
Novel 1,2,3-Triazole-sulphadiazine-ZnO Hybrids as Potent Antimicrobial Agents against Carbapenem Resistant Bacteria

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
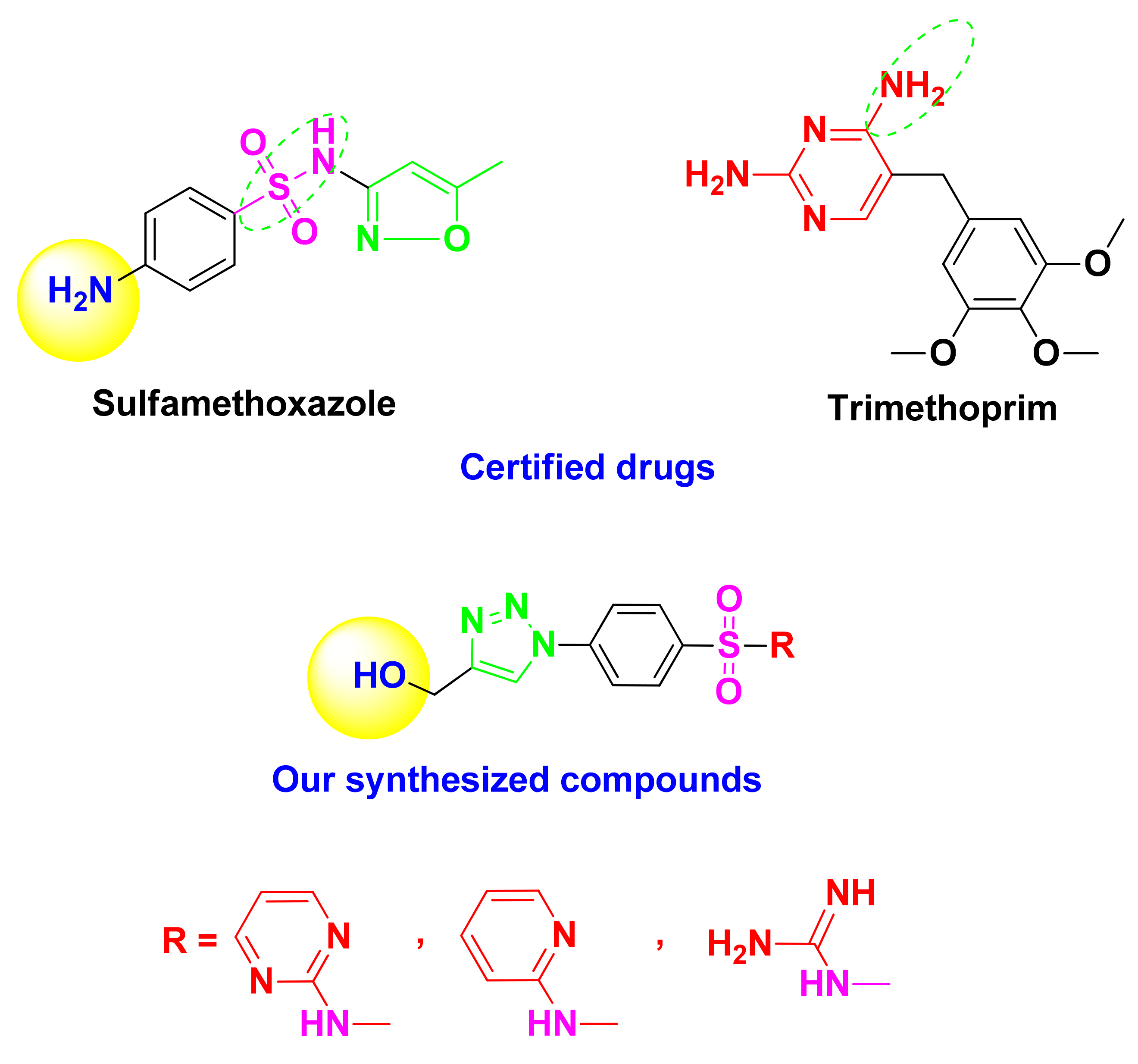
Rational Study
2. Results and Discussion
2.1. Chemistry
2.1.1. Organic Synthetic Part
2.1.2. Characterization of ZnO Nanoparticles
X-ray Diffractometer Analysis
FT-IR Spectroscopy
2.1.3. Molecular Modeling
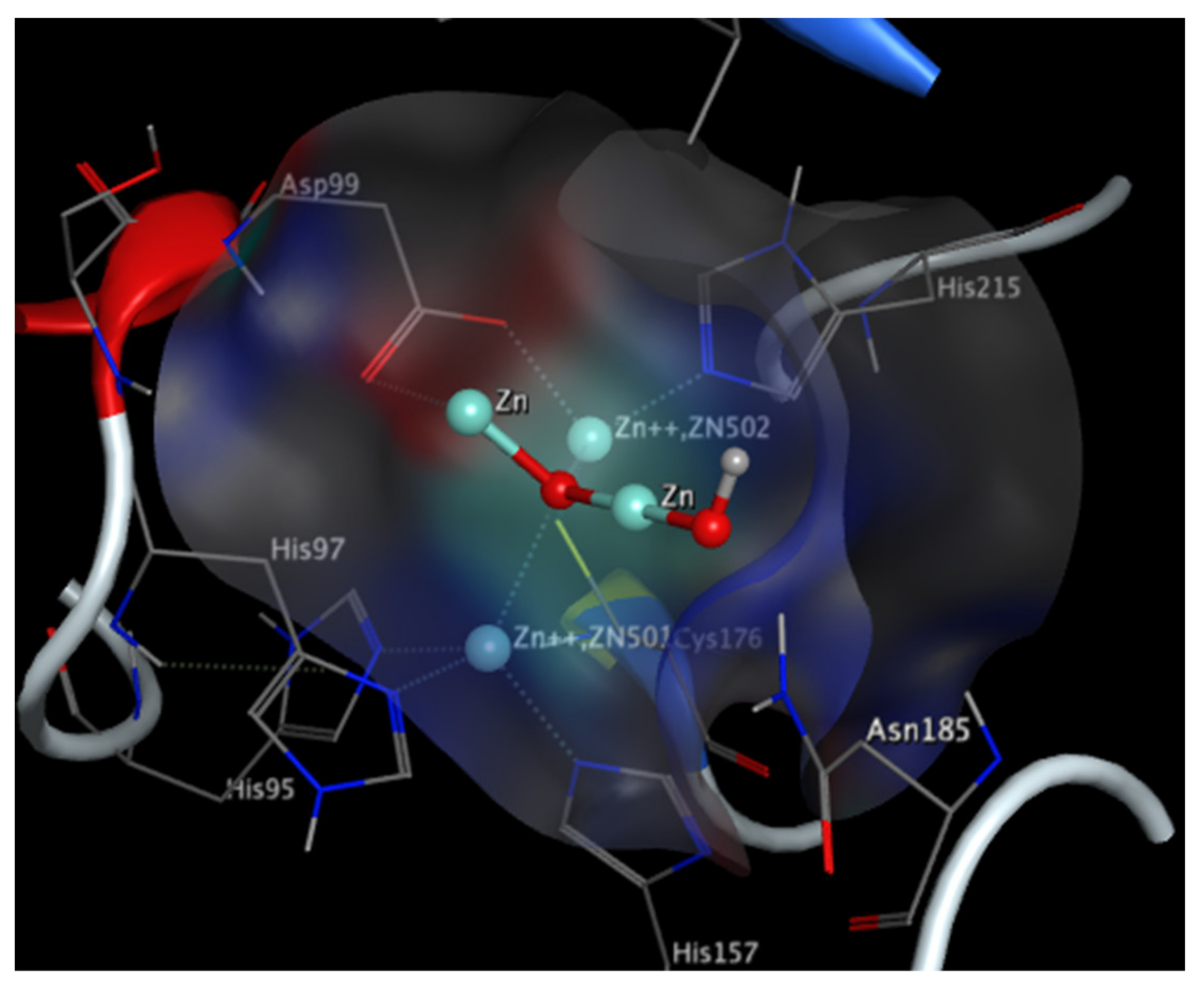
Imipenemase-1 (IMP-1) Enzyme
New Delhi Metallo-β-Lactamase (NDM-1) Enzyme
2.2. In Vitro Analyses
2.2.1. Detection of the IPM and NDM Genes Presence in the Bacterial Isolates
2.2.2. MBLs Inhibition Activity against IMP-1 and NDM-1
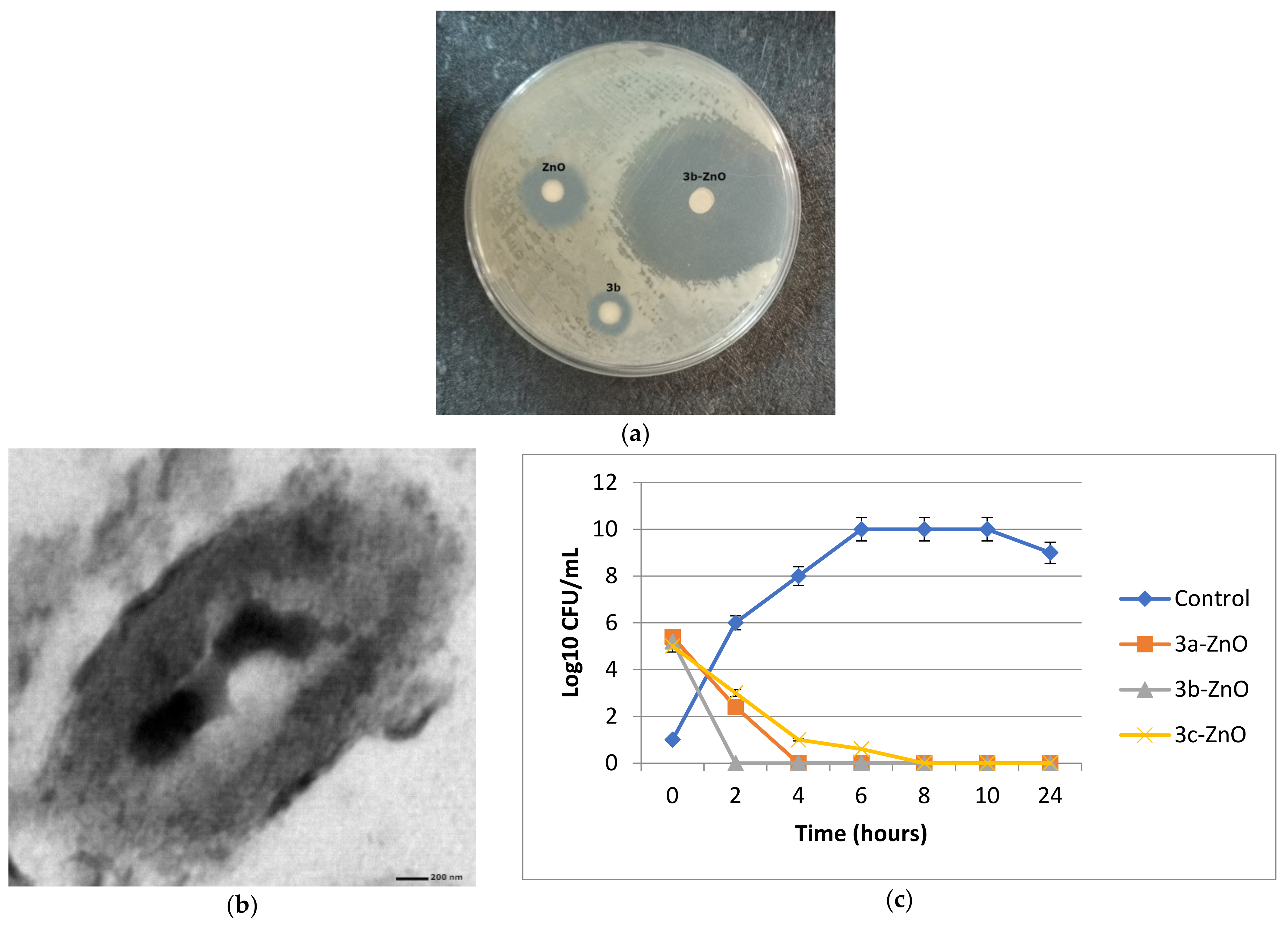
2.2.3. Antibacterial Activity against Metallo-β-Lactamase Producing K. pneumoniae
2.2.4. Cytotoxicity Study
2.3. In Vivo Studies
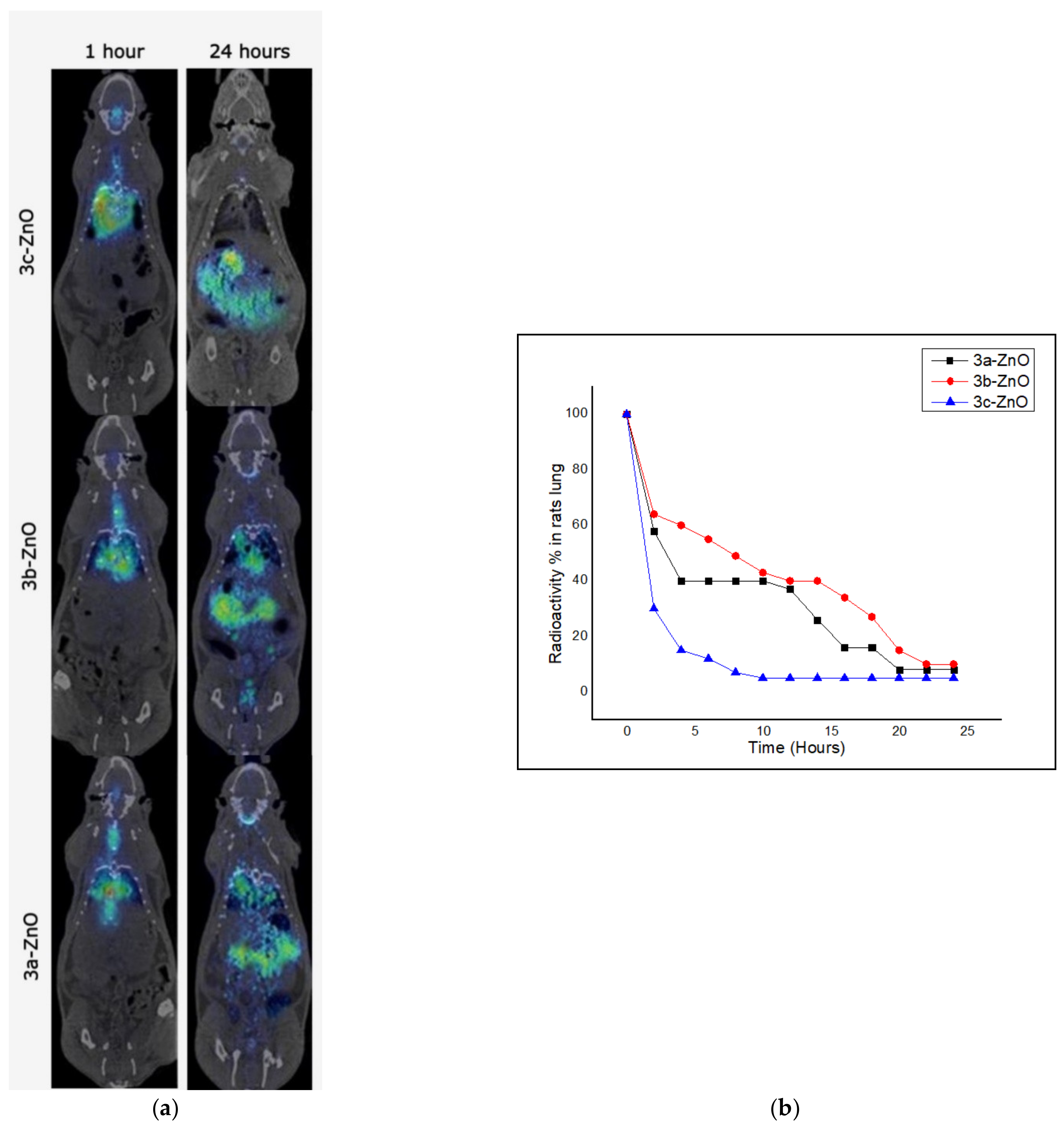
2.3.1. Biodistribution of the 1,2,3-Triazole-sulphadiazine-ZnO Hybrids in Lungs of Uninfected Rats
2.3.2. Residence Time of the 1,2,3-Triazole-sulphadiazine-ZnO Hybrids in Lungs of Uninfected Rats
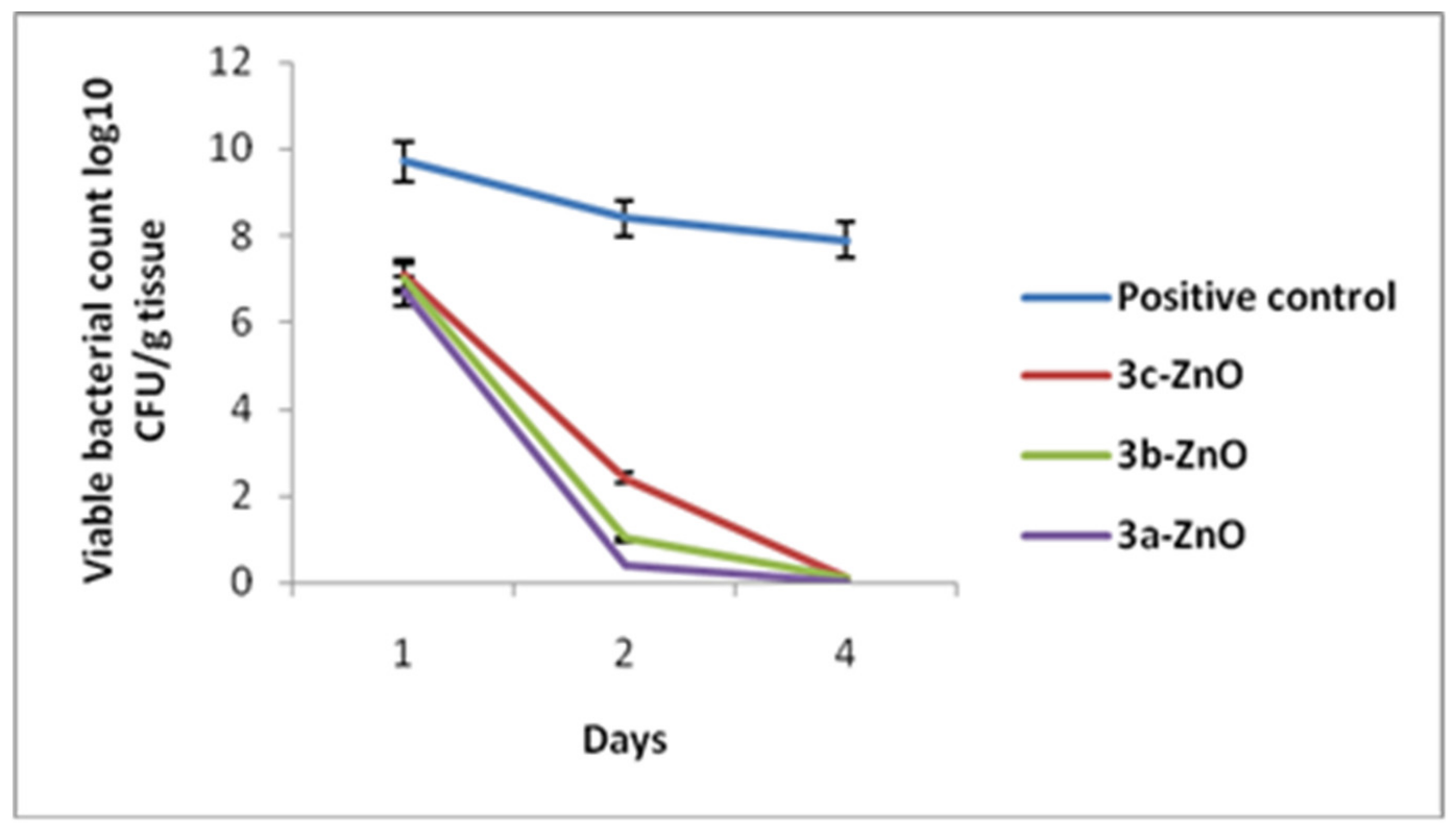
2.3.3. Bacterial Load Assessment
2.3.4. Histopathological Investigations
Light Microscopic Investigations
- (a)
- First Interval
- (b)
- Second Interval
Transmission Electron Microscopical Study
3. Materials and Methods
3.1. Chemistry
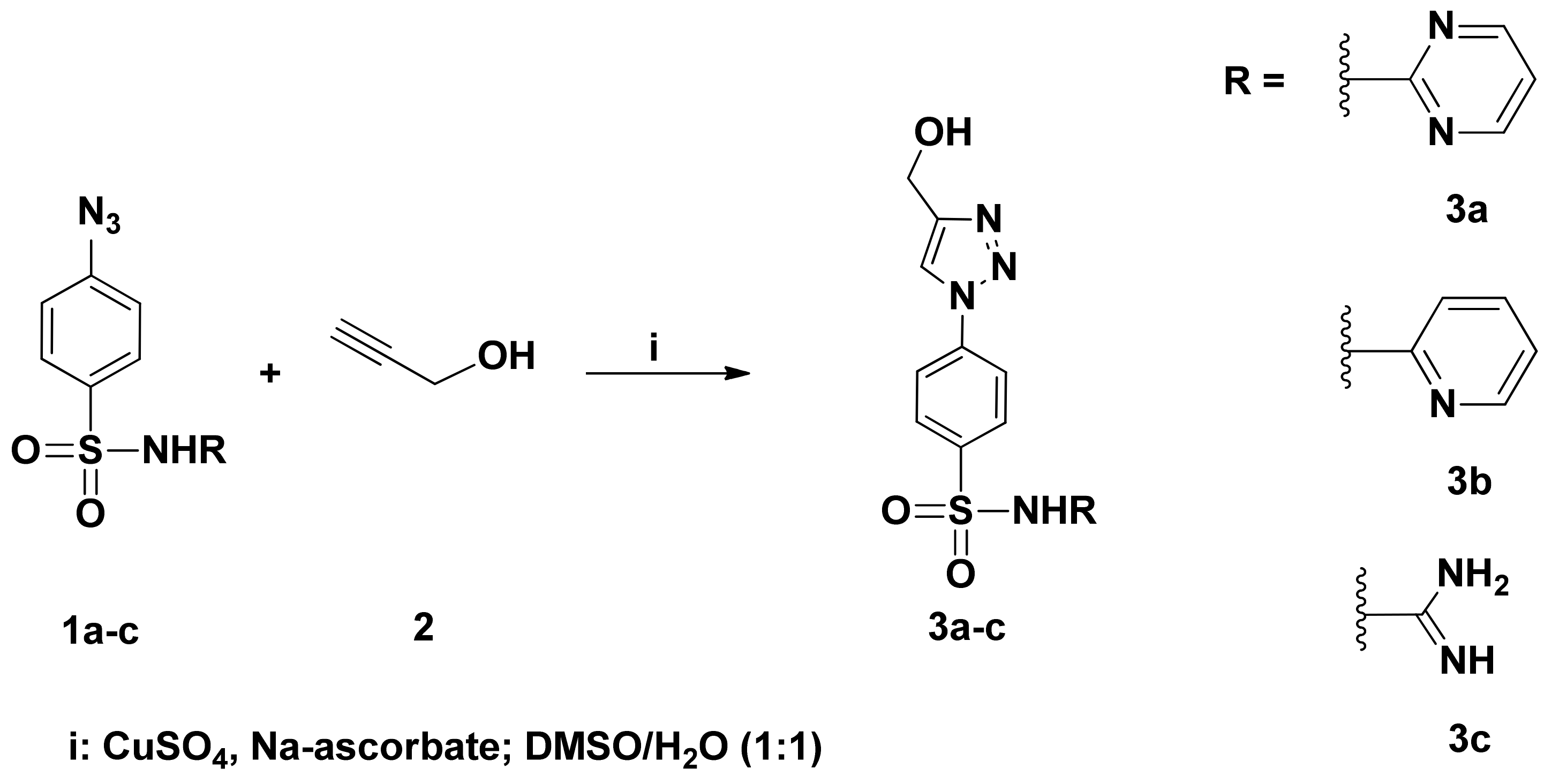
3.1.1. Synthesis of 1,2,3-Triazoles Bearing Sulfa-Drug 3a−c
3.1.2. Synthesis of ZnO Nanoparticles
3.1.3. Combination between 1,2,3-Triazole-sulphadiazine and ZnO NPs
3.1.4. Molecular Modeling
Bacterial Enzymes Preparation
Database Generation and Optimization
3.2. Biological Evaluation
3.2.1. Microorganisms
3.2.2. In Vitro Analyses
Determination of the Metallo-β-Lactamases Production Using Molecular Technique
In Vitro MBLs Inhibition Activity against IMP-1 and NDM-1
Antibacterial Activity of the Prepared Compounds
Cytotoxicity Study in Lung (BEAS-2Bs) Cell Line
3.2.3. In Vivo Studies
Animals
Ethics
Endotracheal Aerosolization of 3a-ZnO, 3b-ZnO and 3c-ZnO in Rats
Radiolabeling of 3a-ZnO, 3b-ZnO and 3c-ZnO for Imaging Studies
Imaging Studies of 1,2,3-Triazole-sulphadiazine-ZnO Hybrids in Uninfected Rats
Pneumonia Model
- (a)
- Bacterial Load Assessment
- (b)
- Histopathological Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wiegand, S.B.; Traeger, L.; Nguyen, H.K.; Rouillard, K.R.; Fischbach, A.; Zadek, F.; Ichinose, F.; Schoenfisch, M.H.; Carroll, R.W.; Blochad, D.B.; et al. Antimicrobial effects of nitric oxide in murine models of Klebsiella pneumonia. Redox Biol. 2021, 39, 101826. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Kapley, A. Antibiotic resistance: Global health crisis and metagenomics. Biotechnol. Rep. 2021, 29, e00604. [Google Scholar] [CrossRef] [PubMed]
- Glover, R.E.; Petticrew, M.P.; Mays, N.B.; Thompson, C. How pharmaceutical and diagnostic stakeholders construct policy solutions to a public health ‘crisis’: An analysis of submissions to a United Kingdom House of Commons inquiry into antimicrobial resistance. Crit. Public Health 2022, 1–10. [Google Scholar] [CrossRef]
- Ana, K.M.S.; Madriaga, J.; Espino, M.P. β-Lactam antibiotics and antibiotic resistance in Asian lakes and rivers: An overview of contamination, sources and detection methods. Environ. Pollut. 2021, 275, 116624. [Google Scholar] [CrossRef]
- Shaaban, M.M.; Ragab, H.M.; Akaji, K.; McGeary, R.P.; Bekhit, A.E.A.; Hussein, W.M.; Kurz, J.L.; Elwakil, B.H.; Bekhit, S.A.; Ibrahim, T.M.; et al. Design, synthesis, biological evaluation and in silico studies of certain aryl sulfonyl hydrazones conjugated with 1, 3-diaryl pyrazoles as potent metallo-β-lactamase inhibitors. Bioorg. Chem. 2020, 105, 104386. [Google Scholar] [CrossRef]
- Mojica, M.F.; Rossi, M.A.; Vila, A.J.; Bonomo, R.A. The urgent need for metallo-β-lactamase inhibitors: An unattended global threat. Lancet Infect. Dis. 2021, 22, e28–e34. [Google Scholar] [CrossRef]
- Salahuddin, P.; Kumar, A.; Khan, A.U. Structure, function of serine and metallo-β-lactamases and their inhibitors. Curr. Protein Pept. Sci. 2018, 19, 130–144. [Google Scholar] [CrossRef]
- Feng, H.; Liu, X.; Wang, S.; Fleming, J.; Wang, D.C.; Liu, W. The mechanism of NDM-1-catalyzed carbapenem hydrolysis is distinct from that of penicillin or cephalosporin hydrolysis. Nat. Commun. 2017, 8, 2242. [Google Scholar] [CrossRef]
- Kaushik, A.; Kaushik, M.; Lather, V.; Dua, J.S. Recent review on subclass B1 metallo-β-lactamases inhibitors: Sword for antimicrobial resistance. Curr. Drug Targets 2019, 20, 756–762. [Google Scholar] [CrossRef]
- Wang, T.; Xu, K.; Zhao, L.; Tong, R.; Xiong, L.; Shi, J. Recent research and development of NDM-1 inhibitors. Eur. J. Med. Chem. 2021, 223, 113667. [Google Scholar] [CrossRef] [PubMed]
- Masters, P.A.; O’Brian, T.A.; Zurlo, J.; Miller, D.Q.; Joshi, N. Trimethoprim-sulfamethoxazole revisited. Arch. Intern. Med. 2003, 163, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Hideo, K.; Hagihara, M.; Asai, N.; Umemura, T.; Shibata, Y.; Hirai, J.; Yamagishi, Y.; Iwamoto, T.; Mikamo, H. Efficacy of Trimethoprim–Sulfamethoxazole in Combination with an Echinocandin as a First-Line Treatment Option for Pneumocystis Pneumonia: A Systematic Review and Meta-Analysis. Antibiotics 2022, 11, 719. [Google Scholar]
- Haseeb, A.; Abourehab, M.A.S.; Almalki, W.A.; Almontashri, A.M.; Bajawi, S.A.; Aljoaid, A.M.; Alsahabi, B.M.; Algethamy, M.; AlQarni, A.; Iqbal, M.S.; et al. Trimethoprim-Sulfamethoxazole (Bactrim) Dose Optimization in Pneumocystis jirovecii Pneumonia (PCP) Management: A Systematic Review. Int. J. Environ. Res. Public Health 2022, 19, 2833. [Google Scholar] [CrossRef]
- Kalarikkal, N.; Mohan, S.; Oluwafemi, S.O.; Thomas, S. Characterization of Nanomaterials: Advances and Key Technologies; Woodhead Publishing: Sawston, UK, 2018. [Google Scholar]
- Miller, K.P.; Wang, L.; Benicewicz, B.C.; Decho, A.W. Inorganic nanoparticles engineered to attack bacteria. Chem. Soc. Rev. 2015, 44, 7787–7807. [Google Scholar] [CrossRef]
- Sawai, J. Quantitative evaluation of antibacterial activities of metallic oxide powders (ZnO, MgO and CaO) by conductimetric assay. J. Microbiol. Methods 2003, 54, 177–182. [Google Scholar] [CrossRef]
- Rivière, G.; Oueslati, S.; Gayral, M.; Créchet, J.-B.; Nhiri, N.; Jacquet, E.; Cintrat, J.-C.; Giraud, F.; van Heijenoort, C.; Lescop, E. NMR Characterization of the Influence of Zinc (II) Ions on the Structural and Dynamic Behavior of the New Delhi Metallo-β-Lactamase-1 and on the Binding with Flavonols as Inhibitors. ACS Omega 2020, 5, 10466–10480. [Google Scholar] [CrossRef]
- Edwards, R.; Hashmi, P.S.; Greenwood, D. The effect of zinc ions on the activity of metallo-beta-lactamases of Bacteroides fragilis. J. Med. Microbiol. 1997, 46, 807–809. [Google Scholar] [CrossRef] [Green Version]
- De Lucas-Gil, E.; Del Campo, A.; Pascual, L.; Monte-Serrano, M.; Menéndez, J.; Fernández, J.F.; Rubio-Marcos, F. The fight against multidrug-resistant organisms: The role of ZnO crystalline defects. Mater. Sci. Eng. C 2019, 99, 575–581. [Google Scholar] [CrossRef]
- Ali, S.G.; Ansari, M.A.; Alzohairy, M.A.; Alomary, M.N.; Jalal, M.; AlYahya, S.; Asiri, S.M.M.; Khan, H.M. Effect of biosynthesized ZnO nanoparticles on multi-drug resistant Pseudomonas aeruginosa. Antibiotics 2020, 9, 260. [Google Scholar] [CrossRef]
- De Lucas-Gil, E.; Reinosa, J.J.; Neuhaus, K.; Vera-Londono, L.; Martín-González, M.; Fernaández, J.F.; Rubio-Marcos, F. Exploring new mechanisms for effective antimicrobial materials: Electric contact-killing based on multiple schottky barriers. ACS Appl. Mater. Interfaces 2017, 9, 26219–26225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, O. Influence of particle size on the antibacterial activity of zinc oxide. Int. J. Inorg. Mater. 2001, 3, 643–646. [Google Scholar] [CrossRef]
- Padmavathy, N.; Vijayaraghavan, R. Enhanced bioactivity of ZnO nanoparticles—An antimicrobial study. Sci. Technol. Adv. Mater. 2008, 9, 035004. [Google Scholar] [CrossRef] [PubMed]
- Aljohani, F.S.; Rezki, N.; Aouad, M.R.; Elwakil, B.H.; Hagar, M.; Sheta, E.; Mogahed, N.M.F.H.; Bardaweel, S.K.; Hagras, N.A.-e. Synthesis, Characterization and Nanoformulation of Novel Sulfonamide-1, 2, 3-triazole Molecular Conjugates as Potent Antiparasitic Agents. Int. J. Mol. Sci. 2022, 23, 4241. [Google Scholar] [CrossRef] [PubMed]
- Yusof, Y.; Tan, D.T.C.; Arjomandi, O.K.; Schenk, G.; McGeary, R.P. Captopril analogues as metallo-beta-lactamase inhibitors. Bioorg. Med. Chem. Lett. 2016, 26, 1589–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Saez, I.; Mercuri, P.S.; Papamicael, C.; Kahn, R.; Frère, J.M.; Galleni, M.; Rossolini, G.M.; Dideberg, O. FEZ-1 metallo-beta-lactamase from Legionella gormanii modelled with D-captopril. J. Mol. Biol. 2003, 325, 651–660. [Google Scholar] [PubMed]
- Liénard, B.M.R.; Garau, G.; Horsfall, L.; Karsisiotis, A.I.; Damblon, C.; Lassaux, P.; Papamicael, C.; Roberts, G.C.K.; Galleni, M.; Dideberg, O.; et al. Structural basis for the broad-spectrum inhibition of metallo-β-lactamases by thiols. Org. Biomol. Chem. 2008, 6, 2282. [Google Scholar] [CrossRef] [Green Version]
- Sirelkhatim, A.; Mahmud, S.; Seeni, A.; Kaus, N.H.M.; Ann, L.C.; Bakhori, S.K.; Hasan, H.; Mohamad, D. Review on zinc oxide nanoparticles: Antibacterial activity and toxicity mechanism. Nano-Micro Lett. 2015, 7, 219–242. [Google Scholar] [CrossRef] [Green Version]
- Pasquet, J.; Chevalier, Y.; Pelletier, J.; Couval, E.; Bouvier, D.; Bolzinger, M.A. The contribution of zinc ions to the antimicrobial activity of zinc oxide. Colloids Surf. A Physicochem. Eng. Asp. 2014, 457, 263–274. [Google Scholar] [CrossRef]
- Xia, T.; Kovochich, M.; Liong, M.; Madler, L.; Gilbert, B.; Shi, H.; Yeh, J.I.; Zink, J.I.; Nel, A.E. Comparison of the mechanism of toxicity of zinc oxide and cerium oxide nanoparticles based on dissolution and oxidative stress properties. ACS Nano 2008, 2, 2121–2134. [Google Scholar] [CrossRef] [Green Version]
- Brem, J.; van Berkel, S.S.; Zollman, D.; Lee, S.Y.; Gileadi, O.; McHugh, P.J.; Walsh, T.R.; McDonough, M.A.; Schofield, C.J. Structural Basis of Metallo-β-Lactamase Inhibition by Captopril Stereoisomers. Antimicrob. Agents Chemother. 2016, 60, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, D.T.; Worrall, L.J.; Gruninger, R.; Strynadka, N.C.J. New Delhi metallo-beta-lactamase: Structural insights into beta-lactam recognition and inhibition. J. Am. Chem. Soc. 2012, 134, 11362–11365. [Google Scholar] [CrossRef]
- Krajnc, A.; Brem, J.; Hinchliffe, P.; Calvopiña, K.; Panduwawala, T.D.; Lang, P.A.; Kamps, J.J.A.G.; Tyrrell, J.M.; Widlake, E.; Saward, B.G.; et al. Bicyclic boronate VNRX-5133 inhibits metallo-and serine-β-lactamases. J. Med. Chem. 2019, 62, 8544–8556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.K.; Demuth, M.; Schlesinger, S.R.; Kim, S.J.; Urbanczyk, J.; Shaw, R.W.; Shin, H. Inhibition of Bacillus anthracis metallo-β-lactamase by compounds with hydroxamic acid functionality. J. Enzym. Inhib. Med. Chem. 2016, 31 (Suppl. S4), 132–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, D.T.; Strynadka, N.C. Targeting metallo-β-lactamase enzymes in antibiotic resistance. Future Med. Chem. 2013, 5, 1243–1263. [Google Scholar] [CrossRef] [PubMed]
- Elnaggar, Y.S.; Elwakil, B.H.; Elshewemi, S.S.; El-Naggar, M.Y.; Bekhit, A.A.; Olama, Z.A. Novel Siwa propolis and colistin-integrated chitosan nanoparticles: Elaboration; in vitro and in vivo appraisal. Nanomedicine 2020, 15, 1269–1284. [Google Scholar] [CrossRef]
- McAlinden, K.D.; Lu, W.; Ferdowsi, P.V.; Myers, S.; Markos, J.; Larby, J.; Chia, C.; Weber, H.C.; Haug, G.; Eapen, M.S.; et al. Electronic cigarette aerosol is cytotoxic and increases ACE2 expression on human airway epithelial cells: Implications for SARS-CoV-2 (COVID-19). J. Clin. Med. 2021, 10, 1028. [Google Scholar] [CrossRef]
- Danos, O.; Davies, K.; Lehn, P.; Mulligan, R. The ARRIVE guidelines, a welcome improvement to standards for reporting animal research. J. Gene Med. 2010, 12, 559–560. [Google Scholar] [CrossRef]
- Simone, E.A.; Zern, B.J.; Chacko, A.M.; Mikitsh, J.L.; Blankemeyer, E.R.; Muro, S.; Stan, R.V.; Muzykantov, V.R. Endothelial targeting of polymeric nanoparticles stably labeled with the PET imaging radioisotope iodine-124. Biomaterials 2012, 33, 5406–5413. [Google Scholar] [CrossRef] [Green Version]
- Lee-Lewis, H.; Anderson, D.M. Absence of inflammation and pneumonia during infection with nonpigmented Yersinia pestis reveals a new role for the pgm locus in pathogenesis. Infect. Immun. 2010, 78, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Li, Y.; Peng, C.; Lin, J.; Yu, F.; Zhao, Y.; Zhao, D. Outer membrane protein a in Acinetobacter baumannii induces autophagy through mTOR signaling pathways in the lung of SD rats. Biomed. Pharmacother. 2021, 135, 111034. [Google Scholar] [CrossRef] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IMP-1 Binding Score (Kcal/mol) | Hydrogen Bond Interactions | Hydrophobic Interactions |
|---|---|---|---|
| Captopril | −5.68 | Lys179 and Asn185 | His215 |
| 3a | −7.37 | Lys179 | Tyr181 and His215 |
| 3b | −7.45 | Lys179 and Gly184 | His215 |
| 3c | −7.15 | Lys179 | His215 |
| ZnO | −12.71 | - | - |
| Compound | NDM-1 Binding Score (Kcal/mol) | Hydrogen Bond Interactions | Hydrophobic Interactions |
|---|---|---|---|
| Captopril | −5.64 | Asn220 | His250 |
| 3a | −5.93 | Asn220 | His250 |
| 3b | −5.57 | Asn220 | His250 |
| 3c | −5.53 | - | His250 |
| ZnO | −13.11 | - | - |
| Bacterial Strain | NDM | IPM |
|---|---|---|
| K. pneumoniae ATCC 13883 | − | − |
| K. pneumoniae strain 1 | + | + |
| K. pneumoniae strain 2 | − | − |
| K. pneumoniae strain 3 | − | − |
| K. pneumoniae strain 4 | − | − |
| K. pneumoniae strain 5 | + | + |
| K. pneumoniae strain 6 | − | − |
| K. pneumoniae strain 7 | − | − |
| K. pneumoniae strain 8 | + | + |
| K. pneumoniae strain 9 | − | − |
| Tested Compounds | % Inhibition | NDM-1 | IPM-1 | |||
|---|---|---|---|---|---|---|
| NDM-1 (5 µM) | IPM-1 (5 µM) | Kic (µM) | Kiuc (µM) | Kic (µM) | Kiuc (µM) | |
| 3a | 71.2 | 75.0 | 9.6 | 6.0 | 11.8 | 7.7 |
| 3b | 68.7 | 75.4 | 9.0 | 6.0 | 11.4 | 7.6 |
| 3c | 64.3 | 72.9 | 8.5 | 3.9 | 10.9 | 5.1 |
| 3a-ZnO | 90.0 | 95.0 | 14.3 | 6.4 | 17.2 | 8.7 |
| 3b-ZnO | 87.0 | 95.5 | 14.7 | 6.5 | 17.8 | 9.4 |
| 3c-ZnO | 85.0 | 90.0 | 10.6 | 4.7 | 15.3 | 8.6 |
| Tested Compounds | Kp 1 | Kp 5 | Kp 8 | |||
|---|---|---|---|---|---|---|
| IZ (mm) | MIC (µg/mL) | IZ (mm) | MIC (µg/mL) | IZ (mm) | MIC (µg/mL) | |
| 3a | 9 | 128 | 7 | 256 | 8 | 128 |
| 3b | 10 | 128 | 7 | 256 | 10 | 128 |
| 3c | 6 | 256 | 6 | 256 | 6 | 256 |
| ZnO | 25 | 16 | 18 | 32 | 20 | 16 |
| 3a-ZnO | 37 | 4 | 30 | 8 | 32 | 8 |
| 3b-ZnO | 43 | 2 | 35 | 4 | 40 | 2 |
| 3c-ZnO | 33 | 8 | 25 | 16 | 28 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aljohani, F.S.; Rezki, N.; Aouad, M.R.; Hagar, M.; Bakr, B.A.; Shaaban, M.M.; Elwakil, B.H. Novel 1,2,3-Triazole-sulphadiazine-ZnO Hybrids as Potent Antimicrobial Agents against Carbapenem Resistant Bacteria. Antibiotics 2022, 11, 916. https://doi.org/10.3390/antibiotics11070916
Aljohani FS, Rezki N, Aouad MR, Hagar M, Bakr BA, Shaaban MM, Elwakil BH. Novel 1,2,3-Triazole-sulphadiazine-ZnO Hybrids as Potent Antimicrobial Agents against Carbapenem Resistant Bacteria. Antibiotics. 2022; 11(7):916. https://doi.org/10.3390/antibiotics11070916
Chicago/Turabian StyleAljohani, Faizah S., Nadjet Rezki, Mohamed R. Aouad, Mohamed Hagar, Basant A. Bakr, Marwa M. Shaaban, and Bassma H. Elwakil. 2022. "Novel 1,2,3-Triazole-sulphadiazine-ZnO Hybrids as Potent Antimicrobial Agents against Carbapenem Resistant Bacteria" Antibiotics 11, no. 7: 916. https://doi.org/10.3390/antibiotics11070916
APA StyleAljohani, F. S., Rezki, N., Aouad, M. R., Hagar, M., Bakr, B. A., Shaaban, M. M., & Elwakil, B. H. (2022). Novel 1,2,3-Triazole-sulphadiazine-ZnO Hybrids as Potent Antimicrobial Agents against Carbapenem Resistant Bacteria. Antibiotics, 11(7), 916. https://doi.org/10.3390/antibiotics11070916