
Nosocomial Outbreak of Extensively Drug-Resistant (Polymyxin B and Carbapenem) Klebsiella pneumoniae in a Collapsed University Hospital Due to COVID-19 Pandemic


 ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Clinical Data and Context
2.3. Identification and Antimicrobial Susceptibility Testing of Isolates
2.4. DNA Extraction and Genome Sequencing
2.5. In Silico Identification of Carbapenem and Polymyxin B Resistance Genes and Characterization of Bacterial Genomic Relationship
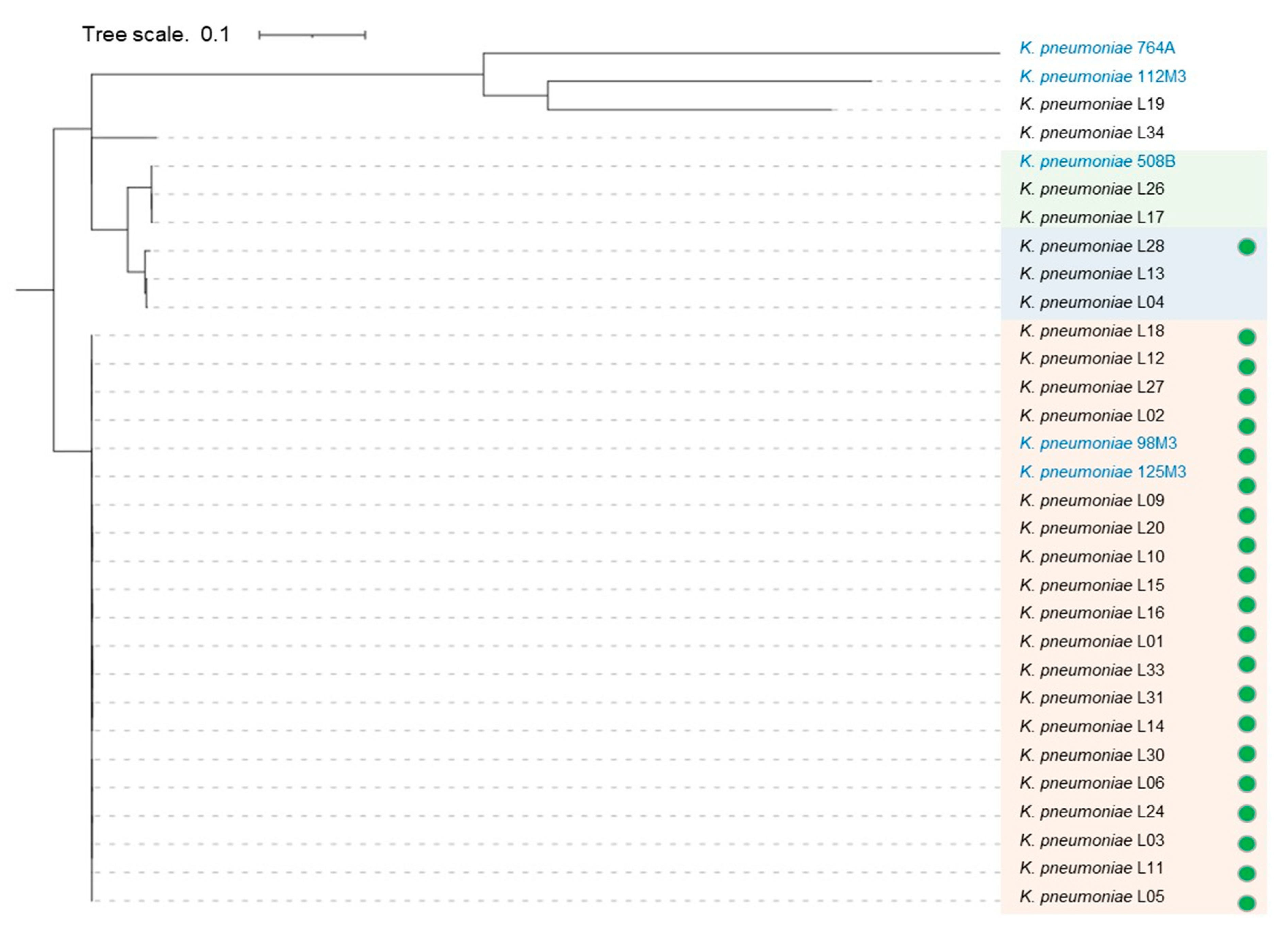
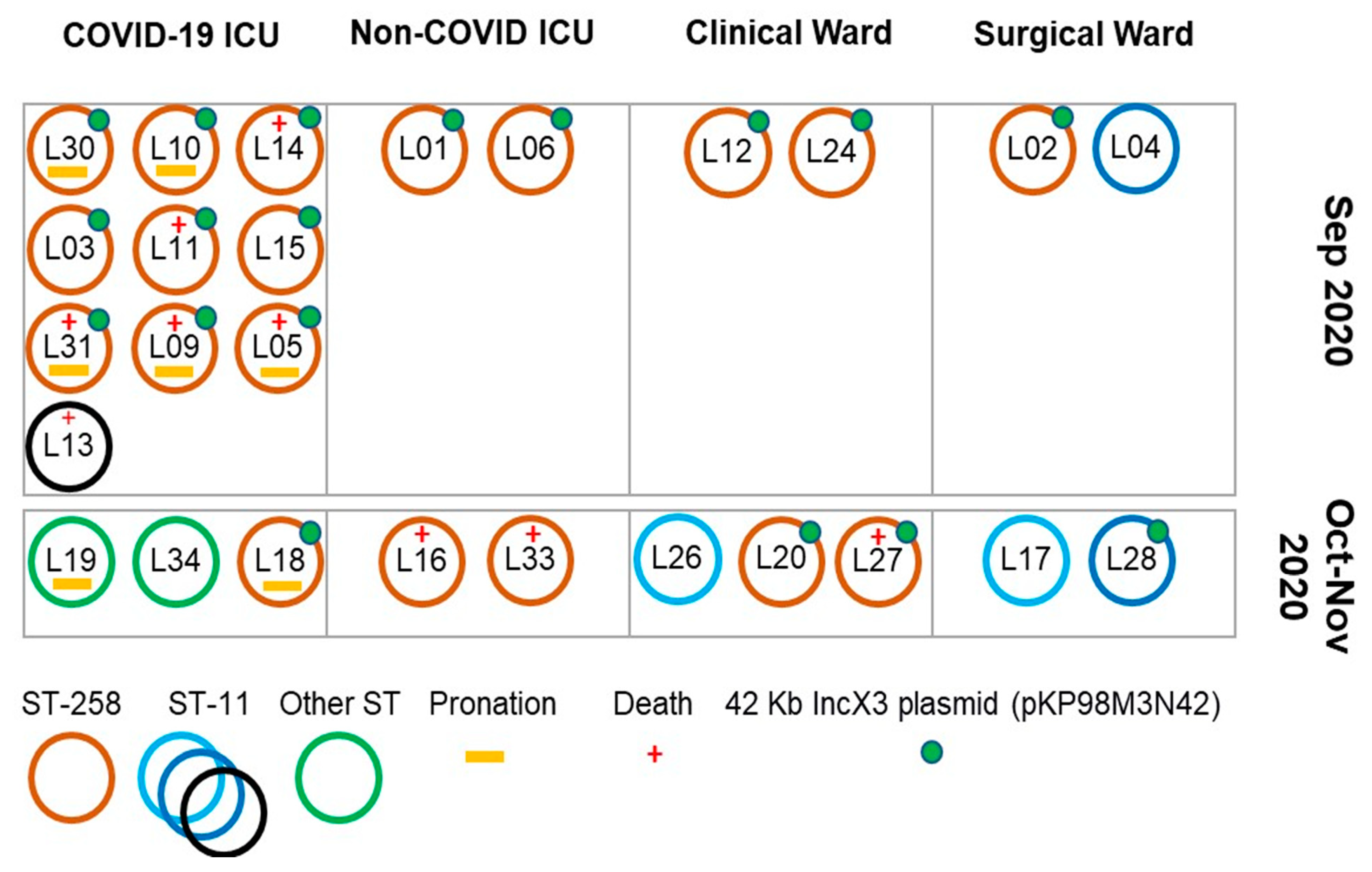
3. Results
3.1. K. pneumoniae Clinical Isolates Features
3.2. Clinical Features and Patients’ Outcomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ranzani, O.T.; Bastos, L.S.L.; Gelli, J.G.M.; Marchesi, J.F.; Baião, F.; Hamacher, S.; Bozza, F.A. Characterisation of the First 250,000 Hospital Admissions for COVID-19 in Brazil: A Retrospective Analysis of Nationwide Data. Lancet Respir. Med. 2021, 9, 407–418. [Google Scholar] [CrossRef]
- Murray, A.K. The Novel Coronavirus COVID-19 Outbreak: Global Implications for Antimicrobial Resistance. Front. Microbiol. 2020, 11, 1020. [Google Scholar] [CrossRef] [PubMed]
- Cantón, R.; Gijón, D.; Ruiz-Garbajosa, P. Antimicrobial Resistance in ICUs: An Update in the Light of the COVID-19 Pandemic. Curr. Opin. Crit. Care 2020, 26, 433–441. [Google Scholar] [CrossRef]
- Segala, F.V.; Bavaro, D.F.; di Gennaro, F.; Salvati, F.; Marotta, C.; Saracino, A.; Murri, R.; Fantoni, M. Impact of SARS-CoV-2 Epidemic on Antimicrobial Resistance: A Literature Review. Viruses 2021, 13, 2110. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, G.G.; Ferreira, L.R.; Feliciano, C.S.; Campos Júnior, C.P.; Molina, F.M.R.; Vendruscolo, A.C.S.; Bradan, G.M.A.; Lopes, N.A.P.; Martinez, R.; Bollela, V.R. Pre- and Post-COVID-19 Evaluation of Antimicrobial Susceptibility for Healthcare-Associated Infections in the Intensive Care Unit of a Tertiary Hospital. Rev. Soc. Bras. Med. Trop. 2021, 54. [Google Scholar] [CrossRef] [PubMed]
- Dalmolin, T.V.; Carneiro, M.D.S.; de Castro, L.P.; Volpato, F.C.Z.; Wink, P.L.; de Lima-Morales, D.; Barth, A.L. Evaluation of the Susceptibility Test of Polymyxin B Using the Commercial Test Policimbac®. Braz. J. Microbiol. 2020, 51, 1135–1137. [Google Scholar] [CrossRef]
- EUCAST. Antimicrobial Susceptibility Testing. Available online: https://www.eucast.org/ast_of_bacteria/ (accessed on 9 April 2022).
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.-M. ARG-ANNOT, a New Bioinformatic Tool to Discover Antibiotic Resistance Genes in Bacterial Genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Wright, M.S.; Suzuki, Y.; Jones, M.B.; Marshall, S.H.; Rudin, S.D.; van Duin, D.; Kaye, K.; Jacobs, M.R.; Bonomo, R.A.; Adams, M.D. Genomic and Transcriptomic Analyses of Colistin-Resistant Clinical Isolates of Klebsiella Pneumoniae Reveal Multiple Pathways of Resistance. Antimicrob. Agents Chemother. 2015, 59, 536–543. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, M.R.; McCulloch, J.A.; Vianello, M.A.; Moura, Q.; Pérez-Chaparro, P.J.; Esposito, F.; Sartori, L.; Dropa, M.; Matté, M.H.; Lira, D.P.A.; et al. First Report of the Globally Disseminated IncX4 Plasmid Carrying the Mcr-1 Gene in a Colistin-Resistant Escherichia Coli Sequence Type 101 Isolate from a Human Infection in Brazil. Antimicrob. Agents Chemother. 2016, 60, 6415–6417. [Google Scholar] [CrossRef] [Green Version]
- Campos, P.A.D.; Fuga, B.; Ferreira, M.L.; Brígido, R.T.E.S.; Lincopan, N.; Gontijo-Filho, P.P.; Ribas, R.M. Genetic Alterations Associated with Polymyxin B Resistance in Nosocomial KPC-2-Producing Klebsiella Pneumoniae from Brazil. Microb. Drug Resist. 2021, 27, 1677–1684. [Google Scholar] [CrossRef]
- Pasteur Institute. Klebsiella Sequence Typing. Available online: https://bigsdb.pasteur.fr/klebsiella/ (accessed on 9 April 2022).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Micro-Biol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borelli, T.C.; Lovate, G.L.; Scaranello, A.F.T.; Ribeiro, L.F.; Zaramela, L.; Pereira-dos-Santos, F.M.; Silva-Rocha, R.; Guazzaroni, M.-E. Combining Functional Genomics and Whole-Genome Sequencing to Detect Antibiotic Resistance Genes in Bacterial Strains Co-Occurring Simultaneously in a Brazilian Hospital. Antibiotics 2021, 10, 419. [Google Scholar] [CrossRef] [PubMed]
- Tiri, B.; Sensi, E.; Marsiliani, V.; Cantarini, M.; Priante, G.; Vernelli, C.; Martella, L.A.; Costantini, M.; Mariottini, A.; Andreani, P.; et al. Antimicrobial Stewardship Program, COVID-19, and Infection Control: Spread of Car-bapenem-Resistant Klebsiella Pneumoniae Colonization in ICU COVID-19 Patients. What Did Not Work? J. Clin. Med. 2020, 9, 2744. [Google Scholar] [CrossRef] [PubMed]
- Adebisi, Y.A.; Alaran, A.J.; Okereke, M.; Oke, G.I.; Amos, O.A.; Olaoye, O.C.; Oladunjoye, I.; Olanrewaju, A.Y.; Ukor, N.A.; Lucero-Prisno, D.E. COVID-19 and Antimicrobial Resistance: A Review. Infectious Diseases. Res. Treat. 2021, 14, 117863372110338. [Google Scholar] [CrossRef]
- Ginsburg, A.S.; Klugman, K.P. COVID-19 Pneumonia and the Appropriate Use of Antibiotics. Lancet Glob. Health 2020, 8, e1453–e1454. [Google Scholar] [CrossRef]
- Zavala-Flores, E.; Salcedo-Matienzo, J. Medicación Prehospitalaria En Pacientes Hospitalizados Por COVID-19 En Un Hospital Público de Lima-Perú. Acta Med. Peru. 2020, 37, 393–395. [Google Scholar] [CrossRef]
- Andrade, L.N.; Vitali, L.; Gaspar, G.G.; Bellissimo-Rodrigues, F.; Martinez, R.; Darini, A.L.C. Expansion and Evolution of a Virulent, Extensively Drug-Resistant (Polymyxin B-Resistant), QnrS1-, CTX-M-2-, and KPC-2-Producing Klebsiella Pneumoniae ST11 International High-Risk Clone. J. Clin. Microbiol. 2014, 52, 2530–2535. [Google Scholar] [CrossRef] [Green Version]
- Andrade, L.N.; Curiao, T.; Ferreira, J.C.; Longo, J.M.; Clímaco, E.C.; Martinez, R.; Bellissimo-Rodrigues, F.; Basile-Filho, A.; Evaristo, M.A.; del Peloso, P.F.; et al. Dissemination of Bla KPC-2 by the Spread of Klebsiella Pneumoniae Clonal Complex 258 Clones (ST258, ST11, ST437) and Plasmids (IncFII, IncN, IncL/M) among Enterobacteriaceae Species in Brazil. Antimicrob. Agents Chemother. 2011, 55, 3579–3583. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, G.G.; Bellissimo-Rodrigues, F.; de Andrade, L.N.; Darini, A.L.; Martinez, R. Induction and Nosocomial Dissemination of Carbapenem and Polymyxin-Resistant Klebsiella Pneumoniae. Rev. Soc. Bras. Med. Trop. 2015, 48, 483–487. [Google Scholar] [CrossRef] [Green Version]
- Cerdeira, L.T.; Cunha, M.P.V.; Francisco, G.R.; Bueno, M.F.C.; Araujo, B.F.; Ribas, R.M.; Gontijo-Filho, P.P.; Knöbl, T.; de Oliveira Garcia, D.; Lincopan, N. IncX3 Plasmid Harboring a Non-Tn 4401 Genetic Element (NTE KPC ) in a Hospital-Associated Clone of KPC-2-Producing Klebsiella Pneumoniae ST340/CG258. Diagn. Microbiol. Infec Tious Dis. 2017, 89, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Tofteland, S.; Naseer, U.; Lislevand, J.H.; Sundsfjord, A.; Samuelsen, Ø. A Long-Term Low-Frequency Hospital Outbreak of KPC-Producing Klebsiella Pneumoniae Involving Intergenus Plasmid Diffusion and a Persisting Envi-ronmental Reservoir. PLoS ONE 2013, 8, e59015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrey, D.O.; Pereira Dantas, P.; Martins, W.B.; De Carvalho, F.M.; Almeida, L.G.P.; Sands, K.; Portal, E.; Sauser, J.; Cayô, R.; Nicolas, M.F.; et al. An Emerging Clone, Klebsiellapneumoniae Carbapenemase 2–Producing K. Pneumoniae Sequence Type 16, Associated With High Mortality Rates in a CC258-Endemic Setting. Clin. Infect. Dis. 2020, 71, e141–e150. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Chen, Q.; Shen, F.; Jiang, Y.; Wu, X.; Hua, X.; Fu, Y.; Yu, Y. Resistance Evolution of Hypervirulent Car-bapenem-Resistant Klebsiella Pneumoniae ST11 during Treatment with Tigecycline and Polymyxin. Emerg. Microbes Infect. 2021, 10, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Cannatelli, A.; D’Andrea, M.M.; Giani, T.; di Pilato, V.; Arena, F.; Ambretti, S.; Gaibani, P.; Rossolini, G.M. In Vivo Emergence of Colistin Resistance in Klebsiella Pneumoniae Producing KPC-Type Carbapenemases Mediated by in-sertional Inactivation of the PhoQ/PhoP MgrB Regulator. Antimicrob. Agents Chemother. 2013, 57, 5521–5526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Mechanisms of Polymyxin Resistance: Acquired and Intrinsic Resistance in Bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Period | Clinical Setting | Date | ID | ST | PB MIC (µg/mL) | Mutated Genes Related to PB Resistance | |
|---|---|---|---|---|---|---|---|
| Policimbac | Broth Microdilution | ||||||
| September 2020 | COVID ICU | 14 | L14 | 258 | ≥64 | 16 | non identified |
| 14 | L03 | 258 | 32 | 64 | mgrB, pmrA | ||
| 14 | L13 | 11 | ≥64 | 32 | mgrB, pmrA, pmrD | ||
| 17 | L11 | 258 | 32 | 32 | pmrA | ||
| 18 | L10 | 258 | 16 | nd | mgrB, pmrA | ||
| 20 | L30 | 258 | ≥64 | ≤0.5 | pmrA, pmrC | ||
| 21 | L09 | 258 | ≥64 | 32 | mgrB, pmrA | ||
| 21 | L05 | 258 | 4 | 8 | pmrA | ||
| 25 | L31 | 258 | ≥64 | 8 | pmrA | ||
| 30 | L15 | 258 | ≥64 | 4 | pmrA | ||
| Non-COVID ICU | 14 | L01 | 258 | 8 | 16 | pmrA | |
| 24 | L06 | 258 | ≥64 | 32 | pmrA | ||
| Clinical Ward | 5 | L12 | 258 | ≥64 | 2 | pmrA | |
| Surgical Ward | 14 | L02 | 258 | 64 | 256 | pmrA | |
| 17 | L04 | 11 | 8 | ≤0.5 | pmrA, pmrD, ramA | ||
| October 2020 | COVID ICU | 1 | L34 | 437 | ≥64 | 32 | pmrA, pmrD, ramA |
| 29 | L18 | 258 | ≥64 | 64 | pmrA | ||
| Non-COVID ICU | 5 | L33 | 258 | 64 | 8 | pmrA | |
| 29 | L16 | 258 | 32 | 4 | pmrA | ||
| Clinical Ward | 8 | L27 | 258 | 4 | 2 | pmrA | |
| 24 | L24 | 258 | 4 | ≤0.5 | pmrA | ||
| 3 | L26 | 11 | ≥64 | 128 | mgrB, pmrA, pmrD | ||
| Surgical Ward | 10 | L28 | 11 | 16 | 256 | mgrB, pmrA, pmrD | |
| 29 | L17 | 11 | ≥64 | 64 | mgrB, pmrA, pmrD, pmrK | ||
| November 2020 | COVID ICU | 7 | L19 | 16 | 32 | 16 | pmrA, pmrC, pmrD, pmrH, pmrI, pmrJ, pmrK, pmrL, ramA |
| Clinical Ward | 5 | L20 | 258 | 32 | 8 | mgrB, pmrA | |
| Demographics Characteristics | Results |
|---|---|
| Mean age (Min-Max) | 57.8 (21–85) |
| Gender (n = 26) | n (%) |
| Male | 15 (57.6) |
| Female | 11 (42.3) |
| Admission Unit (n = 26) | n (%) |
| COVID ICU | 13 (50.2) |
| Non-COVID ICU | 4 (15.3) |
| Clinical Ward | 5 (19.2) |
| Surgical Ward | 4 (15.3) |
| Hospitalization days median (max-min) | 49 (110–8) |
| Comorbidities (n = 26) | n (%) |
| Systemic Arterial Hypertension (SAH) | 10 (40) |
| Diabetes Mellitus (DM) | 4 (16) |
| Obesity | 2 (8) |
| Other diseases | 9 (36) |
| Healthcare-associated infections (n = 18) | n (%) |
| Bloodstream infection | 6 (33.3) |
| Ventilator-Associated Pneumonia | 5 (27.8) |
| Urinary Tract Infection | 3 (16.6) |
| Surgical Site Infection | 2 (11.1) |
| Peritonitis | 1 (5.6) |
| Tracheitis | 1 (5.6) |
| COVID-19 infection | n (%) |
| Positive | 19 (73) |
| Negative | 6 (23) |
| Inconclusive | 1 (4) |
| Passage by the intensive care unit n (%) | 21 (80.7) |
| Pronation n (%) | 7 (26.9) |
| Previous use of polymyxin n (%) | 13 (50) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaspar, G.G.; Tamasco, G.; Abichabki, N.; Scaranello, A.F.T.; Auxiliadora-Martins, M.; Pocente, R.; Andrade, L.N.; Guazzaroni, M.-E.; Silva-Rocha, R.; Bollela, V.R. Nosocomial Outbreak of Extensively Drug-Resistant (Polymyxin B and Carbapenem) Klebsiella pneumoniae in a Collapsed University Hospital Due to COVID-19 Pandemic. Antibiotics 2022, 11, 814. https://doi.org/10.3390/antibiotics11060814
Gaspar GG, Tamasco G, Abichabki N, Scaranello AFT, Auxiliadora-Martins M, Pocente R, Andrade LN, Guazzaroni M-E, Silva-Rocha R, Bollela VR. Nosocomial Outbreak of Extensively Drug-Resistant (Polymyxin B and Carbapenem) Klebsiella pneumoniae in a Collapsed University Hospital Due to COVID-19 Pandemic. Antibiotics. 2022; 11(6):814. https://doi.org/10.3390/antibiotics11060814
Chicago/Turabian StyleGaspar, Gilberto G., Gustavo Tamasco, Nathália Abichabki, Ana Flavia T. Scaranello, Maria Auxiliadora-Martins, Renata Pocente, Leonardo N. Andrade, María-Eugenia Guazzaroni, Rafael Silva-Rocha, and Valdes R. Bollela. 2022. "Nosocomial Outbreak of Extensively Drug-Resistant (Polymyxin B and Carbapenem) Klebsiella pneumoniae in a Collapsed University Hospital Due to COVID-19 Pandemic" Antibiotics 11, no. 6: 814. https://doi.org/10.3390/antibiotics11060814
APA StyleGaspar, G. G., Tamasco, G., Abichabki, N., Scaranello, A. F. T., Auxiliadora-Martins, M., Pocente, R., Andrade, L. N., Guazzaroni, M. -E., Silva-Rocha, R., & Bollela, V. R. (2022). Nosocomial Outbreak of Extensively Drug-Resistant (Polymyxin B and Carbapenem) Klebsiella pneumoniae in a Collapsed University Hospital Due to COVID-19 Pandemic. Antibiotics, 11(6), 814. https://doi.org/10.3390/antibiotics11060814