
Potential Pharmacokinetic Effect of Chicken Xenobiotic Receptor Activator on Sulfadiazine: Involvement of P-glycoprotein Induction


Abstract
:1. Introduction
2. Results
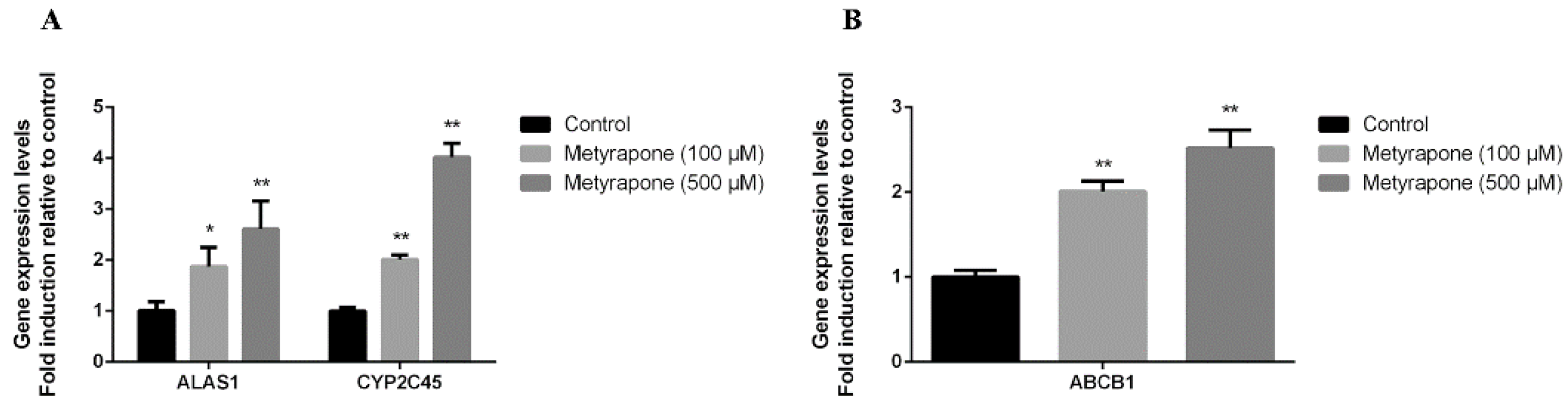
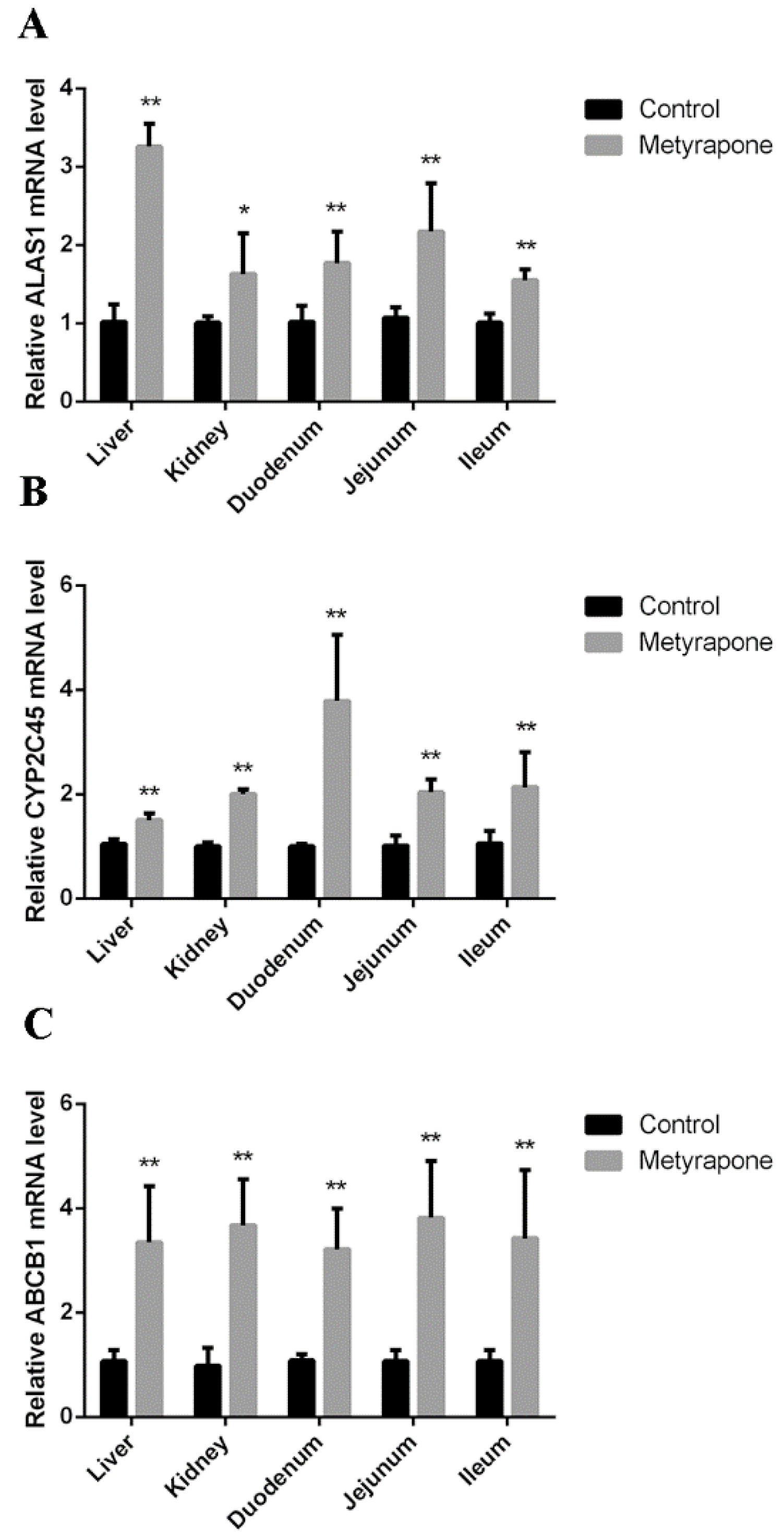
2.1. Metyrapone Upregulates Expression of ABCB1
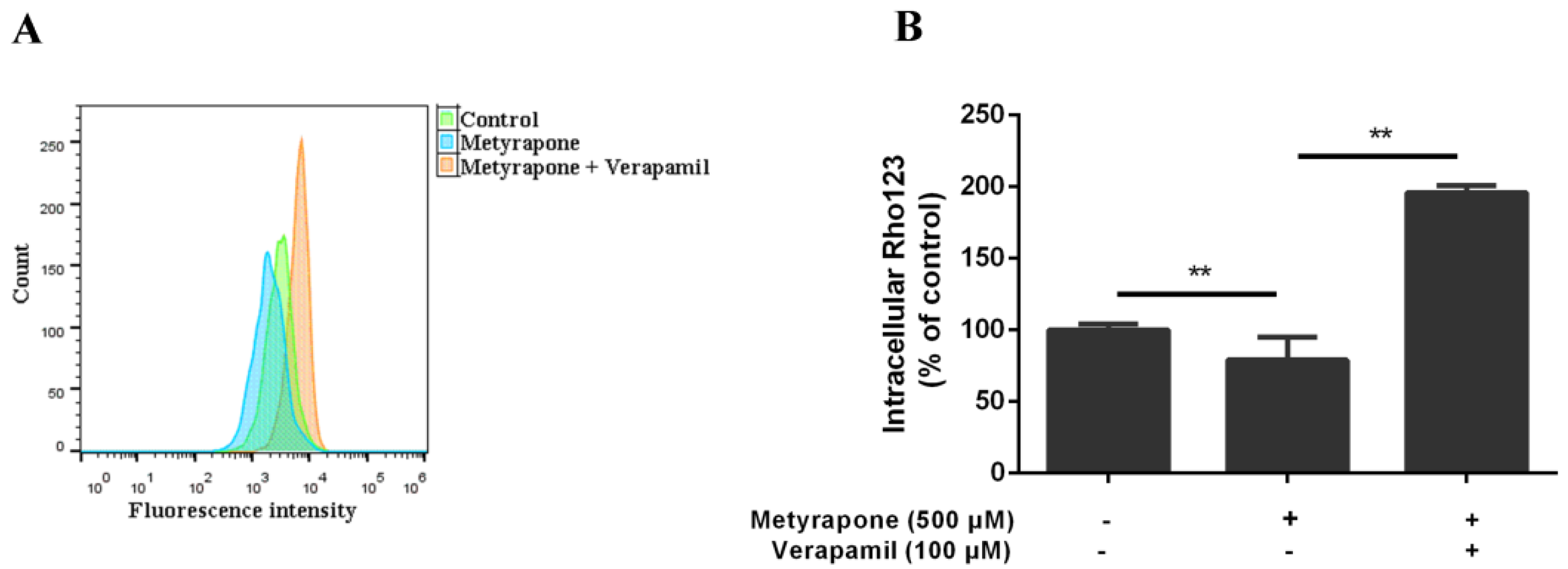
2.2. Agonist-Activated CXR Increases P-gp Function in Chicken Primary Hepatocytes
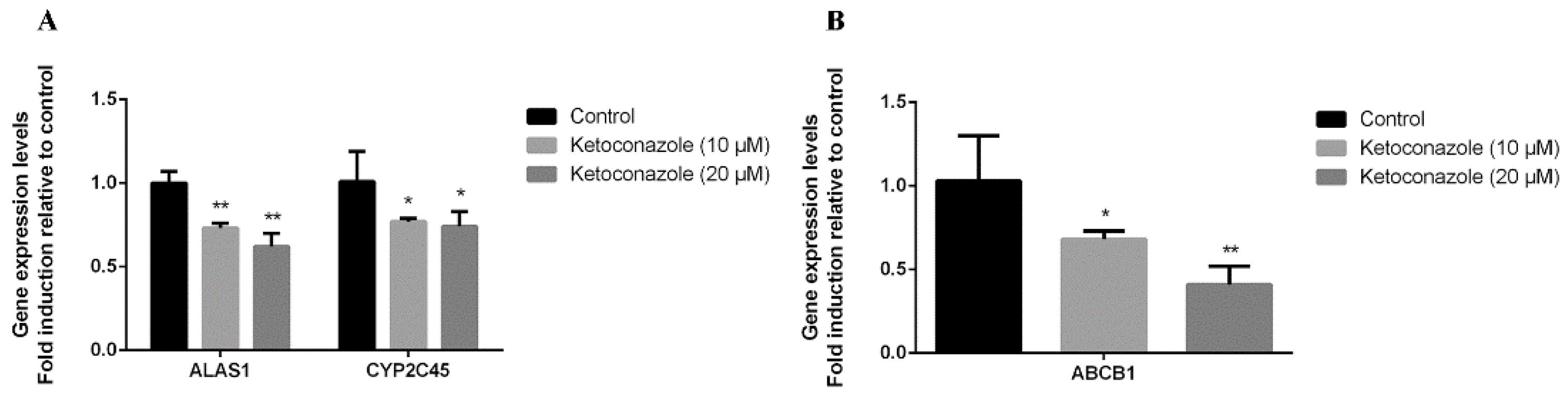
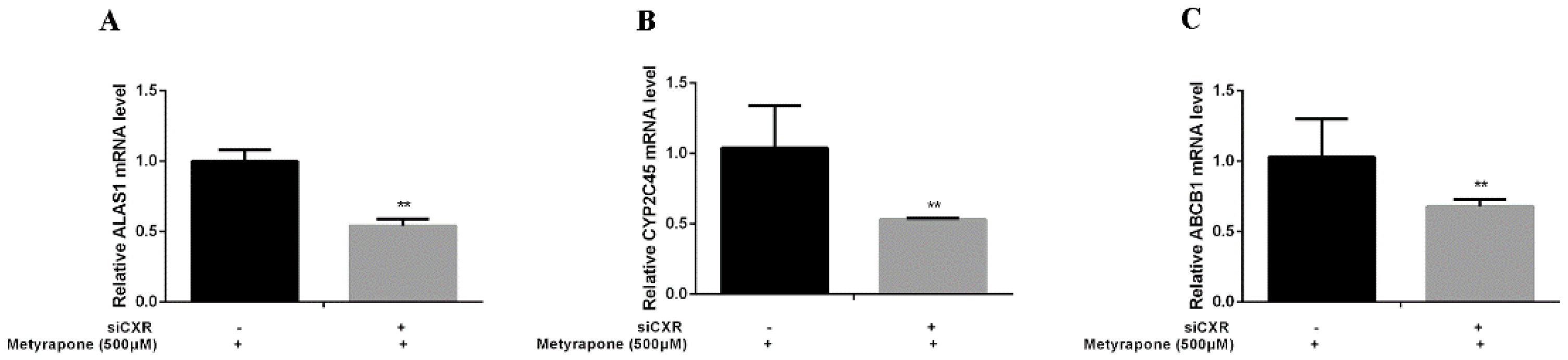
2.3. CXR Dependence of P-gp Induction
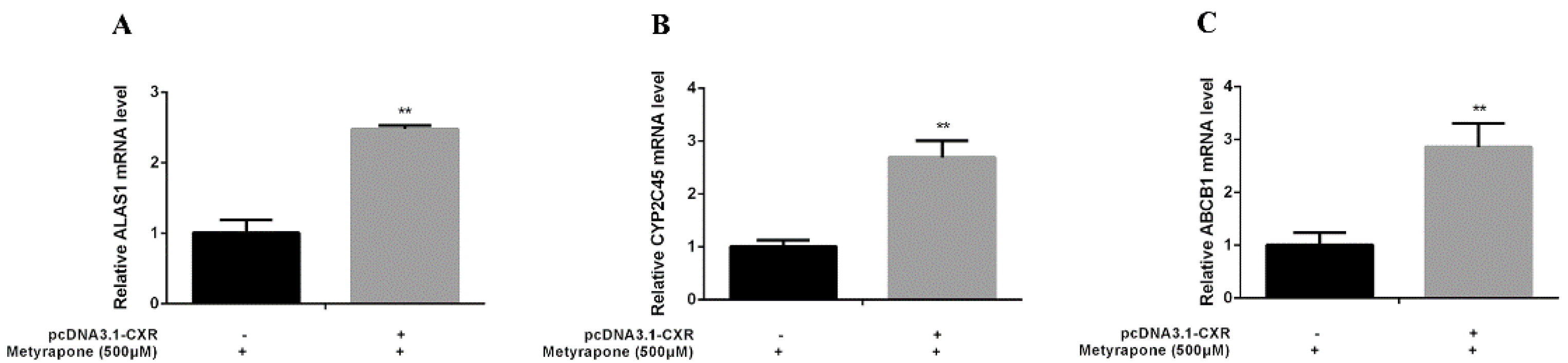
2.4. CXR-Mediated Induction of P-gp in Chicken
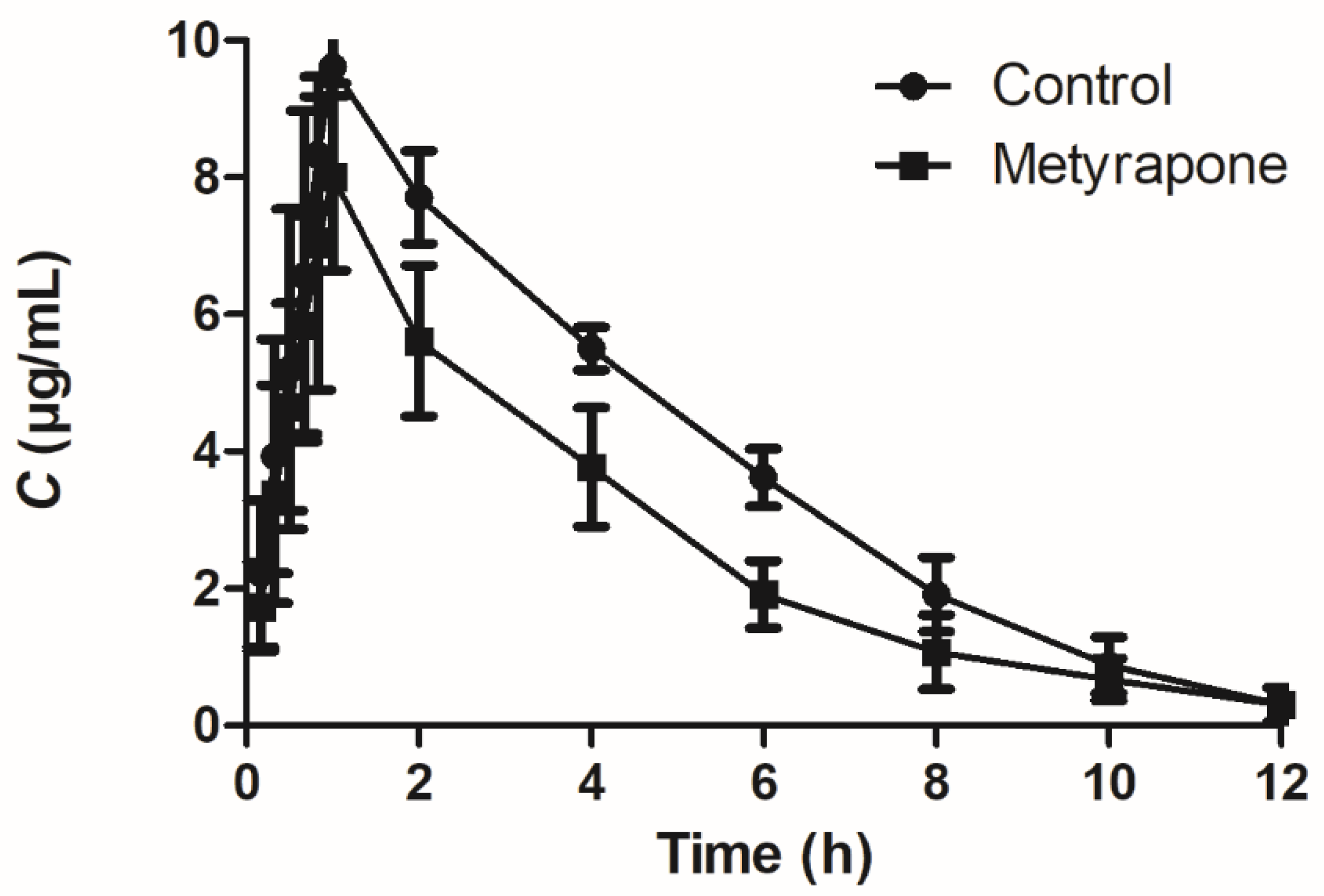
2.5. Agonist-Activated CXR Affects the Pharmacokinetics of the Orally Administered P-gp Substrate Sulfadiazine in Chickens
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Isolation of Chicken Livers and Preparation of Primary Hepatocyte Cultures
4.3. RNA Isolation and RT-PCR Analysis
4.4. Functional Detection of P-gp Activity
4.5. Overexpression of CXR
4.6. siRNA Studies
4.7. Experimental Animals and Sample Collection
4.8. Pharmacokinetic Studies of Sulfadiazine in Chicken
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, X. Abc Family Transporters. In Advances in Experimental Medicine and Biology; Springer: Singapore, 2019; Volume 1141, pp. 13–100. [Google Scholar]
- Mirzaei, S.; Gholami, M.H.; Hashemi, F.; Zabolian, A.; Farahani, M.V.; Hushmandi, K.; Zarrabi, A.; Goldman, A.; Ashrafizadeh, M.; Orive, G. Advances in Understanding the Role of P-Gp in Doxorubicin Resistance: Molecular Pathways, Therapeutic Strategies, and Prospects. Drug Discov. Today 2022, 27, 436–455. [Google Scholar] [CrossRef] [PubMed]
- Winter, H.R.; Unadkat, J.D. Identification of Cytochrome P450 and Arylamine N-Acetyltransferase Isoforms Involved in Sulfadiazine Metabolism. Drug Metab. Dispos. 2005, 33, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, J.; Liu, Y.; Guo, T.; Wang, L. Using the Lentiviral Vector System to Stably Express Chicken P-Gp and Bcrp in Mdck Cells for Screening the Substrates and Studying the Interplay of Both Transporters. Arch. Toxicol. 2018, 92, 2027–2042. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D. Transporter-Mediated Drug-Drug Interactions and Their Significance. In Drug Transporters in Drug Disposition, Effects and Toxicity; Springer: Singapore, 2019; Volume 1141, pp. 241–291. [Google Scholar]
- Kawahara, I.; Nishikawa, S.; Yamamoto, A.; Kono, Y.; Fujita, T. Assessment of Contribution of Bcrp to Intestinal Absorption of Various Drugs Using Portal-Systemic Blood Concentration Difference Model in Mice. Pharmacol. Res. Perspect. 2020, 8, e00544. [Google Scholar] [CrossRef]
- Wang, J.; Bwayi, M.; Gee, R.R.F.; Chen, T. Pxr-Mediated Idiosyncratic Drug-Induced Liver Injury: Mechanistic Insights and Targeting Approaches. Expert Opin. Drug Metab. Toxicol. 2020, 16, 711–722. [Google Scholar] [CrossRef]
- Daujat-Chavanieu, M.; Gerbal-Chaloin, S. Regulation of Car and Pxr Expression in Health and Disease. Cells 2020, 9, 2395. [Google Scholar] [CrossRef]
- Cai, X.; Young, G.M.; Xie, W. The Xenobiotic Receptors Pxr and Car in Liver Physiology, an Update. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166101. [Google Scholar] [CrossRef]
- Buchman, C.D.; Chai, S.C.; Chen, T. A Current Structural Perspective on Pxr and Car in Drug Metabolism. Expert Opin. Drug Metab. Toxicol. 2018, 14, 635–647. [Google Scholar] [CrossRef]
- McMillan, J.M.; Cobb, D.A.; Lin, Z.; Banoub, M.G.; Dagur, R.S.; Woods, A.A.B.; Wang, W.; Makarov, E.; Kocher, T.; Joshi, P.S.; et al. Antiretroviral Drug Metabolism in Humanized Pxr-Car-Cyp3a-Nog Mice. J. Pharmacol. Exp. Ther. 2018, 365, 272–280. [Google Scholar] [CrossRef]
- Wen, J.; Zhou, Y.; Li, Y. Pxr-Abc Drug Transporters/Cyp-Mediated Ursolic Acid Transport and Metabolism in Vitro and Vivo. Archiv. der Pharm. 2020, 353, 2000082. [Google Scholar]
- Elmeliegy, M.; Vourvahis, M.; Guo, C.; Wang, D.D. Effect of P-Glycoprotein (P-Gp) Inducers on Exposure of P-Gp Substrates: Review of Clinical Drug-Drug Interaction Studies. Clin. Pharmacokinet. 2020, 59, 699–714. [Google Scholar] [CrossRef]
- Ke, X.J.; Cheng, Y.F.; Yu, N.; Di, Q. Effects of Carbamazepine on the P-Gp and Cyp3a Expression Correlated with Pxr or Nf-Kappa B Activity in the Bend.3 Cells. Neurosci. Lett. 2019, 690, 48–55. [Google Scholar] [CrossRef]
- Handschin, C.; Podvinec, M.; Meyer, U.A. Cxr, a Chicken Xenobiotic-Sensing Orphan Nuclear Receptor, Is Related to Both Mammalian Pregnane X Receptor (Pxr) and Constitutive Androstane Receptor (Car). Proc. Natl. Acad. Sci. USA 2000, 97, 10769–10774. [Google Scholar] [CrossRef]
- Handschin, C.; Meyer, U.A. A Conserved Nuclear Receptor Consensus Sequence (Dr-4) Mediates Transcriptional Activation of the Chicken Cyp2h1 Gene by Phenobarbital in a Hepatoma Cell Line. J. Biol. Chem. 2000, 275, 13362–13369. [Google Scholar] [CrossRef]
- Baader, M.; Gnerre, C.; Stegeman, J.J.; Meyer, U.A. Transcriptional Activation of Cytochrome P450 Cyp2c45 by Drugs Is Mediated by the Chicken Xenobiotic Receptor (Cxr) Interacting with a Phenobarbital Response Enhancer Unit. J. Biol. Chem. 2002, 277, 15647–15653. [Google Scholar] [CrossRef]
- Fraser, D.J.; Podvinec, M.; Kaufmann, M.R.; Meyer, U.A. Drugs Mediate the Transcriptional Activation of the 5-Aminolevulinic Acid Synthase (AlAS1) Gene Via the Chicken Xenobiotic-Sensing Nuclear Receptor (Cxr). J. Biol. Chem. 2002, 277, 34717–34726. [Google Scholar] [CrossRef]
- Li, H.; Redinbo, M.R.; Venkatesh, M.; Ekins, S.; Chaudhry, A.; Bloch, N.; Negassa, A.; Mukherjee, P.; Kalpana, G.; Mani, S. Novel Yeast-Based Strategy Unveils Antagonist Binding Regions on the Nuclear Xenobiotic Receptor Pxr. J. Biol. Chem. 2013, 288, 13655–13668. [Google Scholar] [CrossRef]
- Das, B.C.; Madhukumar, A.V.; Anguiano, J.; Kim, S.; Sinz, M.; Zvyaga, T.A.; Power, E.C.; Ganellin, C.R.; Mani, S. Synthesis of Novel Ketoconazole Derivatives as Inhibitors of the Human Pregnane X Receptor (Pxr; Nr1i2; Also Termed Sxr, Par). Bioorg. Med. Chem. Lett. 2008, 18, 3974–3977. [Google Scholar] [CrossRef]
- Semeniuk, M.; Ceré, L.I.; Ciriaci, N.; Bucci-Muñoz, M.; Villanueva, S.S.M.; Mottino, A.D.; Catania, V.A.; Rigalli, J.P.; Ruiz, M.L. Regulation of Hepatic P-Gp Expression and Activity by Genistein in Rats. Arch. Toxicol. 2020, 94, 1625–1635. [Google Scholar] [CrossRef]
- Akamine, Y.; Yasui-Furukori, N.; Uno, T. Drug-Drug Interactions of P-Gp Substrates Unrelated to Cyp Metabolism. Curr. Drug Metab. 2019, 20, 124–129. [Google Scholar] [CrossRef]
- Zhang, L.; Ye, B.; Lin, Y.; Li, Y.; Wang, J.Q.; Chen, Z.; Ping, F.F.; Chen, Z.S. Ribociclib Inhibits P-Gp-Mediated Multidrug Resistance in Human Epidermoid Carcinoma Cells. Front. Pharmacol. 2022, 13, 867128. [Google Scholar] [CrossRef]
- Saib, S.; Hodin, S.; Bin, V.; Ollier, E.; Delavenne, X. In Vitro Evaluation of P-Gp-Mediated Drug-Drug Interactions Using the Rptec/Tert1 Human Renal Cell Model. Eur. J. Drug Metab. Pharmacokinet. 2022, 47, 223–233. [Google Scholar] [CrossRef]
- Bhutto, Z.A.; He, F.; Zloh, M.; Yang, J.; Huang, J.; Guo, T.; Wang, L. Use of Quercetin in Animal Feed: Effects on the P-Gp Expression and Pharmacokinetics of Orally Administrated Enrofloxacin in Chicken. Sci. Rep. 2018, 8, 4400. [Google Scholar] [CrossRef]
- Guo, M.; Dai, X.; Hu, D.; Zhang, Y.; Sun, Y.; Ren, W.; Wang, L. Potential Pharmacokinetic Effect of Rifampicin on Enrofloxacin in Broilers: Roles of P-Glycoprotein and Bcrp Induction by Rifampicin. Poult. Sci. 2016, 95, 2129–2135. [Google Scholar] [CrossRef]
- Whyte-Allman, S.K.; Hoque, M.T.; Jenabian, M.A.; Routy, J.P.; Bendayan, R. Xenobiotic Nuclear Receptors Pregnane X Receptor and Constitutive Androstane Receptor Regulate Antiretroviral Drug Efflux Transporters at the Blood-Testis Barriers. J. Pharmacol. Exp. Ther. 2017, 363, 324–335. [Google Scholar] [CrossRef]
- Manda, V.K.; Avula, B.; Dale, O.R.; Ali, Z.; Khan, I.A.; Walker, L.A.; Khan, S.I. Pxr Mediated Induction of Cyp3a4, Cyp1a2, and P-Gp by Mitragyna Speciosa and Its Alkaloids. Phytother. Res. 2017, 31, 1935–1945. [Google Scholar] [CrossRef]
- Lv, C.; Huang, L. Xenobiotic Receptors in Mediating the Effect of Sepsis on Drug Metabolism. Acta Pharm. Sin. B 2020, 10, 33–41. [Google Scholar] [CrossRef]
- Zhao, L.; Bin, S.; He, H.L.; Yang, J.M.; Pu, Y.C.; Gao, C.H.; Wang, H.; Wang, B.L. Sodium Butyrate Increases P-Gp Expression in Lung Cancer by Upregulation of Stat3 and Mrna Stabilization of Abcb1. Anti-Cancer Drugs 2018, 29, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, J.; Li, X.; Fang, C.; Wang, L. Identification of Functional Transcriptional Binding Sites within Chicken Abcg2 Gene Promoter and Screening Its Regulators. Genes 2020, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Hunt, T.L.; Tzanis, E.; Bai, S.; Manley, A.; Chitra, S.; McGovern, P.C. The Effect of Verapamil, a P-Gp Inhibitor, on the Pharmacokinetics, Safety, and Tolerability of Omadacycline in Healthy Adults: A Phase I, Open-Label, Single-Sequence Study. Eur. J. Drug Metab. Pharmacokinet. 2021, 46, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.S.; Nocera, A.; Workman, A.; Amiji, M.M.; Bleier, B.S. P-Glycoprotein Inhibition with Verapamil Overcomes Mometasone Resistance in Chronic Sinusitis with Nasal Polyps. Rhinology 2021, 59, 205–211. [Google Scholar] [CrossRef]
- Marier, J.F.; Deschênes, J.L.; Hage, A.; Seliniotakis, E.; Gritsas, A.; Flarakos, T.; Beaudry, F.; Vachon, P. Enhancing the Uptake of Dextromethorphan in the Cns of Rats by Concomitant Administration of the P-Gp Inhibitor Verapamil. Life Sci. 2005, 77, 2911–2926. [Google Scholar] [CrossRef]
- Merelli, A.; Ramos, A.J.; Lazarowski, A.; Auzmendi, J. Convulsive Stress Mimics Brain Hypoxia and Promotes the P-Glycoprotein (P-Gp) and Erythropoietin Receptor Overexpression. Recombinant Human Erythropoietin Effect on P-Gp Activity. Front. Neurosci. 2019, 13, 750. [Google Scholar] [CrossRef]
- Ogushi, N.; Sasaki, K.; Shimoda, M. Can a P-Gp Modulator Assist in the Control of Methotrexate Concentrations in the Rat Brain? -Inhibitory Effects of Rhodamine 123, a Specific Substrate for P-Gp, on Methotrexate Excretion from the Rat Brain and Its Optimal Route of Administration. J. Vet. Med. Sci. 2017, 79, 320–327. [Google Scholar] [CrossRef]
- Yano, K.; Seto, S.; Kamioka, H.; Mizoi, K.; Ogihara, T. Testosterone and Androstenedione Are Endogenous Substrates of P-Glycoprotein. Biochem. Biophys. Res. Commun. 2019, 520, 166–170. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Sulfadiazine | Sulfadiazine + Metyrapone | p-Value |
|---|---|---|---|
| Cmax (μg/mL) | 9.61 ± 0.41 | 8.01 ± 1.37 * | 0.03 |
| Tmax (h) | 0.94 ± 0.14 | 1.11 ± 0.44 | 0.40 |
| T1/2λ (h) | 1.67 ± 0.38 | 2.42 ± 0.67 * | 0.04 |
| Vz/F (L/kg) | 1.03 ± 0.19 | 2.16 ± 0.56 ** | 0.00 |
| Cl/F (L/h/kg) | 0.43 ± 0.02 | 0.62 ± 0.10 ** | 0.00 |
| AUC0-t (h·mg/L) | 45.59 ± 1.87 | 31.46 ± 4.46 ** | 0.00 |
| Name | Primer Sequence (5′–3′) |
|---|---|
| Primers for plasmid construction | |
| pcDNA3.1-CXR-F | CCCAAGCTTATGTCCCAGTCCAGCCCCTCGGAC |
| pcDNA3.1-CXR-R | CTAGTCTAGATCAGCTGATGATTTCGGAGAGCAGCGGAGTCATGC |
| Primers for RNA interference | |
| siCXR-F | UCAGGCGCUCCAUCCUUAATT |
| siCXR-R | UUAAGGAUGGAGCGCCUGATT |
| NC-F | UUCUCCGAACGUGUCACGUTT |
| NC-R | ACGUGACACGUUCGGAGAATT |
| Primers for RT-PCR | |
| ABCB1 (P-gp)-F | ACCAGTCTCCCTATAGCAATG |
| ABCB1 (P-gp)-R | GGATATAAGCAGCCACAAGAAC |
| ALAS1-F | CATCTCTGGAACGCTCGGCAAG |
| ALAS1-R | CCAGCAGCATACGAACGGACAG |
| CYP2C45-F | CGATACGGGCTTCTGCTTCTTCTC |
| CYP2C45-R | TGGACCACTGCGTCTGTGTAGG |
| β-actin-F | TCCCTTCGGCATCCCTGTC |
| β-actin-R | GGCGTTGGTCTCCTCGTTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Xu, Z.; Lu, W.; Wang, L.; Zhang, Y. Potential Pharmacokinetic Effect of Chicken Xenobiotic Receptor Activator on Sulfadiazine: Involvement of P-glycoprotein Induction. Antibiotics 2022, 11, 1005. https://doi.org/10.3390/antibiotics11081005
Li M, Xu Z, Lu W, Wang L, Zhang Y. Potential Pharmacokinetic Effect of Chicken Xenobiotic Receptor Activator on Sulfadiazine: Involvement of P-glycoprotein Induction. Antibiotics. 2022; 11(8):1005. https://doi.org/10.3390/antibiotics11081005
Chicago/Turabian StyleLi, Mei, Ziyong Xu, Wang Lu, Liping Wang, and Yujuan Zhang. 2022. "Potential Pharmacokinetic Effect of Chicken Xenobiotic Receptor Activator on Sulfadiazine: Involvement of P-glycoprotein Induction" Antibiotics 11, no. 8: 1005. https://doi.org/10.3390/antibiotics11081005
APA StyleLi, M., Xu, Z., Lu, W., Wang, L., & Zhang, Y. (2022). Potential Pharmacokinetic Effect of Chicken Xenobiotic Receptor Activator on Sulfadiazine: Involvement of P-glycoprotein Induction. Antibiotics, 11(8), 1005. https://doi.org/10.3390/antibiotics11081005