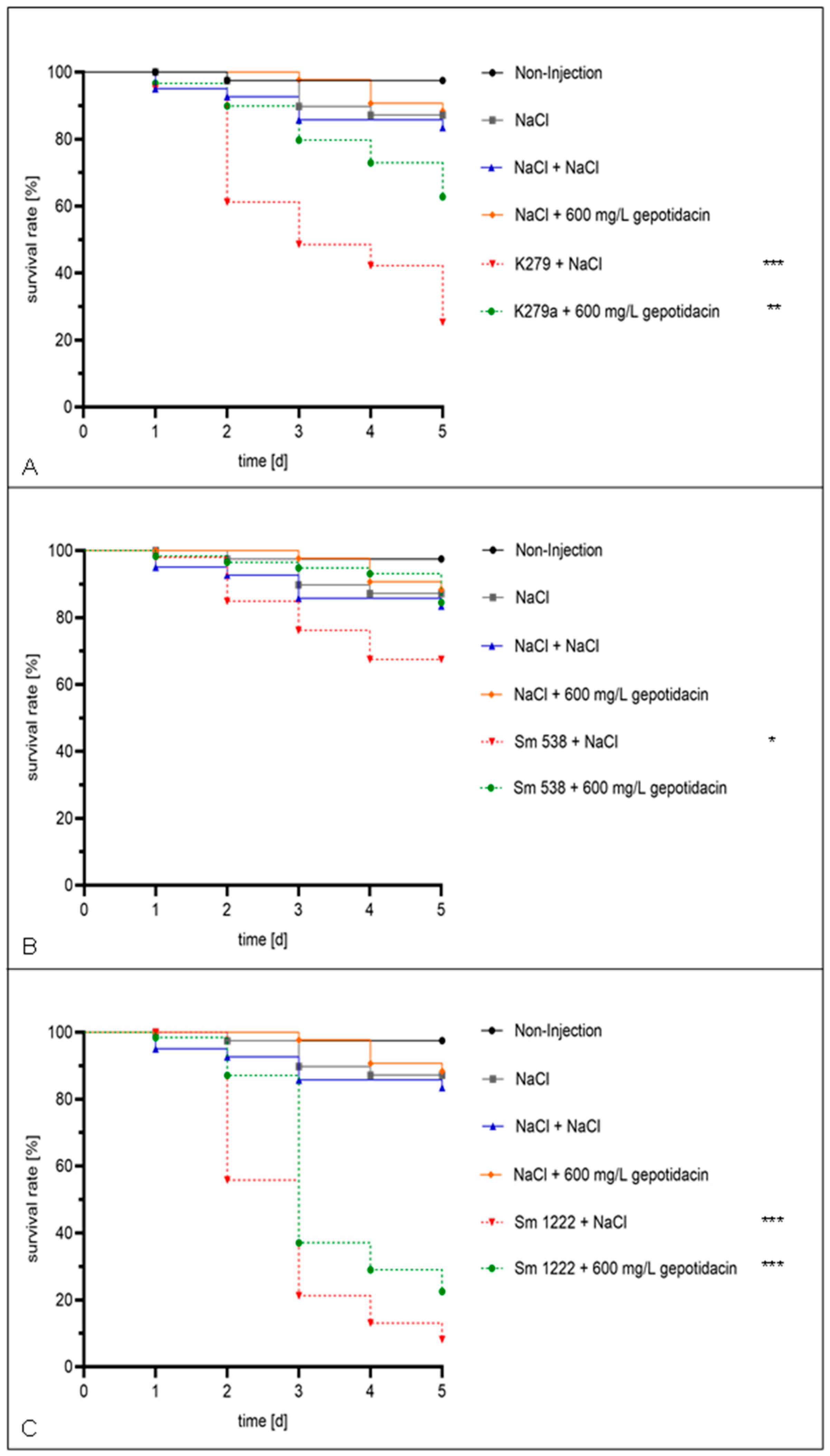
2. Results
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Susceptibility Testing
4.3. Time–Kill Assay
4.4. In Vivo Infection Assay
4.5. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denton, M.; Todd, N.J.; Kerr, K.G.; Hawkey, P.M.; Littlewood, J.M. Molecular epidemiology of Stenotrophomonas maltophilia isolated from clinical specimens from patients with cystic fibrosis and associated environmental samples. J. Clin. Microbiol. 1998, 36, 1953–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, R.P.; Fouhy, Y.; Garcia, B.F.; Watt, S.A.; Niehaus, K.; Yang, L.; Tolker-Nielsen, T.; Dow, J.M. Interspecies signaling via the Stenotrophomonas maltophilia diffusible signal factor influences biofilm formation and polymyxin tolerance in Pseudomonas aeruginosa. Mol. Microbiol. 2008, 68, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Kastoris, A.C.; Vouloumanou, E.K.; Rafailidis, P.I.; Kapaskelis, A.M.; Dimopoulos, G. Attributable mortality of Stenotrophomonas maltophilia infections: A systematic review of the literature. Future Microbiol. 2009, 4, 1103–1109. [Google Scholar] [CrossRef]
- Nicodemo, A.C.; Paez, J.I.G. Antimicrobial therapy for Stenotrophomonas maltophilia infections. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 229–237. [Google Scholar] [CrossRef]
- Nseir, S.; Di Pompeo, C.; Cavestri, B.; Jozefowicz, E.; Nyunga, M.; Soubrier, S.; Roussel-Delvallez, M.; Saulnier, F.; Mathieu, D.; Durocher, A. Multiple-drug-resistant bacteria in patients with severe acute exacerbation of chronic obstructive pulmonary disease: Prevalence, risk factors, and outcome. Crit. Care Med. 2006, 34, 2959–2966. [Google Scholar] [CrossRef]
- Weber, D.J.; Rutala, W.A.; Sickbert-Bennett, E.E.; Samsa, G.P.; Brown, V.; Niederman, M.S. Microbiology of ventilator–associated pneumonia compared with that of hospital-acquired pneumonia. Infect. Control Hosp. Epidemiol. 2007, 28, 825–831. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.-T.; Lin, C.-Y.; Lu, P.-L.; Lai, C.-C.; Chen, T.-C.; Chen, C.-Y.; Wu, D.-C.; Wang, T.-P.; Lin, C.-M.; Lin, W.-R.; et al. Stenotrophomonas maltophilia bloodstream infection: Comparison between community-onset and hospital-acquired infections. J. Microbiol. Immunol. Infect. 2014, 47, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.-H.; Chi, C.-Y.; Chen, H.-P.; Chen, T.-L.; Lai, C.-J.; Fung, C.-P.; Yu, K.-W.; Wong, W.-W.; Liu, C.-Y. Clinical characteristics and prognostic factors of patients with Stenotrophomonas maltophilia bacteremia. J. Microbiol. Immunol. Infect. 2004, 37, 350–358. [Google Scholar]
- Crossman, L.C.; Gould, V.C.; Dow, J.M.; Vernikos, G.S.; Okazaki, A.; Sebaihia, M.; Saunders, D.; Arrowsmith, C.; Carver, T.; Peters, N.; et al. The complete genome, comparative and functional analysis of Stenotrophomonas maltophilia reveals an organism heavily shielded by drug resistance determinants. Genome Biol. 2008, 9, R74. [Google Scholar] [CrossRef] [Green Version]
- Ryan, R.P.; Monchy, S.; Cardinale, M.; Taghavi, S.; Crossman, L.; Avison, M.B.; Berg, G.; van der Lelie, D.; Dow, J.M. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat. Rev. Microbiol. 2009, 7, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Gröschel, M.I.; Meehan, C.J.; Barilar, I.; Diricks, M.; Gonzaga, A.; Steglich, M.; Conchillo-Solé, O.; Scherer, I.-C.; Mamat, U.; Luz, C.F.; et al. The phylogenetic landscape and nosocomial spread of the multidrug-resistant opportunist Stenotrophomonas maltophilia. Nat. Commun. 2020, 11, 2044. [Google Scholar] [CrossRef] [PubMed]
- Vidigal, P.G.; Schmidt, D.; Stehling, F.; Mellies, U.; Steinmann, E.; Buer, J.; Rath, P.-M.; Steinmann, J. Development of a quantitative immunofluorescence assay for detection of Stenotrophomonas maltophilia antibodies in patients with cystic fibrosis. J. Cyst. Fibros. 2013, 12, 651–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, V.; Yau, Y.; Prasad, S.; Lu, A.; Atenafu, E.; Crandall, I.; Tom, S.; Tullis, E.; Ratjen, F. Stenotrophomonas maltophilia in cystic fibrosis: Serologic response and effect on lung disease. Am. J. Respir. Crit. Care Med. 2011, 183, 635–640. [Google Scholar] [CrossRef]
- Waters, V.; Atenafu, E.G.; Lu, A.; Yau, Y.; Tullis, E.; Ratjen, F. Chronic Stenotrophomonas maltophilia infection and mortality or lung transplantation in cystic fibrosis patients. J. Cyst. Fibros. 2013, 12, 482–486. [Google Scholar] [CrossRef] [Green Version]
- Vidigal, P.G.; Dittmer, S.; Steinmann, E.; Buer, J.; Rath, P.-M.; Steinmann, J. Adaptation of Stenotrophomonas maltophilia in cystic fibrosis: Molecular diversity, mutation frequency and antibiotic resistance. Int. J. Med. Microbiol. 2014, 304, 613–619. [Google Scholar] [CrossRef]
- Falagas, M.E.; Valkimadi, P.E.; Huang, Y.T.; Matthaiou, D.K.; Hsueh, P.R. Therapeutic options for Stenotrophomonas maltophilia infections beyond co-trimoxazole: A systematic review. J. Antimicrob. Chemother. 2008, 62, 889–894. [Google Scholar] [CrossRef] [Green Version]
- Bax, B.D.; Chan, P.F.; Eggleston, D.S.; Fosberry, A.; Gentry, D.R.; Gorrec, F.; Giordano, I.; Hann, M.M.; Hennessy, A.; Hibbs, M.; et al. Type IIA topoisomerase inhibition by a new class of antibacterial agents. Nature 2010, 466, 935–940. [Google Scholar] [CrossRef]
- Gibson, E.G.; Ashley, R.E.; Kerns, R.J.; Osheroff, N. Antimicrobial Resistance in the 21st Century; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Dougherty, T.J.; Nayar, A.; Newman, J.V.; Hopkins, S.; Stone, G.G.; Johnstone, M.; Shapiro, A.B.; Cronin, M.; Reck, F.; Ehmann, D.E. NBTI 5463 is a novel bacterial type ii topoisomerase inhibitor with activity against gram-negative bacteria and in vivo efficacy. Antimicrob. Agents Chemother. 2014, 58, 2657–2664. [Google Scholar] [CrossRef] [Green Version]
- Flamm, R.K.; Farrell, D.J.; Rhomberg, P.R.; Scangarella-Oman, N.E.; Sader, H.S. Gepotidacin (GSK2140944) in vitro activity against gram-positive and gram-negative bacteria. Antimicrob. Agents Chemother. 2017, 61, e00468-17. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.N.; Morris, D.H.; Avery, A.K.; Workowski, K.A.; Batteiger, B.E.; Tiffany, C.A.; Perry, C.R.; Raychaudhuri, A.; Scangarella-Oman, N.E.; Hossain, M.; et al. Gepotidacin for the treatment of uncomplicated urogenital gonorrhea: A phase 2, randomized, doseranging, single-oral dose evaluation. Clin. Infect. Dis. 2018, 67, 504–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooke, J.S.; Di Bonaventura, G.; Berg, G.; Martinez, J.-L. Editorial: A multidisciplinary look at Stenotrophomonas maltophilia: An emerging multi-drug-resistant global opportunistic pathogen. Front. Microbiol. 2017, 8, 1511. [Google Scholar] [CrossRef] [PubMed]
- Biedenbach, D.J.; Bouchillon, S.K.; Hackel, M.; Miller, L.A.; Scangarella-Oman, N.E.; Jakielaszek, C.; Sahm, D.F. In vitro activity of gepotidacin, a novel triazaacenaphthylene bacterial topoisomerase inhibitor, against a broad spectrum of bacterial pathogens. Antimicrob. Agents Chemother. 2016, 60, 1918–1923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahiri, S.D.; Kutschke, A.; McCormack, K.; Alm, R.A. Insights into the mechanism of inhibition of novel bacteria topoisomerase inhibitors from characterization of resistant mutants of Staphylococcus aureus. Antimicrob. Agents Chemother. 2015, 59, 5278–5287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoover, J.L.; Singley, C.M.; Elefante, P.; Rittenhouse, S. Efficacy of human exposures of gepotidacin (GSK2140944) against Escherichia coli in a rat pyelonephritis model. Antimicrob. Agents Chemother. 2019, 63, e00086-19. [Google Scholar] [CrossRef] [Green Version]
- Barth, A.; Hossain, M.; Brimhall, D.B.; Perry, C.R.; Tiffany, C.A.; Xu, S.; Dumont, E.F. Pharmakokinetics of oral formulations of gepotidacin (GSK2140944), a triazaacenaphthylene bacterial type II topoisomerase inhibitor, in healthy adult and adolescent participants. Antimicrob. Agents Chemother. 2022, 66, e0126321. [Google Scholar] [CrossRef]
- GlaxoSmithKline. GSK Starts a Phase III Clinical Programme for a Potential First-in-Class Antibiotic, Gepotidacin. Available online: https://www.gsk.com/en-gb/media/press-releases/gsk-starts-a-phase-iii-clinical-programme-for-a-potential-first-in-class-antibiotic-gepotidacin/ (accessed on 10 August 2021).




{kind=link}
{kind=link}
{kind=link}
| Agent | N | Method | MIC (mg/L) | ||
|---|---|---|---|---|---|
| Range | MIC50 | MIC90 | |||
| Gepotidacin | 100 | Microdilution | 0.25–16 | 2 | 8 |
| Levofloxacin | 100 | Microdilution | 0.25–>16 | 1 | 4 |
| Moxifloxacin | 100 | Microdilution | ≤0.003–>16 | 0.5 | 2 |
| Co-trimoxazole | 100 | MIC test strips | 0.016–>32 | 0.19 | >32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanders, M.I.; Ali, E.; Buer, J.; Steinmann, J.; Rath, P.-M.; Verhasselt, H.L.; Kirchhoff, L. Antibacterial Activity of the Novel Drug Gepotidacin against Stenotrophomonas maltophilia—An In Vitro and In Vivo Study. Antibiotics 2022, 11, 192. https://doi.org/10.3390/antibiotics11020192
Sanders MI, Ali E, Buer J, Steinmann J, Rath P-M, Verhasselt HL, Kirchhoff L. Antibacterial Activity of the Novel Drug Gepotidacin against Stenotrophomonas maltophilia—An In Vitro and In Vivo Study. Antibiotics. 2022; 11(2):192. https://doi.org/10.3390/antibiotics11020192
Chicago/Turabian StyleSanders, Maike Isabell, Eyhab Ali, Jan Buer, Joerg Steinmann, Peter-Michael Rath, Hedda Luise Verhasselt, and Lisa Kirchhoff. 2022. "Antibacterial Activity of the Novel Drug Gepotidacin against Stenotrophomonas maltophilia—An In Vitro and In Vivo Study" Antibiotics 11, no. 2: 192. https://doi.org/10.3390/antibiotics11020192
APA StyleSanders, M. I., Ali, E., Buer, J., Steinmann, J., Rath, P. -M., Verhasselt, H. L., & Kirchhoff, L. (2022). Antibacterial Activity of the Novel Drug Gepotidacin against Stenotrophomonas maltophilia—An In Vitro and In Vivo Study. Antibiotics, 11(2), 192. https://doi.org/10.3390/antibiotics11020192