
Persistence and Dissemination Capacities of a blaNDM-5-Harboring IncX-3 Plasmid in Escherichia coli Isolated from an Urban River in Montpellier, France


Abstract
:
1. Introduction
2. Results
2.1. Characteristics of the Two NDM-5 Producing E. coli Isolates B26 and B28
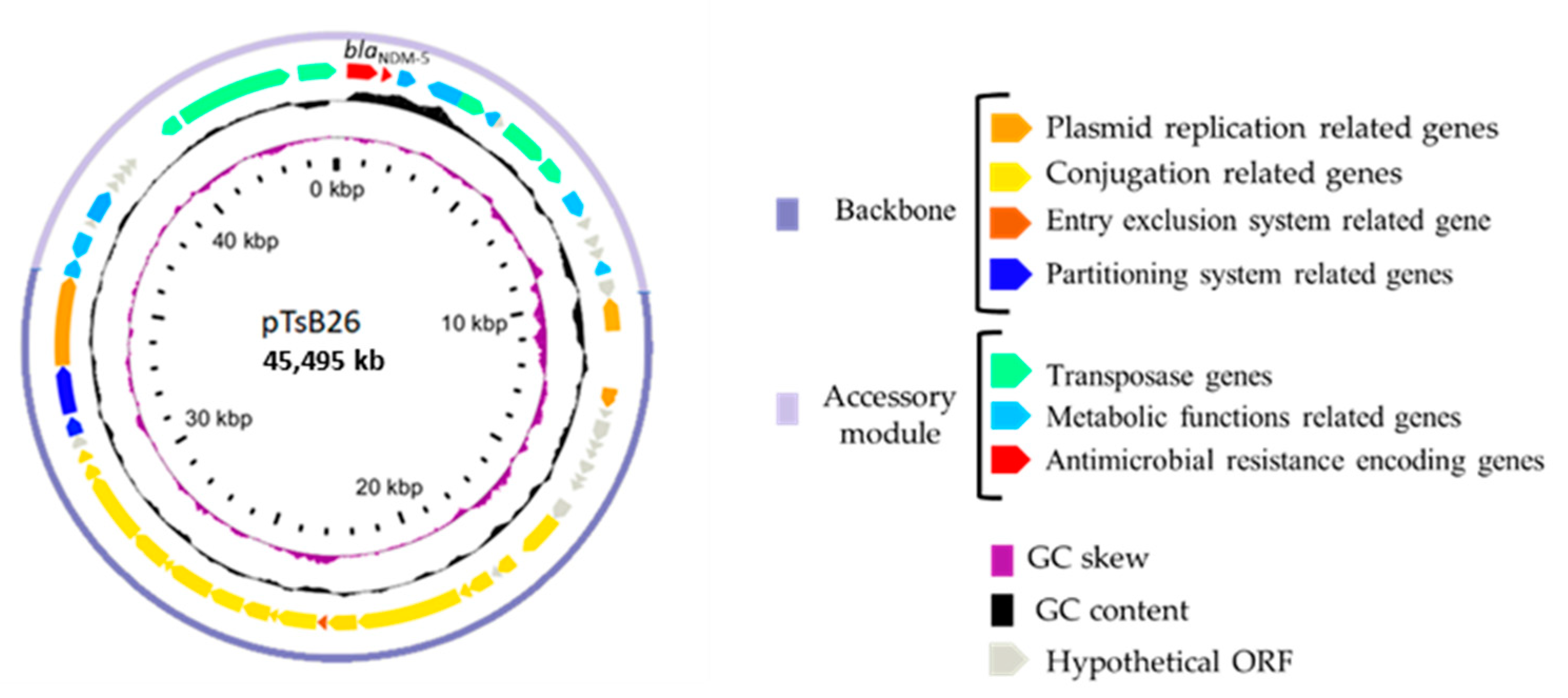
2.2. Characteristics of the blaNDM-5-Harboring IncX-3 Plasmids
2.3. Conjugative Transfer Success of pTsB26 In Vitro
2.4. In Silico Population Study of blaNDM-5-Harboring IncX-3 Plasmids among E. coli Species
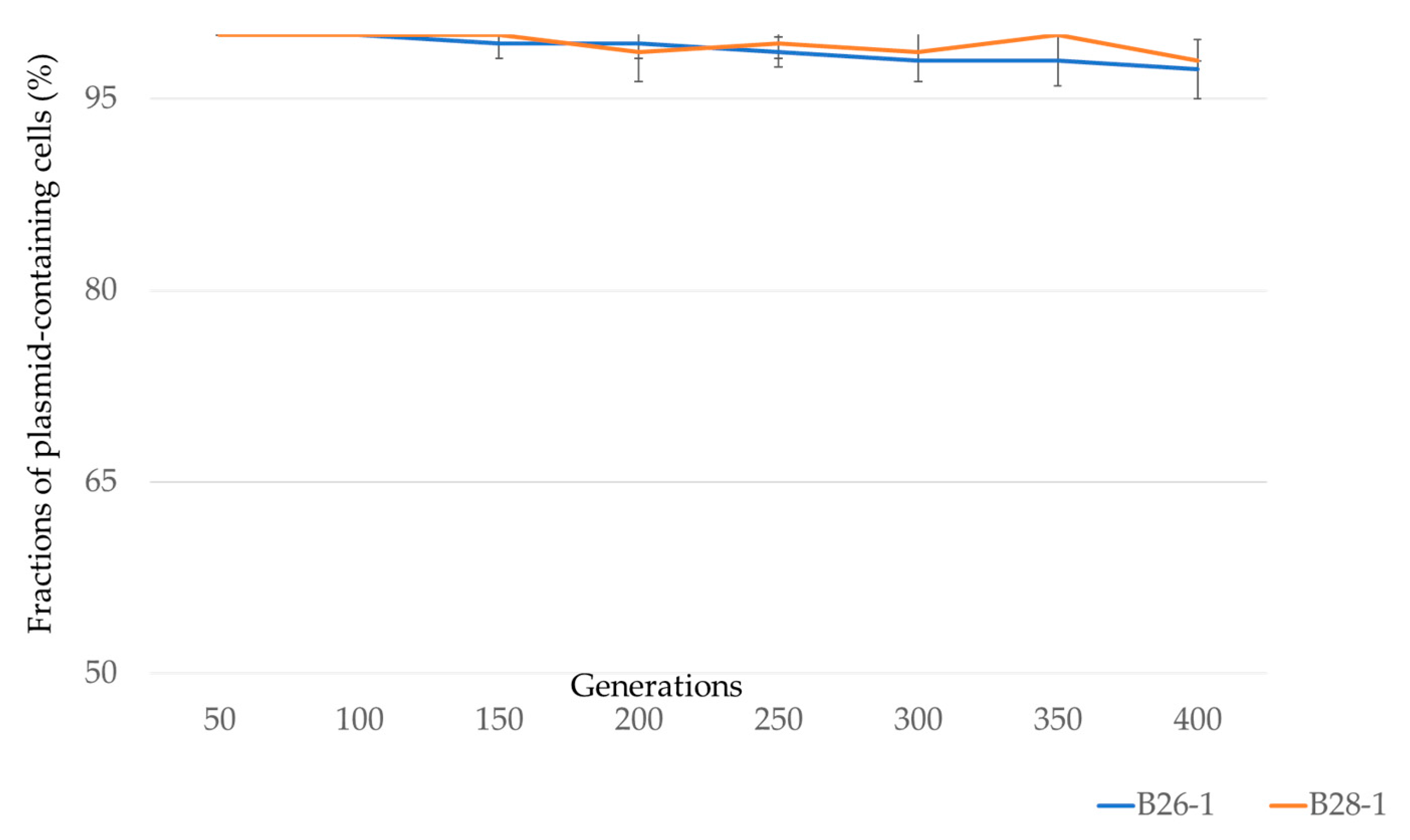
2.5. Stability and Fitness Cost of pTsB26 on B26 and B28 E. coli
3. Discussion
4. Materials and Methods
4.1. Escherichia coli Strains
4.2. Transformation and Conjugation Assays
4.3. Evaluation of Plasmid Stability
4.4. Fitness Cost of Plasmid Carriage
4.5. Carbapenem Susceptibility Testing
4.6. In Silico Analysis
4.6.1. DNA Extraction and Whole-Genome Sequencing
4.6.2. Genotyping Methods
4.6.3. Plasmid Sequence and Annotation
4.6.4. Comparative Genomics of B26 and B28 Genomes
4.6.5. Distribution of IncX3-blaNDM Plasmids in E. coli Population
4.7. Statistical Assays
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Infectious Diseases Society of America The 10 x ’20 Initiative: Pursuing a Global Commitment to Develop 10 New Antibacterial Drugs by 2020. Clin. Infect. Dis. 2010, 50, 1081–1083. [CrossRef] [PubMed] [Green Version]
- Zellweger, R.M.; Carrique-Mas, J.; Limmathurotsakul, D.; Day, N.P.J.; Thwaites, G.E.; Baker, S. Southeast Asia Antimicrobial Resistance Network A Current Perspective on Antimicrobial Resistance in Southeast Asia. J. Antimicrob. Chemother. 2017, 72, 2963–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harbarth, S.; Balkhy, H.H.; Goossens, H.; Jarlier, V.; Kluytmans, J.; Laxminarayan, R.; Saam, M.; Van Belkum, A.; Pittet, D. Antimicrobial Resistance: One World, One Fight! Antimicrob. Resist. Infect. Control 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Stewardson, A.J.; Marimuthu, K.; Sengupta, S.; Allignol, A.; El-Bouseary, M.; Carvalho, M.J.; Hassan, B.; Delgado-Ramirez, M.A.; Arora, A.; Bagga, R.; et al. Effect of Carbapenem Resistance on Outcomes of Bloodstream Infection Caused by Enterobacteriaceae in Low-Income and Middle-Income Countries (PANORAMA): A Multinational Prospective Cohort Study. Lancet Infect. Dis. 2019, 19, 601–610. [Google Scholar] [CrossRef]
- Almakki, A.; Jumas-Bilak, E.; Marchandin, H.; Licznar-Fajardo, P. Antibiotic Resistance in Urban Runoff. Sci. Total Environ. 2019, 667, 64–76. [Google Scholar] [CrossRef]
- Cherak, Z.; Loucif, L.; Moussi, A.; Rolain, J.-M. Carbapenemase-Producing Gram-Negative Bacteria in Aquatic Environments: A Review. J. Glob. Antimicrob. Resist. 2021, 25, 287–309. [Google Scholar] [CrossRef]
- Hammer-Dedet, F.; Jumas-Bilak, E.; Licznar-Fajardo, P. The Hydric Environment: A Hub for Clinically Relevant Carbapenemase Encoding Genes. Antibiotics 2020, 9, E699. [Google Scholar] [CrossRef]
- Hooban, B.; Joyce, A.; Fitzhenry, K.; Chique, C.; Morris, D. The Role of the Natural Aquatic Environment in the Dissemination of Extended Spectrum Beta-Lactamase and Carbapenemase Encoding Genes: A Scoping Review. Water Res. 2020, 180, 115880. [Google Scholar] [CrossRef]
- WHO Report on Surveillance of Antibiotic Consumption. Available online: https://www.who.int/publications-detail-redirect/who-report-on-surveillance-of-antibiotic-consumption (accessed on 18 January 2022).
- Codjoe, F.S.; Donkor, E.S. Carbapenem Resistance: A Review. Med. Sci. 2017, 6, E1. [Google Scholar] [CrossRef] [Green Version]
- Bonomo, R.A.; Burd, E.M.; Conly, J.; Limbago, B.M.; Poirel, L.; Segre, J.A.; Westblade, L.F. Carbapenemase-Producing Organisms: A Global Scourge. Clin. Infect. Dis. 2018, 66, 1290–1297. [Google Scholar] [CrossRef]
- Papp-Wallace, K.M.; Endimiani, A.; Taracila, M.A.; Bonomo, R.A. Carbapenems: Past, Present, and Future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [Green Version]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a New Metallo-Beta-Lactamase Gene, bla(NDM-1), and a Novel Erythromycin Esterase Gene Carried on a Unique Genetic Structure in Klebsiella pneumoniae Sequence Type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Dortet, L.; Bernabeu, S.; Nordmann, P. Genetic Features of blaNDM-1-Positive Enterobacteriaceae. Antimicrob. Agents Chemother. 2011, 55, 5403–5407. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Poirel, L.; Walsh, T.R.; Livermore, D.M. The Emerging NDM Carbapenemases. Trends Microbiol. 2011, 19, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Naas, T.; Poirel, L. Global Spread of Carbapenemase-Producing Enterobacteriaceae. Emerg. Infect. Dis. 2011, 17, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of NDM-1 Positive Bacteria in the New Delhi Environment and Its Implications for Human Health: An Environmental Point Prevalence Study. Lancet Infect. Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Basu, S. Variants of the New Delhi Metallo-β-Lactamase: New Kids on the Block. Future Microbiol. 2020, 15, 465–467. [Google Scholar] [CrossRef]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef] [Green Version]
- Bontron, S.; Nordmann, P.; Poirel, L. Transposition of Tn125 Encoding the NDM-1 Carbapenemase in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 7245–7251. [Google Scholar] [CrossRef] [Green Version]
- Rolain, J.M.; Parola, P.; Cornaglia, G. New Delhi Metallo-Beta-Lactamase (NDM-1): Towards a New Pandemia? Clin. Microbiol. Infect. 2010, 16, 1699–1701. [Google Scholar] [CrossRef] [Green Version]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a New Antibiotic Resistance Mechanism in India, Pakistan, and the UK: A Molecular, Biological, and Epidemiological Study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Khan, N.H.; Ishii, Y.; Kimata-Kino, N.; Esaki, H.; Nishino, T.; Nishimura, M.; Kogure, K. Isolation of Pseudomonas aeruginosa from Open Ocean and Comparison with Freshwater, Clinical, and Animal Isolates. Microb. Ecol. 2007, 53, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Mitra, S.; Chattopadhyay, P.; Som, T.; Mukherjee, S.; Basu, S. Spread and Exchange of bla NDM-1 in Hospitalized Neonates: Role of Mobilizable Genetic Elements. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 255–265. [Google Scholar] [CrossRef]
- Johnson, T.J.; Bielak, E.M.; Fortini, D.; Hansen, L.H.; Hasman, H.; Debroy, C.; Nolan, L.K.; Carattoli, A. Expansion of the IncX Plasmid Family for Improved Identification and Typing of Novel Plasmids in Drug-Resistant Enterobacteriaceae. Plasmid 2012, 68, 43–50. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Guo, L.; Zhou, L.; Ma, Y.; Luo, Y.; Tao, C.; Yang, J. NDM-Producing Enterobacteriaceae in a Chinese Hospital, 2014–2015: Identification of NDM-Producing Citrobacter werkmanii and Acquisition of blaNDM-1-Carrying Plasmid in Vivo in a Clinical Escherichia coli Isolate. J. Med. Microbiol. 2016, 65, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xie, L.; Wang, X.; Han, L.; Guo, X.; Ni, Y.; Qu, H.; Sun, J. Further Spread of bla NDM-5 in Enterobacteriaceae via IncX3 Plasmids in Shanghai, China. Front. Microbiol. 2016, 7, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, T.; Fu, J.; Xie, N.; Ma, S.; Lei, L.; Zhai, W.; Shen, Y.; Sun, C.; Wang, S.; Shen, Z.; et al. Fitness Cost of blaNDM-5-Carrying P3R-IncX3 Plasmids in Wild-Type NDM-Free Enterobacteriaceae. Microorganisms 2020, 8, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almakki, A.; Maure, A.; Pantel, A.; Romano-Bertrand, S.; Masnou, A.; Marchandin, H.; Jumas-Bilak, E.; Licznar-Fajardo, P. NDM-5-Producing Escherichia coli in an Urban River in Montpellier, France. Int. J. Antimicrob. Agents 2017, 50, 123–124. [Google Scholar] [CrossRef]
- Liakopoulos, A.; van der Goot, J.; Bossers, A.; Betts, J.; Brouwer, M.S.M.; Kant, A.; Smith, H.; Ceccarelli, D.; Mevius, D. Genomic and Functional Characterisation of IncX3 Plasmids Encoding blaSHV-12 in Escherichia coli from Human and Animal Origin. Sci. Rep. 2018, 8, 7674. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Fu, Y.; Shen, M.; Huang, D.; Du, X.; Hu, Q.; Zhou, Y.; Wang, D.; Yu, Y. Dissemination of blaNDM-5 Gene via an IncX3-Type Plasmid among Non-Clonal Escherichia coli in China. Antimicrob. Resist. Infect. Control 2018, 7, 59. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, M.; Shen, X.; Wang, L.; Liu, L.; Hao, Z.; Duan, J.; Yu, F. Outbreak of blaNDM-5-Harboring Klebsiella pneumoniae ST290 in a Tertiary Hospital in China. Microb. Drug Resist. 2019, 25, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Parvez, S.; Khan, A.U. Hospital Sewage Water: A Reservoir for Variants of New Delhi Metallo-β-Lactamase (NDM)- and Extended-Spectrum β-Lactamase (ESBL)-Producing Enterobacteriaceae. Int. J. Antimicrob. Agents 2018, 51, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Zurfluh, K.; Stevens, M.J.A.; Stephan, R.; Nüesch-Inderbinen, M. Complete and Assembled Genome Sequence of an NDM-5- and CTX-M-15-Producing Escherichia coli Sequence Type 617 Isolated from Wastewater in Switzerland. J. Glob. Antimicrob. Resist. 2018, 15, 105–106. [Google Scholar] [CrossRef] [Green Version]
- Caractéristiques et Évolution Des Souches D’entérobactéries Productrices de Carbapénémases (EPC) Isolées En France, 2012–2020. Available online: https://www.santepubliquefrance.fr/maladies-et-traumatismes/infections-associees-aux-soins-et-resistance-aux-antibiotiques/resistance-aux-antibiotiques/documents/article/caracteristiques-et-evolution-des-souches-d-enterobacteries-productrices-de-carbapenemases-epc-isolees-en-france-2012-2020 (accessed on 21 January 2022).
- Li, Y.; Tang, M.; Dai, X.; Zhou, Y.; Zhang, Z.; Qiu, Y.; Li, C.; Zhang, L. Whole-Genomic Analysis of NDM-5-Producing Enterobacteriaceae Recovered from an Urban River in China. Infect. Drug Resist. 2021, 14, 4427–4440. [Google Scholar] [CrossRef]
- Bleichenbacher, S.; Stevens, M.J.A.; Zurfluh, K.; Perreten, V.; Endimiani, A.; Stephan, R.; Nüesch-Inderbinen, M. Environmental Dissemination of Carbapenemase-Producing Enterobacteriaceae in Rivers in Switzerland. Environ. Pollut. 2020, 265, 115081. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Berglund, B.; Zou, H.; Zhou, Z.; Xia, H.; Zhao, L.; Nilsson, L.E.; Li, X. Dissemination of blaNDM-5 via IncX3 Plasmids in Carbapenem-Resistant Enterobacteriaceae among Humans and in the Environment in an Intensive Vegetable Cultivation Area in Eastern China. Environ. Pollut. 2021, 273, 116370. [Google Scholar] [CrossRef]
- Zhai, R.; Fu, B.; Shi, X.; Sun, C.; Liu, Z.; Wang, S.; Shen, Z.; Walsh, T.R.; Cai, C.; Wang, Y.; et al. Contaminated In-House Environment Contributes to the Persistence and Transmission of NDM-Producing Bacteria in a Chinese Poultry Farm. Environ. Int. 2020, 139, 105715. [Google Scholar] [CrossRef]
- Zou, H.; Jia, X.; Liu, H.; Li, S.; Wu, X.; Huang, S. Emergence of NDM-5-Producing Escherichia coli in a Teaching Hospital in Chongqing, China: IncF-Type Plasmids May Contribute to the Prevalence of Bla NDM-5. Front. Microbiol. 2020, 11, 334. [Google Scholar] [CrossRef]
- Wang, Y.; Tong, M.-K.; Chow, K.-H.; Cheng, V.C.-C.; Tse, C.W.-S.; Wu, A.K.-L.; Lai, R.W.-M.; Luk, W.-K.; Tsang, D.N.-C.; Ho, P.-L. Occurrence of Highly Conjugative IncX3 Epidemic Plasmid Carrying bla NDM in Enterobacteriaceae Isolates in Geographically Widespread Areas. Front. Microbiol. 2018, 9, 2272. [Google Scholar] [CrossRef]
- Sonnevend, A.; Al Baloushi, A.; Ghazawi, A.; Hashmey, R.; Girgis, S.; Hamadeh, M.B.; Al Haj, M.; Pál, T. Emergence and Spread of NDM-1 Producer Enterobacteriaceae with Contribution of IncX3 Plasmids in the United Arab Emirates. J. Med. Microbiol. 2013, 62, 1044–1050. [Google Scholar] [CrossRef] [Green Version]
- Ho, P.-L.; Li, Z.; Lai, E.L.; Chiu, S.S.; Cheng, V.C.C. Emergence of NDM-1-Producing Enterobacteriaceae in China. J. Antimicrob. Chemother. 2012, 67, 1553–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortini, D.; Villa, L.; Feudi, C.; Pires, J.; Bonura, C.; Mammina, C.; Endimiani, A.; Carattoli, A. Double Copies of Bla(KPC-3)::Tn4401a on an IncX3 Plasmid in Klebsiella pneumoniae Successful Clone ST512 from Italy. Antimicrob. Agents Chemother. 2016, 60, 646–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espedido, B.A.; Dimitrijovski, B.; van Hal, S.J.; Jensen, S.O. The Use of Whole-Genome Sequencing for Molecular Epidemiology and Antimicrobial Surveillance: Identifying the Role of IncX3 Plasmids and the Spread of BlaNDM-4-like Genes in the Enterobacteriaceae. J. Clin. Pathol. 2015, 68, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Wailan, A.M.; Paterson, D.L.; Kennedy, K.; Ingram, P.R.; Bursle, E.; Sidjabat, H.E. Genomic Characteristics of NDM-Producing Enterobacteriaceae Isolates in Australia and Their blaNDM Genetic Contexts. Antimicrob. Agents Chemother. 2016, 60, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Wang, Y.; Walsh, T.R.; Liu, D.; Shen, Z.; Zhang, R.; Yin, W.; Yao, H.; Li, J.; Shen, J. Plasmid-Mediated Novel blaNDM-17 Gene Encoding a Carbapenemase with Enhanced Activity in a Sequence Type 48 Escherichia coli Strain. Antimicrob. Agents Chemother. 2017, 61, e02233-16. [Google Scholar] [CrossRef] [Green Version]
- Kieffer, N.; Nordmann, P.; Aires-de-Sousa, M.; Poirel, L. High Prevalence of Carbapenemase-Producing Enterobacteriaceae among Hospitalized Children in Luanda, Angola. Antimicrob. Agents Chemother. 2016, 60, 6189–6192. [Google Scholar] [CrossRef] [Green Version]
- Petrosillo, N.; Vranić-Ladavac, M.; Feudi, C.; Villa, L.; Fortini, D.; Barišić, N.; Bedenić, B.; Ladavac, R.; D’Arezzo, S.; Andrašević, A.T.; et al. Spread of Enterobacter cloacae Carrying blaNDM-1, blaCTX-M-15, blaSHV-12 and Plasmid-Mediated Quinolone Resistance Genes in a Surgical Intensive Care Unit in Croatia. J. Glob. Antimicrob. Resist. 2016, 4, 44–48. [Google Scholar] [CrossRef]
- Ho, P.-L.; Wang, Y.; Liu, M.C.-J.; Lai, E.L.-Y.; Law, P.Y.-T.; Cao, H.; Chow, K.-H. IncX3 Epidemic Plasmid Carrying blaNDM-5 in Escherichia coli from Swine in Multiple Geographic Areas in China. Antimicrob. Agents Chemother. 2018, 62, e02295-17. [Google Scholar] [CrossRef] [Green Version]
- Volante, A.; Alonso, J.C. Molecular Anatomy of ParA-ParA and ParA-ParB Interactions during Plasmid Partitioning. J. Biol. Chem. 2015, 290, 18782–18795. [Google Scholar] [CrossRef] [Green Version]
- De Gelder, L.; Ponciano, J.M.; Joyce, P.; Top, E.M. Stability of a Promiscuous Plasmid in Different Hosts: No Guarantee for a Long-Term Relationship. Microbiology 2007, 153, 452–463. [Google Scholar] [CrossRef] [Green Version]
- Andersson, D.I.; Patin, S.M.; Nilsson, A.I.; Kugelberg, E. The Biological Cost of Antibiotic Resistance. In Enzyme-Mediated Resistance to Antibiotics; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007; pp. 339–348. ISBN 978-1-68367-166-4. [Google Scholar]
- Göttig, S.; Pfeifer, Y.; Wichelhaus, T.A.; Zacharowski, K.; Bingold, T.; Averhoff, B.; Brandt, C.; Kempf, V.A. Global Spread of New Delhi Metallo-β-Lactamase 1. Lancet Infect. Dis. 2010, 10, 828–829. [Google Scholar] [CrossRef]
- Baltrus, D.A. Exploring the Costs of Horizontal Gene Transfer. Trends Ecol. Evol. 2013, 28, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Long, D.; Zhu, L.-L.; Du, F.-L.; Xiang, T.-X.; Wan, L.-G.; Wei, D.-D.; Zhang, W.; Liu, Y. Phenotypical Profile and Global Transcriptomic Profile of Hypervirulent Klebsiella pneumoniae Due to Carbapenemase-Encoding Plasmid Acquisition. BMC Genom. 2019, 20, 480. [Google Scholar] [CrossRef]
- Doyle, M.; Fookes, M.; Ivens, A.; Mangan, M.W.; Wain, J.; Dorman, C.J. An H-NS-like Stealth Protein Aids Horizontal DNA Transmission in Bacteria. Science 2007, 315, 251–252. [Google Scholar] [CrossRef] [PubMed]
- Lucchini, S.; Rowley, G.; Goldberg, M.D.; Hurd, D.; Harrison, M.; Hinton, J.C.D. H-NS Mediates the Silencing of Laterally Acquired Genes in Bacteria. PLoS Pathog. 2006, 2, e81. [Google Scholar] [CrossRef]
- Navarre, W.W.; Porwollik, S.; Wang, Y.; McClelland, M.; Rosen, H.; Libby, S.J.; Fang, F.C. Selective Silencing of Foreign DNA with Low GC Content by the H-NS Protein in Salmonella. Science 2006, 313, 236–238. [Google Scholar] [CrossRef] [PubMed]
- San Millan, A.; MacLean, R.C. Fitness Costs of Plasmids: A Limit to Plasmid Transmission. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Kumwenda, G.P.; Sugawara, Y.; Abe, R.; Akeda, Y.; Kasambara, W.; Chizani, K.; Takeuchi, D.; Sakamoto, N.; Tomono, K.; Hamada, S. First Identification and Genomic Characterization of Multidrug-Resistant Carbapenemase-Producing Enterobacteriaceae Clinical Isolates in Malawi, Africa. J. Med. Microbiol. 2019, 68, 1707–1715. [Google Scholar] [CrossRef] [Green Version]
- van Hoek, A.H.A.M.; Veenman, C.; Florijn, A.; Huijbers, P.M.C.; Graat, E.A.M.; de Greeff, S.; Dierikx, C.M.; van Duijkeren, E. Longitudinal Study of ESBL Escherichia coli Carriage on an Organic Broiler Farm. J. Antimicrob. Chemother. 2018, 73, 3298–3304. [Google Scholar] [CrossRef]
- Baniga, Z.; Hounmanou, Y.M.G.; Kudirkiene, E.; Kusiluka, L.J.M.; Mdegela, R.H.; Dalsgaard, A. Genome-Based Analysis of Extended-Spectrum β-Lactamase-Producing Escherichia coli in the Aquatic Environment and Nile Perch (Lates Niloticus) of Lake Victoria, Tanzania. Front. Microbiol. 2020, 11, 108. [Google Scholar] [CrossRef] [Green Version]
- Büdel, T.; Kuenzli, E.; Clément, M.; Bernasconi, O.J.; Fehr, J.; Mohammed, A.H.; Hassan, N.K.; Zinsstag, J.; Hatz, C.; Endimiani, A. Polyclonal Gut Colonization with Extended-Spectrum Cephalosporin- and/or Colistin-Resistant Enterobacteriaceae: A Normal Status for Hotel Employees on the Island of Zanzibar, Tanzania. J. Antimicrob. Chemother. 2019, 74, 2880–2890. [Google Scholar] [CrossRef] [PubMed]
- Grimont, F.; Grimont, P.A.D. The Genus Enterobacter. In The Prokaryotes: A Handbook on the Biology of Bacteria: Proteobacteria: Gamma Subclass; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 6, pp. 197–214. ISBN 978-0-387-30746-6. [Google Scholar]
- Drancourt, M.; Bollet, C.; Carta, A.; Rousselier, P. Phylogenetic Analyses of Klebsiella Species Delineate Klebsiella and Raoultella Gen. Nov., with Description of Raoultella ornithinolytica Comb. Nov., Raoultella terrigena Comb. Nov. and Raoultella planticola Comb. Nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Mills, M.C.; Lee, J. The Threat of Carbapenem-Resistant Bacteria in the Environment: Evidence of Widespread Contamination of Reservoirs at a Global Scale. Environ. Pollut. 2019, 255, 113143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, W.; Chen, J.; Qi, W.; Wang, F.; Zhou, Y. Impact of Biofilm Formation and Detachment on the Transmission of Bacterial Antibiotic Resistance in Drinking Water Distribution Systems. Chemosphere 2018, 203, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Sanganyado, E.; Gwenzi, W. Antibiotic Resistance in Drinking Water Systems: Occurrence, Removal, and Human Health Risks. Sci. Total Environ. 2019, 669, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Hölzel, C.S.; Tetens, J.L.; Schwaiger, K. Unraveling the Role of Vegetables in Spreading Antimicrobial-Resistant Bacteria: A Need for Quantitative Risk Assessment. Foodborne Pathog. Dis. 2018, 15, 671–688. [Google Scholar] [CrossRef] [PubMed]
- Laurens, C.; Jean-Pierre, H.; Licznar-Fajardo, P.; Hantova, S.; Godreuil, S.; Martinez, O.; Jumas-Bilak, E. Transmission of IMI-2 Carbapenemase-Producing Enterobacteriaceae from River Water to Human. J. Glob. Antimicrob. Resist. 2018, 15, 88–92. [Google Scholar] [CrossRef]
- Bosch, T.; Schade, R.; Landman, F.; Schouls, L.; Dijk, K. van A blaVIM-1 Positive Aeromonas hydrophila Strain in a near-Drowning Patient: Evidence for Interspecies Plasmid Transfer within the Patient. Future Microbiol. 2019, 14, 1191–1197. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of Plasmids by PCR-Based Replicon Typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Sandegren, L.; Linkevicius, M.; Lytsy, B.; Melhus, Å.; Andersson, D.I. Transfer of an Escherichia coli ST131 Multiresistance Cassette Has Created a Klebsiella pneumoniae-Specific Plasmid Associated with a Major Nosocomial Outbreak. J. Antimicrob. Chemother. 2012, 67, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Bonnel, C.; Legrand, B.; Simon, M.; Clavié, M.; Masnou, A.; Jumas-Bilak, E.; Kang, Y.K.; Licznar-Fajardo, P.; Maillard, L.T.; Masurier, N. Tailoring the Physicochemical Properties of Antimicrobial Peptides onto a Thiazole-Based γ-Peptide Foldamer. J. Med. Chem. 2020, 63, 9168–9180. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Gordon, D.; Denamur, E. Guide to the Various Phylogenetic Classification Schemes for Escherichia coli and the Correspondence among Schemes. Microbiology 2015, 161, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Waters, N.R.; Abram, F.; Brennan, F.; Holmes, A.; Pritchard, L. Easy Phylotyping of Escherichia coli via the EzClermont Web App and Command-Line Tool. Access Microbiol. 2020, 2. [Google Scholar] [CrossRef]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple Alignment of Conserved Genomic Sequence with Rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids Using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; van den Beek, M.; Blankenberg, D.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Eberhard, C.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2016 Update. Nucleic Acids Res. 2016, 44, W3–W10. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.M.; Thomson, N.R.; Cerdeño-Tárraga, A.M.; Brown, C.J.; Top, E.M.; Frost, L.S. Annotation of Plasmid Genes. Plasmid 2017, 91, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Madden, T.L.; Tatusov, R.L.; Zhang, J. Applications of Network BLAST Server. Methods Enzymol. 1996, 266, 131–141. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro Protein Families and Domains Database: 20 Years On. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Conant, G.C.; Wolfe, K.H. GenomeVx: Simple Web-Based Creation of Editable Circular Chromosome Maps. Bioinformatics 2008, 24, 861–862. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Alikhan, N.-F.; Mohamed, K.; Fan, Y.; Agama Study Group; Achtman, M. The EnteroBase User’s Guide, with Case Studies on Salmonella Transmissions, Yersinia pestis Phylogeny, and Escherichia Core Genomic Diversity. Genome Res. 2020, 30, 138–152. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, M.; Sousa, A.; Ramirez, M.; Francisco, A.P.; Carriço, J.A.; Vaz, C. PHYLOViZ 2.0: Providing Scalable Data Integration and Visualization for Multiple Phylogenetic Inference Methods. Bioinformatics 2017, 33, 128–129. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor Strains | Conjugative Frequency |
|---|---|
| B26 | 2.09 × 10−3 |
| B28 | 4.81 × 10−4 |
| TsB26 | 3.52 × 10−3 |
| TsB28 | 3.76 × 10−3 |
| Strain | Accession Number | ST | Phylotype | Country | Source Type | Source Niche | Year |
|---|---|---|---|---|---|---|---|
| CRE1493 | CP019071 | 167 | A | China | rectal swab | homo sapiens | 2013 |
| 165 | CP020509 | 101 | B1 | USA | abdominal | homo sapiens | 2015 |
| CREC-591 | CP024821 | 101 | B1 | South Korea | peritoneal fluid | homo sapiens | 2015 |
| WCHEC025943 | CP027205 | 410 | C | China | wastewater | environment | 2017 |
| WCHEC005784 | CP028578 | 617 | A | China | rectal swab | homo sapiens | 2014 |
| 135 | CP028632 | 11 | E | Canada | NA | livestock | 2006 |
| ECCRA-119 | CP029242 | 156 | B1 | China | stools | dog | 2017 |
| CH613 | MCRE01000001 | 10 | A | China | urine | homo sapiens | 2015 |
| GSH8M-2 | NZ_AP019675 | 542 | A | Japan | wastewater treatment plant | environment | 2018 |
| WP8-S18-CRE-02 | NZ_AP022245 | 542 | A | Japan | wastewater treatment plant | environment | 2018 |
| TUM18781 | NZ_AP023205 | 2040 | B1 | Japan | NA | homo sapiens | 2018 |
| YJ3 | NZ_AP023226 | 10 | A | Myanma | stools | homo sapiens | 2018 |
| WCHEC005237 | NZ_CP026580 | 167 | A | China | rectal swab | homo sapiens | 2014 |
| SCEC020001 | NZ_CP032426 | 410 | C | China | urinary tract | homo sapiens | 2016 |
| SCEC020022 | NZ_CP032892 | 156 | B1 | China | stools | homo sapiens | 2016 |
| WCHEC020031 | NZ_CP033401 | 410 | C | China | NA | homo sapiens | 2016 |
| L37 | NZ_CP034589 | 48 | A | China | rectal swab | homo sapiens | 2018 |
| L65 | NZ_CP034738 | 3076 | B1 | China | NA | homo sapiens | 2018 |
| SCEC020026 | NZ_CP034958 | 410 | C | China | NA | homo sapiens | 2016 |
| WCHEC020032 | NZ_CP034966 | 410 | C | China | NA | homo sapiens | 2016 |
| WCHEC025970 | NZ_CP036177 | 167 | A | China | NA | homo sapiens | 2017 |
| L725 | NZ_CP036202 | 2161 | B1 | China | stools | homo sapiens | 2018 |
| EC-129 | NZ_CP038453 | 167 | A | Japan | sputum | homo sapiens | 2018 |
| GZ04-0086 | NZ_CP042336 | 44 | A | China | stools | homo sapiens | 2018 |
| GZEC065 | NZ_CP048025 | 156 | B1 | China | blood | homo sapiens | 2017 |
| pV11-19-E11-025-038 | NZ_CP049050 | 1721 | A | South Korea | NA | dog | 2019 |
| 3R | NZ_CP049348 | 156 | B1 | China | NA | poultry | 2015 |
| SFE8 | NZ_CP051219 | 533 | B1 | China | stools | pork | 2019 |
| B26 | B26 | 636 | B2 | France | urban water | environment | 2015 |
| B28 | B28 | 636 | B2 | France | urban water | environment | 2015 |
| Strain | µmax (h−1) (±sd) |
|---|---|
| B26 | 0.43351667 (±0.02560949) a |
| B26ΔpJ40 | 0.4441375 (±0.02775884) a |
| B26J40a | 0.44380714 (±0.03785335) a |
| B28 | 0.43683889 (±0.0217273) a, b |
| B28ΔpJ19 | 0.40048824 (±0.0333677) c |
| B28J19a | 0.43095 (±0.03460016) a, b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hammer-Dedet, F.; Aujoulat, F.; Jumas-Bilak, E.; Licznar-Fajardo, P. Persistence and Dissemination Capacities of a blaNDM-5-Harboring IncX-3 Plasmid in Escherichia coli Isolated from an Urban River in Montpellier, France. Antibiotics 2022, 11, 196. https://doi.org/10.3390/antibiotics11020196
Hammer-Dedet F, Aujoulat F, Jumas-Bilak E, Licznar-Fajardo P. Persistence and Dissemination Capacities of a blaNDM-5-Harboring IncX-3 Plasmid in Escherichia coli Isolated from an Urban River in Montpellier, France. Antibiotics. 2022; 11(2):196. https://doi.org/10.3390/antibiotics11020196
Chicago/Turabian StyleHammer-Dedet, Florence, Fabien Aujoulat, Estelle Jumas-Bilak, and Patricia Licznar-Fajardo. 2022. "Persistence and Dissemination Capacities of a blaNDM-5-Harboring IncX-3 Plasmid in Escherichia coli Isolated from an Urban River in Montpellier, France" Antibiotics 11, no. 2: 196. https://doi.org/10.3390/antibiotics11020196
APA StyleHammer-Dedet, F., Aujoulat, F., Jumas-Bilak, E., & Licznar-Fajardo, P. (2022). Persistence and Dissemination Capacities of a blaNDM-5-Harboring IncX-3 Plasmid in Escherichia coli Isolated from an Urban River in Montpellier, France. Antibiotics, 11(2), 196. https://doi.org/10.3390/antibiotics11020196