
Campylobacter Bacteriophage Cocktail Design Based on an Advanced Selection Scheme

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Bacteriophage Isolation and Propagation
2.3. Phage Characterization
2.3.1. Host Range/EOP
2.3.2. Phage Classification
Negatively Stained Virions for Electron Micrographs
DNA-Based Analysis
2.3.3. Phage Tests in Liquid Culture
Planktonic Killing Assay (PKA)
Comparison of Phage Concentrations after 24 h
2.4. Data Analysis
3. Results
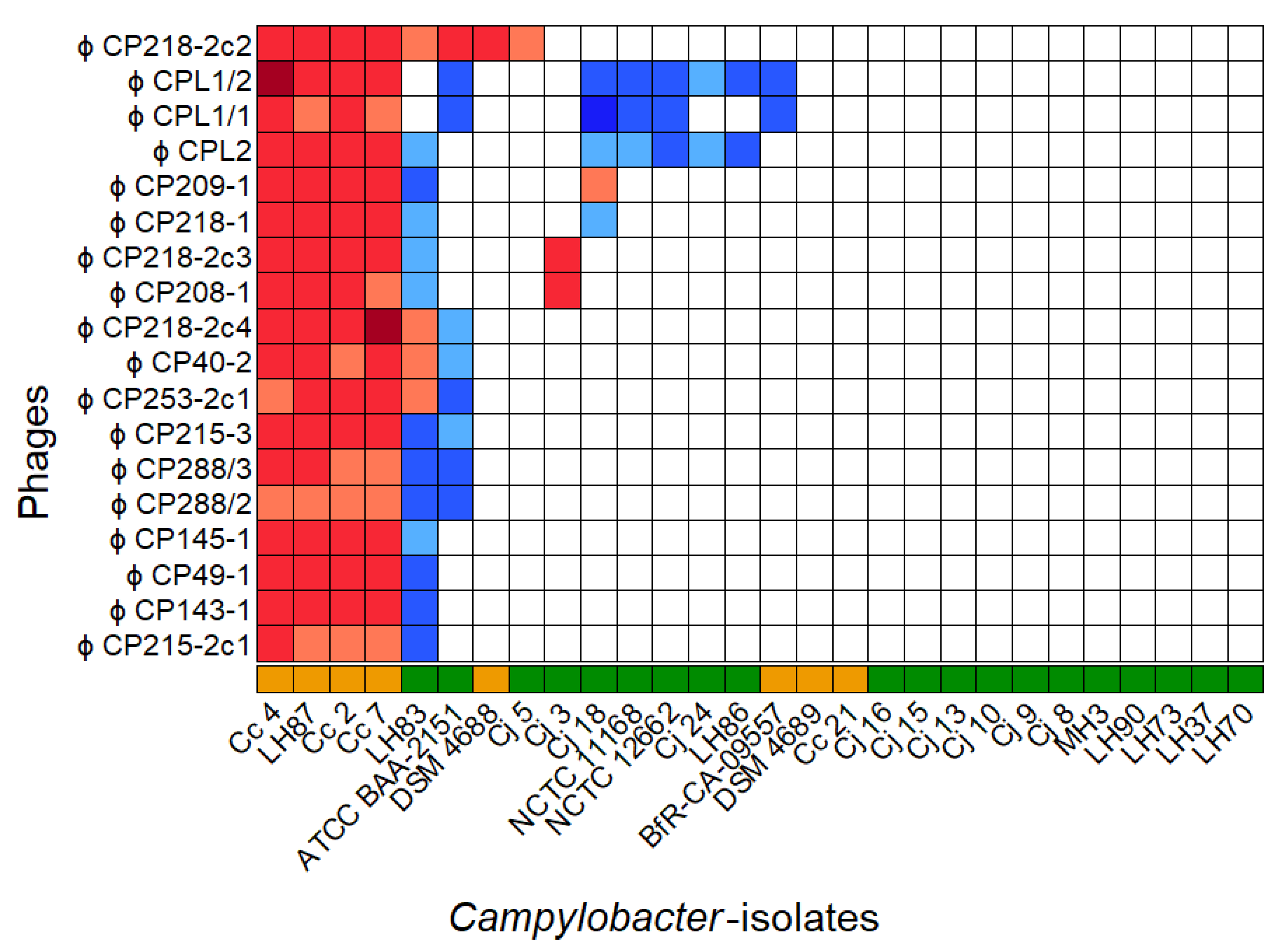
3.1. Phage Isolation and Host Range Determination
3.2. Phage Classification
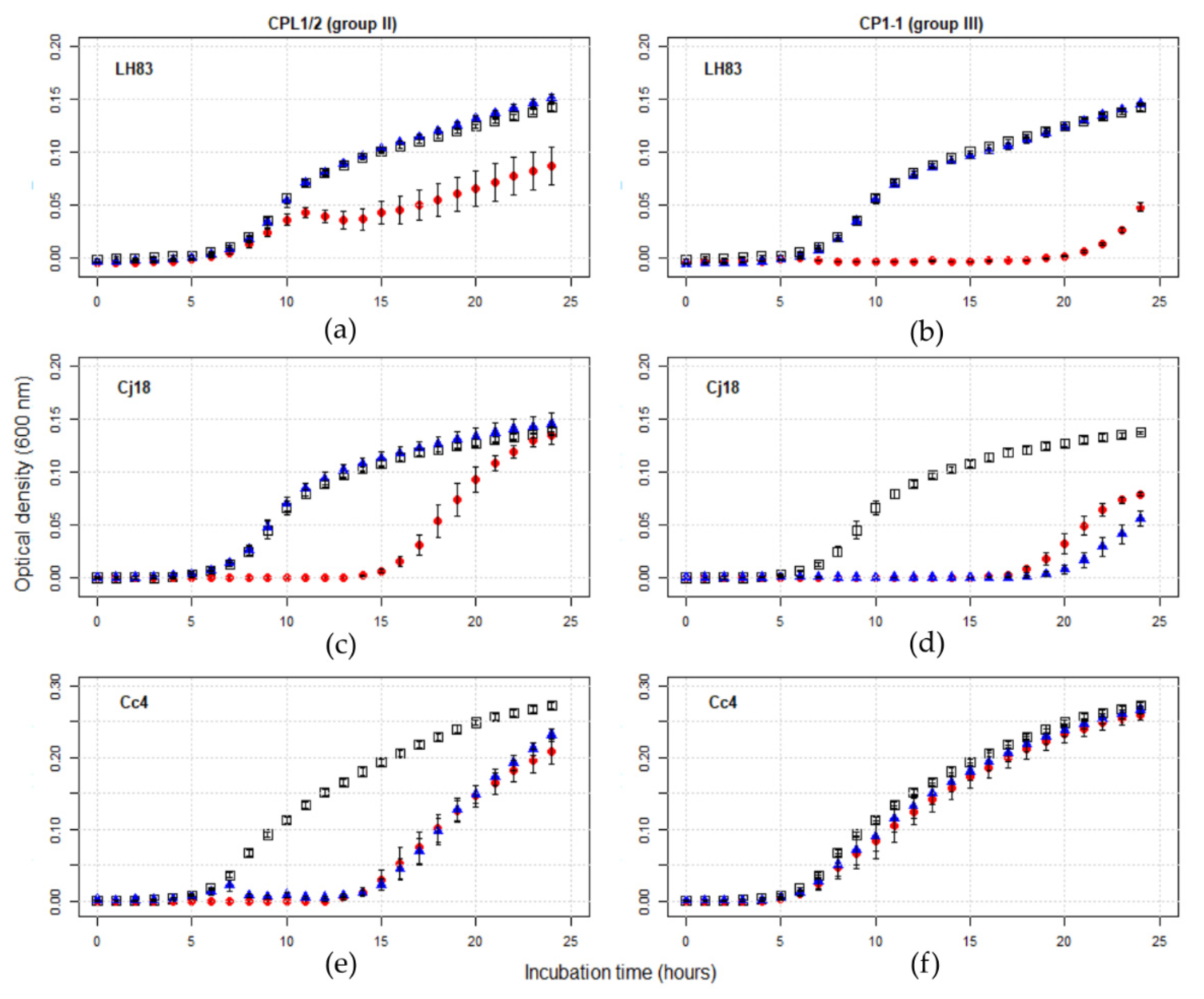
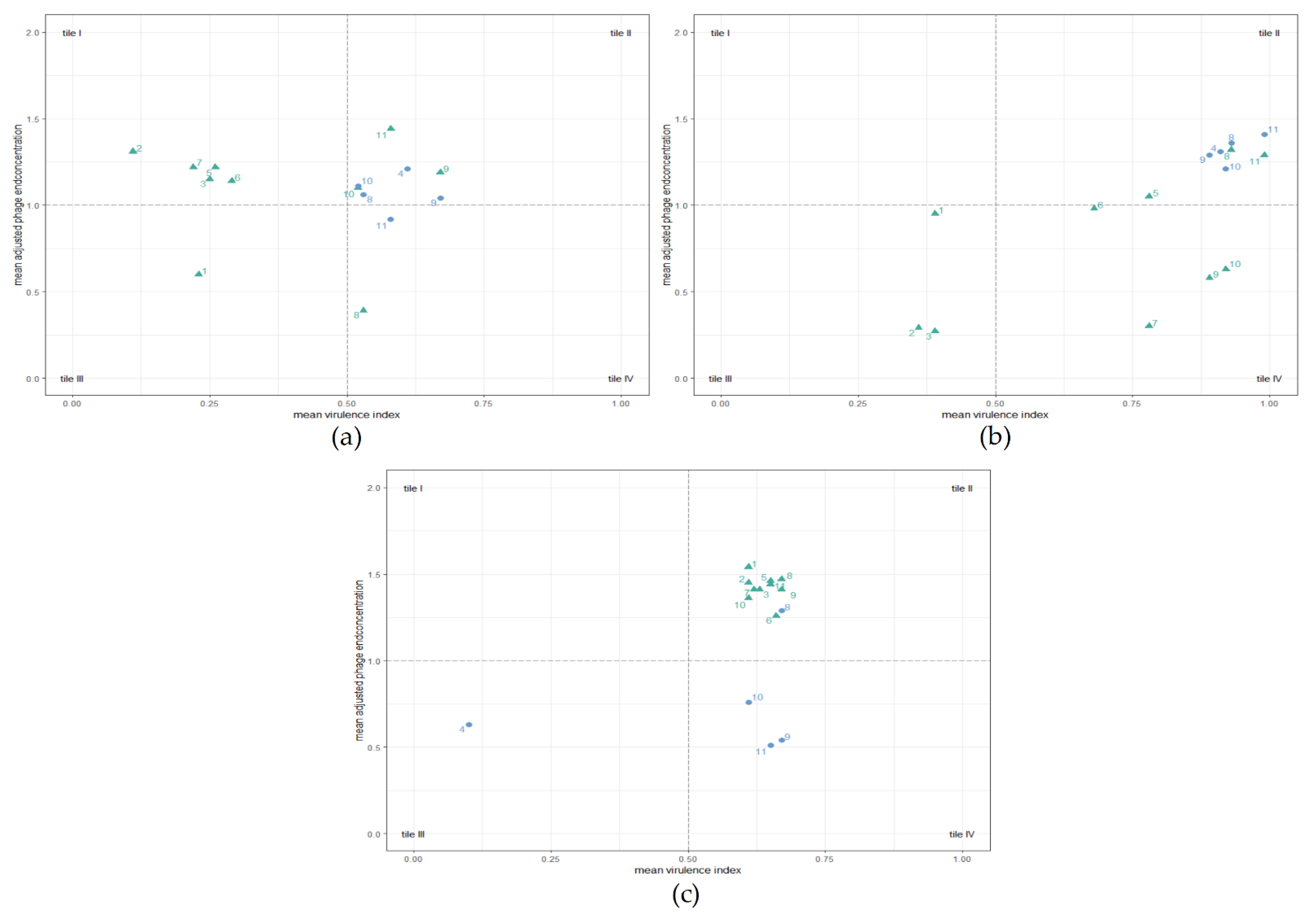
3.3. Phage Testing in Liquid Culture
3.3.1. Planktonic Killing Assay (PKA)
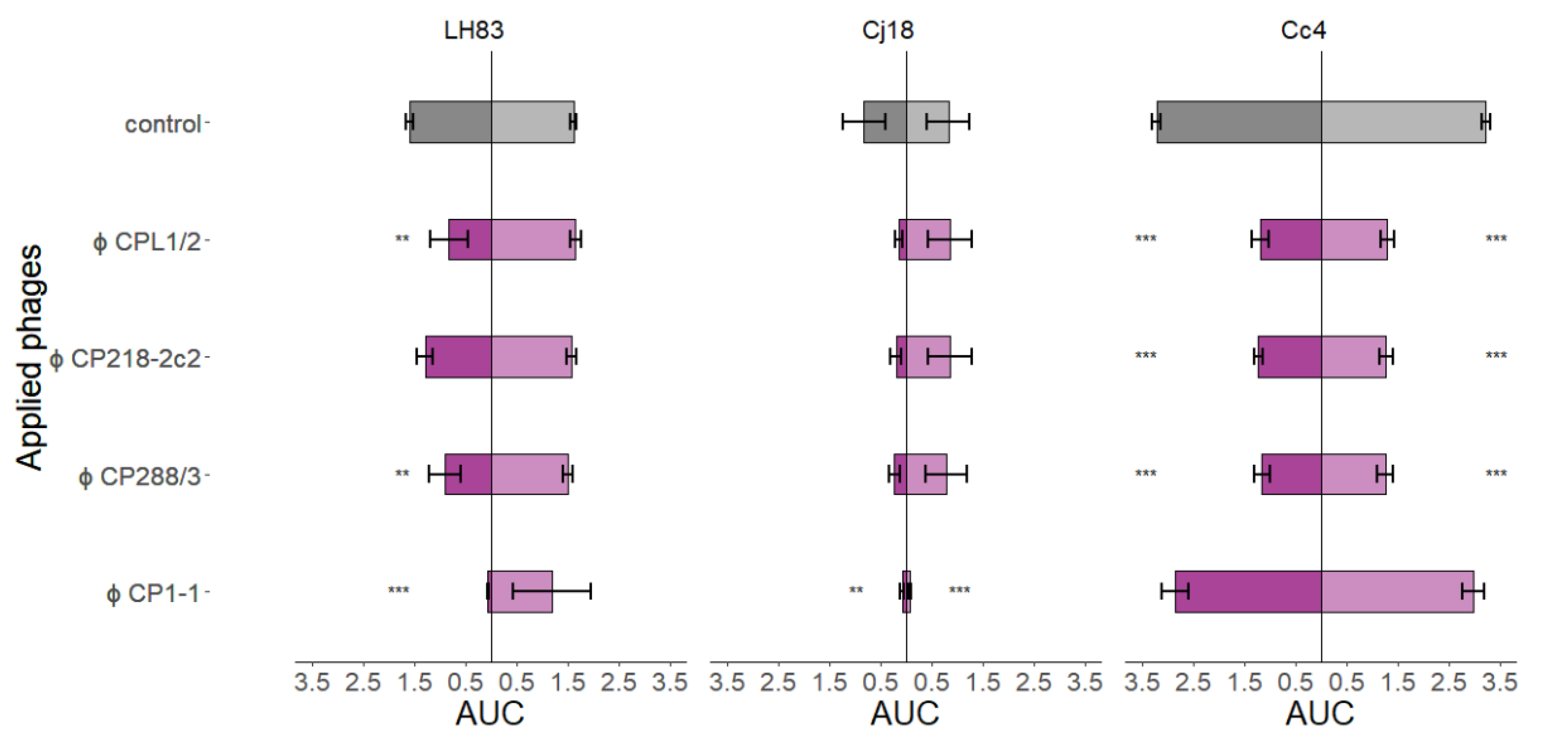
3.3.2. Comparison of AUC Values after Single-Phage Application
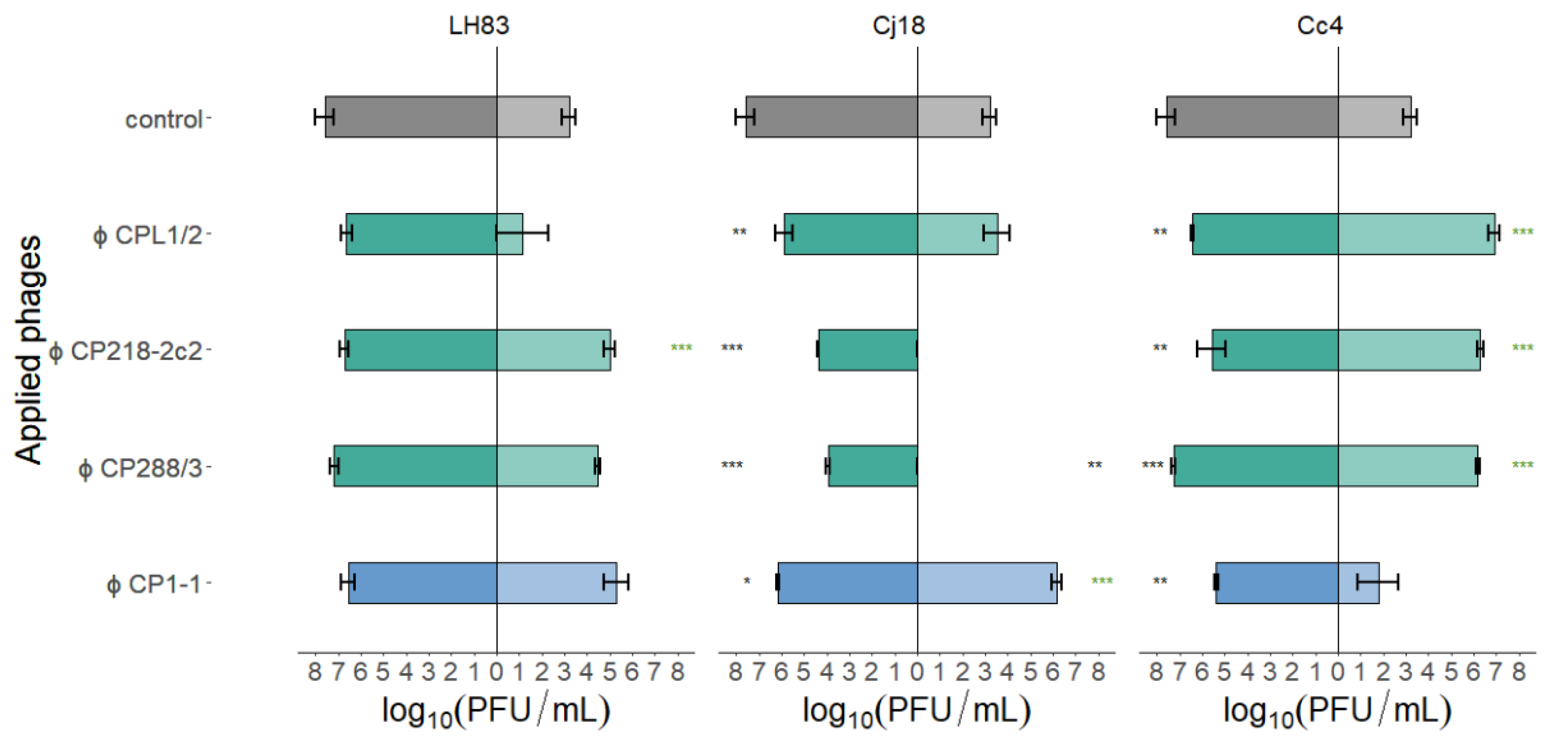
3.3.3. Comparison of Phage Concentrations after 24 h in Single-Phage PKAs
3.3.4. Multi-Phage PKAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- EFSA. The European Union One Health 2019 zoonoses report. EFSA J. 2021, 19, 1–286. [Google Scholar] [CrossRef]
- Skarp, C.P.A.; Hanninen, M.L.; Rautelin, H.I.K. Campylobacteriosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, J.E.; Corcoran, D.; Dooley, J.S.; Fanning, S.; Lucey, B.; Matsuda, M.; McDowell, D.A.; Megraud, F.; Millar, B.C.; O’Mahony, R.; et al. Campylobacter. Vet. Res. 2005, 36, 351–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachamkin, I. Chronic effects of Campylobacter infection. Microbes Infect. 2002, 4, 399–403. [Google Scholar] [CrossRef]
- Mulder, A.C.; Franz, E.; de Rijk, S.; Versluis, M.A.J.; Coipan, C.; Buij, R.; Muskens, G.; Koene, M.; Pijnacker, R.; Duim, B.; et al. Tracing the animal sources of surface water contamination with Campylobacter jejuni and Campylobacter coli. Water Res. 2020, 187, 116421. [Google Scholar] [CrossRef]
- Sasaki, Y.; Maruyama, N.; Zou, B.; Haruna, M.; Kusukawa, M.; Murakami, M.; Asai, T.; Tsujiyama, Y.; Yamada, Y. Campylobacter cross-contamination of chicken products at an abattoir. Zoonoses Public Health 2013, 60, 134–140. [Google Scholar] [CrossRef]
- Cardoso, M.J.; Ferreira, V.; Truninger, M.; Maia, R.; Teixeira, P. Cross-contamination events of Campylobacter spp. in domestic kitchens associated with consumer handling practices of raw poultry. Int. J. Food Microbiol. 2021, 338, 108984. [Google Scholar] [CrossRef]
- Lee, M.D.; Newell, D.G. Campylobacter in poultry: Filling an ecological niche. Avian Dis. 2006, 50, 1–9. [Google Scholar] [CrossRef]
- Luber, P.; Brynestad, S.; Topsch, D.; Scherer, K.; Bartelt, E. Quantification of Campylobacter species cross-contamination during handling of contaminated fresh chicken parts in kitchens. Appl. Environ. Microbiol. 2006, 72, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Teunis, P.F.M.; Marinovic, A.B.; Tribble, D.R.; Porter, C.K.; Swart, A. Acute illness from Campylobacter jejuni may require high doses while infection occurs at low doses. Epidemics 2018, 24, 1–20. [Google Scholar] [CrossRef]
- Kothary, M.H.; Babu, U.S. Infective dose of foodborne pathogens in volunteers: A review. J. Food Safety 2001, 21, 49–73. [Google Scholar] [CrossRef]
- EFSA. Update and review of control options for Campylobacter in broilers at primary production. EFSA J. 2020, 18, e06090. [Google Scholar] [CrossRef]
- Islam, M.R.; Martinez-Soto, C.E.; Lin, J.E.T.; Khursigara, C.M.; Barbut, S.; Anany, H. A systematic review from basics to omics on bacteriophage applications in poultry production and processing. Crit. Rev. Food Sci. 2021, 5, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Haines, M.E.K.; Hodges, F.E.; Nale, J.Y.; Mahony, J.; van Sinderen, D.; Kaczorowska, J.; Alrashid, B.; Akter, M.; Brown, N.; Sauvageau, D.; et al. Analysis of Selection Methods to Develop Novel Phage Therapy Cocktails Against Antimicrobial Resistant Clinical Isolates of Bacteria. Front. Microbiol. 2021, 12, 564. [Google Scholar] [CrossRef] [PubMed]
- Gayder, S.; Parcey, M.; Nesbitt, D.; Castle, A.J.; Svircev, A.M. Population Dynamics between Erwinia amylovora, Pantoea agglomerans and Bacteriophages: Exploiting Synergy and Competition to Improve Phage Cocktail Efficacy. Microorganisms 2020, 8, 1449. [Google Scholar] [CrossRef] [PubMed]
- Schmerer, M.; Molineux, I.J.; Bull, J.J. Synergy as a rationale for phage therapy using phage cocktails. PeerJ 2014, 2, e590. [Google Scholar] [CrossRef] [Green Version]
- Zachary, J.; Storms, Z.J.; Teel, M.R.; Mercurio, K.; Sauvageau, D. The Virulence Index: A Metric for Quantitative Analysis of Phage Virulence. PHAGE Ther. Appl. Res. 2020, 1, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.C.; Wahab, L.; Gill, J.J. Development and validation of a microtiter plate-based assay for determination of bacteriophage host range and virulence. Viruses 2018, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Richards, P.J.; Connerton, P.L.; Connerton, I.F. Phage biocontrol of Campylobacter jejuni in chickens does not produce collateral effects on the gut microbiota. Front. Microbiol. 2019, 10, 476. [Google Scholar] [CrossRef]
- Sails, A.D.; Wareing, D.R.A.; Bolton, F.J.; Fox, A.J.; Curry, A. Characterisation of 16 Campylobacter jejuni and C. coli typing bacteriophages. J. Med. Microbiol. 1998, 47, 123–128. [Google Scholar] [CrossRef]
- Nowaczek, A.; Urban-Chmiel, R.; Dec, M.; Puchalski, A.; Stepien-Pysniak, D.; Marek, A.; Pyzik, E. Campylobacter spp. and bacteriophages from broiler chickens: Characterization of antibiotic susceptibility profiles and lytic bacteriophages. MicrobiologyOpen 2019, 8, e00784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javed, M.A.; Ackermann, H.W.; Azeredo, J.; Carvalho, C.M.; Connerton, I.; Evoy, S.; Hammerl, J.A.; Hertwig, S.; Lavigne, R.; Singh, A.; et al. A suggested classification for two groups of Campylobacter myoviruses. Arch. Virol. 2014, 159, 181–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorensen, M.C.H.; Gencay, Y.E.; Birk, T.; Baldvinsson, S.B.; Jackel, C.; Hammerl, J.A.; Vegge, C.S.; Neve, H.; Brondsted, L. Primary isolation strain determines both phage type and receptors recognised by Campylobacter jejuni bacteriophages. PLoS ONE 2015, 10, e0116287. [Google Scholar] [CrossRef] [Green Version]
- Zampara, A.; Sørensen, M.C.H.; Elsser-Gravesen, A.; Brøndsted, L. Significance of phage-host interactions for biocontrol of Campylobacter jejuni in food. Food Control 2017, 73, 1169–1175. [Google Scholar] [CrossRef]
- Carvalho, C.M.; Gannon, B.W.; Halfhide, D.E.; Santos, S.B.; Hayes, C.M.; Roe, J.M.; Azeredo, J. The in vivo efficacy of two administration routes of a phage cocktail to reduce numbers of Campylobacter coli and Campylobacter jejuni in chickens. BMC Microbiol. 2010, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Kittler, S.; Steffan, S.; Peh, E.; Plotz, M. Phage Biocontrol of Campylobacter: A One Health Approach. Curr. Top. Microbiol. 2021, 431, 127–168. [Google Scholar] [CrossRef]
- Ushanov, L.; Lasareishvili, B.; Janashia, I.; Zautner, A.E. Application of Campylobacter jejuni phages: Challenges and perspectives. Animals 2020, 10, 279. [Google Scholar] [CrossRef] [Green Version]
- GRAS Notice (GRN) No. 966 Preparations Containing Three to Eight Bacteriophages Specific to Campylobacter jejuni. Available online: https://www.fda.gov/media/152868/download (accessed on 22 November 2021).
- Acs, N.; Gambino, M.; Brondsted, L. Bacteriophage Enumeration and Detection Methods. Front. Microbiol. 2020, 11, 2662. [Google Scholar] [CrossRef]
- Anderson, B.; Rashid, M.H.; Carter, C.; Pasternack, G.; Rajanna, C.; Revazishvili, T.; Dean, T.; Senecal, A.; Sulakvelidze, A. Enumeration of bacteriophage particles: Comparative analysis of the traditional plaque assay and real-time QPCR- and nanosight-based assays. Bacteriophage 2011, 1, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Turner, P.E.; Draghi, J.A.; Wilpiszeski, R. High-throughput analysis of growth differences among phage strains. J. Microbiol. Meth. 2012, 88, 117–121. [Google Scholar] [CrossRef]
- Steffan, S.M.; Shakeri, G.; Hammerl, J.A.; Kehrenberg, C.; Peh, E.; Rohde, M.; Jäckel, C.; Plötz, M.; Kittler, S. Isolation and characterization of group III Campylobacter jejuni—Specific bacteriophages from Germany and their potential use in food production. Front. Microbiol. 2021, 12, 3653. [Google Scholar] [CrossRef] [PubMed]
- Korf, I.H.E.; Kittler, S.; Bierbrodt, A.; Mengden, R.; Rohde, C.; Rohde, M.; Kroj, A.; Lehnherr, T.; Fruth, A.; Flieger, A.; et al. In vitro evaluation of a phage cocktail controlling infections with Escherichia coli. Viruses 2020, 12, 1470. [Google Scholar] [CrossRef] [PubMed]
- Valentine, R.C.; Shapiro, B.M.; Stadtman, E.R. Regulation of glutamine synthetase. XII. Electron microscopy of the enzyme from Escherichia coli. Biochemistry 1968, 7, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- Hansen, V.M.; Rosenquist, H.; Baggesen, D.L.; Brown, S.; Christensen, B.B. Characterization of Campylobacter phages including analysis of host range by selected Campylobacter Penner serotypes. BMC Microbiol. 2007, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Jackel, C.; Hammerl, J.A.; Rau, J.; Hertwig, S. A multiplex real-time PCR for the detection and differentiation of Campylobacter phages. PLoS ONE 2017, 12, e0190240. [Google Scholar] [CrossRef] [Green Version]
- Curtright, A.J.; Abedon, S.T. Phage Therapy: Emergent Property Pharmacology. J. Bioanal. Biomed. 2011, S6, 1–13. [Google Scholar] [CrossRef]
- Sandhu, S.K.; Bayliss, C.D.; Morozov, A.Y. How does feedback from phage infections influence the evolution of phase variation in Campylobacter? PLoS Comput. Biol. 2021, 17, e1009067. [Google Scholar] [CrossRef]
- Molina, F.; Simancas, A.; Ramirez, M.; Tabla, R.; Roa, I.; Rebollo, J.E. A New Pipeline for Designing Phage Cocktails Based on Phage-Bacteria Infection Networks. Front. Microbiol. 2021, 12, 149. [Google Scholar] [CrossRef]
- Niu, Y.D.; Hyun, J.E.; Nguyen, N. Bacteriophage Effectiveness for Biocontrol of Foodborne Pathogens Evaluated via High-Throughput Settings. J. Vis. Exp. 2021, 174, e62812. [Google Scholar] [CrossRef]
- Gu, J.M.; Liu, X.H.; Li, Y.; Han, W.Y.; Lei, L.C.; Yang, Y.J.; Zhao, H.L.; Gao, Y.; Song, J.; Lu, R.; et al. A Method for Generation Phage Cocktail with Great Therapeutic Potential. PLoS ONE 2012, 7, e31698. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Phage | Isolation Source | Year |
|---|---|---|---|
| 1 | CP40-2 | caeca | 2017 |
| 2 | CP49-1 | skin | 2017 |
| 3 | CP143-1 | skin | 2017 |
| 4 | CP145-1 | skin | 2017 |
| 5 | CP208-1 | feces | 2017 |
| 6 | CP209-1 | feces | 2017 |
| 7 | CP215-2c1 a | caeca | 2017 |
| 8 | CP215-3 a | caeca | 2017 |
| 9 | CP218-1 b | skin | 2017 |
| 10 | CP218-2c2 b | skin | 2017 |
| 11 | CP218-2c3 b | skin | 2017 |
| 12 | CP218-2c4 b | skin | 2017 |
| 13 | CP253-2c1 | feces | 2017 |
| 14 | CP288/2 c | feces | 2017 |
| 15 | CP288/3 c | feces | 2017 |
| 16 | CPL1/1 | n/a | n/a |
| 17 | CPL1/2 | n/a | n/a |
| 18 | CPL2 | n/a | n/a |
| Campylobacter Field Isolate | EOP Values of Campylobacter Phages | |||
|---|---|---|---|---|
| CPL1/2 | CP218-2c2 | CP288/3 | CP1-1 | |
| C. jejuni LH83 | 0 | 0.9 | 0.6 | 0.6 |
| C. jejuni Cj18 | 0.4 | 0 | 0 | 1.0 |
| C. coli Cc4 | 1.3 | 1.0 | 1.0 | 0 |
| Campylobacter Phage | ||||
|---|---|---|---|---|
| CPL1/2 | CP218-2c2 | CP288/3 | CP1-1 | |
| Genome length by PFGE (kb) | ~175 | ~175 | ~174 | ~145 |
| HhaI sensitivity | − | − | − | + |
| SwaI sensitivity | + | + | + | not tested |
| Dimensions Based on Electron Micrographs (n = 5) | ||||
| Mean tail length (nm) | 122.49 ± 5.01 | 133.46 ± 2.19 | 139.18 ± 1.56 | 112.21 ± 1.21 |
| Mean head diameter (nm) | 97.49 ± 2.61 | 91.41 ± 3.58 | 101.04 ± 8.41 | 97.79 ± 2.4 |
| Mean head length (nm) | 103.2 ± 8.05 | 108.28 ± 8.01 | 104.86 ± 3.88 | 108.34 ± 2.76 |
| Virus family | Myoviridae | Myoviridae | Myoviridae | Myoviridae |
| qPCR | group II | group II | group II | group III |
| Mean Plaque Diameter (24 h, 0.7% overlay, n = 200) (mm) | ||||
| 0.64 ± 0.07 | 0.75 ± 0.08 | 0.93 ± 0.1 | 1.13 ± 0.34 | |
| Campylobacter Phage | Starting Concentrations c0 (log10(PFU/mL)) | |
|---|---|---|
| MOIinput 10 | MOIinput 0.001 | |
| CP1-1 | 7.2 ± 0.1 * | 3.5 ± 0.0 ** |
| CPL/1/2 | 8.2 ± 0.8 * | 3.0 ± 0.2 ** |
| CP218-2c2 | 7.6 ± 0.6 * | 2.9 ± 0.2 ** |
| CP288/3 | 7.3 ± 0.1 * | 3.4 ± 0.1 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steffan, S.M.; Shakeri, G.; Kehrenberg, C.; Peh, E.; Rohde, M.; Plötz, M.; Kittler, S. Campylobacter Bacteriophage Cocktail Design Based on an Advanced Selection Scheme. Antibiotics 2022, 11, 228. https://doi.org/10.3390/antibiotics11020228
Steffan SM, Shakeri G, Kehrenberg C, Peh E, Rohde M, Plötz M, Kittler S. Campylobacter Bacteriophage Cocktail Design Based on an Advanced Selection Scheme. Antibiotics. 2022; 11(2):228. https://doi.org/10.3390/antibiotics11020228
Chicago/Turabian StyleSteffan, Severin Michael, Golshan Shakeri, Corinna Kehrenberg, Elisa Peh, Manfred Rohde, Madeleine Plötz, and Sophie Kittler. 2022. "Campylobacter Bacteriophage Cocktail Design Based on an Advanced Selection Scheme" Antibiotics 11, no. 2: 228. https://doi.org/10.3390/antibiotics11020228
APA StyleSteffan, S. M., Shakeri, G., Kehrenberg, C., Peh, E., Rohde, M., Plötz, M., & Kittler, S. (2022). Campylobacter Bacteriophage Cocktail Design Based on an Advanced Selection Scheme. Antibiotics, 11(2), 228. https://doi.org/10.3390/antibiotics11020228