
Prevalence and Antimicrobial Resistance of Virulent Listeria monocytogenes and Cronobacter sakazakii in Dairy Cattle, the Environment, and Dried Milk with the In Vitro Application of Natural Alternative Control

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Prevalence of L. monocytogenes and C. sakazakii in Cattle, the Environment, and Dried Milk
2.2. Antibiogram of Isolated Strains of L. monocytogenes and C. sakazakii
2.3. Prevalence and Distribution of L. monocytogenes Virulence Genes with Their Antimicrobial Resistance Profile and Multiple Antibiotic Resistance (MAR) Index
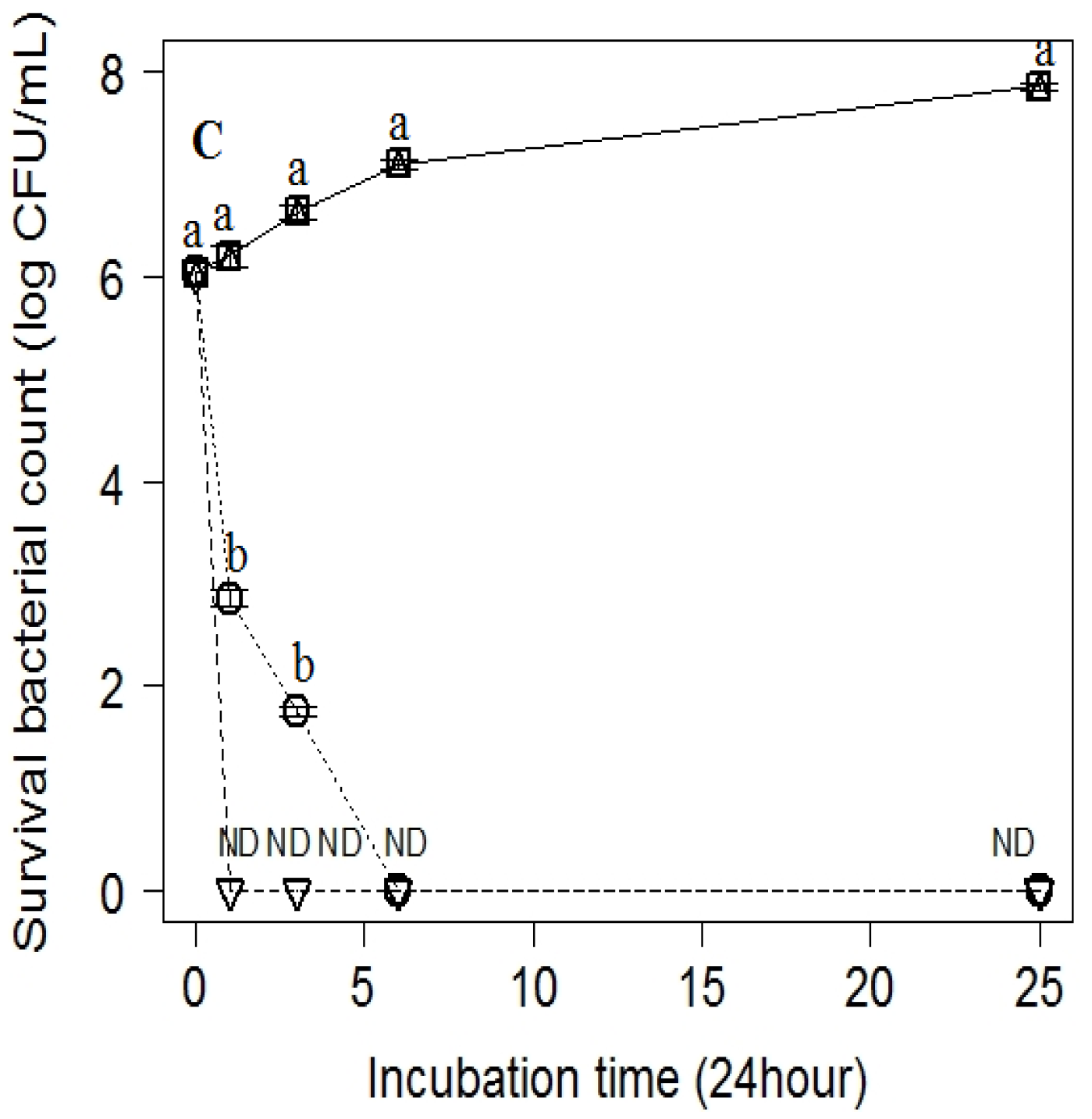
2.4. Antibacterial Activity Assessment of the Selected Natural Products against Standard Bacterial Strain
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Bacterial Isolation and Identification
4.3. DNA Extraction
4.4. Antibiotic Resistance of L. monocytogenes and C. sakazakii Isolated from Dairy Cattle and the Environment
4.5. In Vitro Trials for Microbial Reduction in L. monocytogenes and C. sakazakii with Rose Water, Rose, and Orange EOs
4.5.1. Preparation of Bacterial Suspension
4.5.2. Preparation of Plant Extract Products
4.5.3. Determination of Minimal Inhibitory Concentration (MIC) and Minimal Microbicidal Concentration (MMC)
4.6. Time-Kill Assay
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tahoun, A.B.M.B.; Abou Elez, R.M.M.; Abdelfatah, E.N.; Elsohaby, I.; El-Gedawy, A.A.; Elmoslemany, A.M. Listeria monocytogenes in raw milk, milking equipment and dairy workers: Molecular characterization and antimicrobial resistance patterns. J. Glob. Antimicrob. Resist. 2017, 10, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, M.M.; Elkenany, R.M.; Zakaria, A.I.; Badawy, B.M. Epidemiological study on Listeria monocytogenes in Egyptian dairy cattle farms’ insights into genetic diversity of multi-antibiotic-resistant strains by ERIC-PCR. Environ. Sci. Pollut. Res. 2022, 29, 54359–54377. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, C.; Braun, P.; Hammer, P. Reservoir and routes of transmission of Enterobacter sakazakii (Cronobacter spp.) in a milk powder-producing plant. J. Dairy Sci. 2011, 94, 3801–3810. [Google Scholar] [CrossRef]
- Bandelj, P.; Jamnikar-Ciglenecki, U.; Ocepek, M.; Blagus, R.; Vengust, M. Risk factors associated with fecal shedding of Listeria monocytogenes by dairy cows and calves. J. Vet. Intern. Med. 2018, 32, 1773–1779. [Google Scholar] [CrossRef] [PubMed]
- Iversen, C.; Druggan, P.; Schumacher, S.; Lehner, A.; Feer, C.; Gschwend, K.; Joosten, H.; Stephan, R. Development of a novel screening method for the isolation of “Cronobacter” spp. (Enterobacter sakazakii). Appl. Environ. Microbiol. 2008, 74, 2550–2553. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Ge, W.; Li, K.; Gan, J.; Zhang, Y.; Zhang, Q.; Luo, R.; Chen, L.; Liang, Y.; Wang, Q.; et al. Prevalence and Characterization of Cronobacter sakazakii in Retail Milk-Based Infant and Baby Foods in Shaanxi, China. Foodborne Pathog. Dis. 2016, 13, 221–227. [Google Scholar] [CrossRef]
- Brandão, M.L.L.; Umeda, N.S.; Jackson, E.; Forsythe, S.J.; de Filippis, I. Isolation, molecular and phenotypic characterization, and antibiotic susceptibility of Cronobacter spp. from Brazilian retail foods. Food Microbiol. 2017, 63, 129–138. [Google Scholar] [CrossRef]
- Awadallah, M.A.I.; Ahmed, H.A.; Merwad, A.M.A.; Abou Elez, R.M.M.; Saleh, K.M.A. Molecular Characterization of Cronobacter sakazakii in Egypt, Survival and Thermoresistance at Different Temperatures: A Potential Public Health Risk. Vector-Borne Zoonotic Dis. 2018, 18, 101–107. [Google Scholar] [CrossRef]
- Jamali, H.; Radmehr, B.; Thong, K.L. Prevalence, characterisation, and antimicrobial resistance of Listeria species and Listeria monocytogenes isolates from raw milk in farm bulk tanks. Food Control 2013, 34, 121–125. [Google Scholar] [CrossRef]
- Dahshan, H.; Abd-Elall, A.M.M.; Megahed, A.M.; Abd-El-Kader, M.A.; Nabawy, E.E. Veterinary antibiotic resistance, residues, and ecological risks in environmental samples obtained from poultry farms, Egypt. Environ. Monit. Assess. 2015, 187, 2. [Google Scholar] [CrossRef]
- World Health Organization. Report on the consultative meeting on antimicrobial resistance for countries in the Eastern Mediterranean Region:from policies to action Sharm el Sheikh, Egypt, 12–14 November 2013. In Consultative Meeting on Antimicrobial Resistance for Countries in the Eastern Mediterranean Region: From Policies to Action Sharm ElSheikh; Egypt, November 2013; Available online: http://applications.emro.who.i (accessed on 12 July 2022).
- Swetha, C.S.; Porteen, K.; Elango, A.; Ronald, B.S.M.; Senthil Kumar, T.M.A.; Milton, A.P.; Sureshkannan, S. Genetic diversity, virulence and distribution of antimicrobial resistance among Listeria monocytogenes isolated from milk, beef, and bovine farm environment. Iran. J. Vet. Res. 2021, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Aziza, A.E.; Abdelhamid, F.M.; Risha, E.F.; Elsayed, M.M.; Awadin, W.F. Influence of Nigella sativa and rosemary oils on growth performance, biochemical, antioxidant and immunological parameters, and pathological changes in Japanese quail challenged with Escherichia coli. J. Anim. Feed. Sci. 2019, 28, 354–366. [Google Scholar] [CrossRef]
- da Silva, C.F.; de Oliveira, F.S.M.; Caetano, V.F.; Vinhas, G.M.; Cardoso, S.A. Orange essential oil as antimicrobial additives in poly(vinyl chloride) films. Polimeros 2018, 5169, 332–338. [Google Scholar] [CrossRef]
- Ulusoy, S.; Boşgelmez-TInaz, G.; Seçilmiş-Canbay, H. Tocopherol, carotene, phenolic contents and antibacterial properties of rose essential oil, hydrosol and absolute. Curr. Microbiol. 2009, 59, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, A.A.; Al-Askar, A.A.; Almaary, K.S.; Dawoud, T.M.; Sholkamy, E.N.; Bakri, M.M. Antimicrobial activity of some plant extracts against bacterial strains causing food poisoning diseases. Saudi J. Biol. Sci. 2018, 25, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Hamoud, R.; Sporer, F.; Reichling, J.; Wink, M. Antimicrobial activity of a traditionally used complex essential oil distillate (Olbas® Tropfen) in comparison to its individual essential oil ingredients. Phytomedicine 2012, 19, 969–976. [Google Scholar] [CrossRef]
- Kim, S.W.; Haendiges, J.; Keller, E.N.; Myers, R.; Kim, A.; Lombard, J.E.; Karns, J.S.; Van Kessel, J.A.S.; Haley, B.J. Genetic diversity and virulence profiles of Listeria monocytogenes recovered from bulk tank milk, milk filters, and milking equipment from dairies in the United States (2002 to 2014). PLoS ONE 2018, 13, 1–17. [Google Scholar] [CrossRef]
- Mohammed, A.N.; Abdel Aziz, S.A.A. Ecological Study on Listeria monocytogenes and the Extent of its Resistance to Different Disinfectants in Dairy Farm for Improving Animal Health. Asian J. Anim. Vet. Adv. 2017, 12, 302–310. [Google Scholar] [CrossRef] [Green Version]
- Esteban, J.I.; Oporto, B.; Aduriz, G.; Juste, R.A.; Hurtado, A. Faecal shedding and strain diversity of Listeria monocytogenes in healthy ruminants and swine in Northern Spain. BMC Vet. Res. 2009, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Vilar, M.J.; Yus, E.; Sanjuán, M.L.; Diéguez, F.J.; Rodríguez-Otero, J.L. Prevalence of and risk factors for Listeria species on dairy farms. J. Dairy Sci. 2007, 90, 5083–5088. [Google Scholar] [CrossRef]
- Castro, H.; Jaakkonen, A.; Hakkinen, M.; Korkeala, H.; Lindström, M. Occurrence, persistence, and contamination routes of Listeria monocytogenes genotypes on three Finnish dairy cattle farms: A longitudinal study. Appl. Environ. Microbiol. 2018, 84, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdeen, E.E.; Mousa, W.S.; Harb, O.H.; Fath-Elbab, G.A.; Nooruzzaman, M.; Gaber, A.; Alsanie, W.F.; Abdeen, A. Prevalence, antibiogram and genetic characterization of listeria monocytogenes from food products in Egypt. Foods 2021, 10, 1381. [Google Scholar] [CrossRef] [PubMed]
- Ismaiel, A.A.R.; Ali, A.E.S.; Enan, G. Incidence of Listeria in Egyptian meat and dairy samples. Food Sci. Biotechnol. 2014, 23, 179–185. [Google Scholar] [CrossRef]
- Mohammed, H.O.; Stipetic, K.; McDonough, P.L.; Gonzalez, R.N.; Nydam, D.V.; Atwill, E.R. Identification of potential on-farm sources of Listeria monocytogenes in herds of dairy cattle. J. Am. Vet. Med. Assoc. 2009, 234, 673. Available online: https://avmajournals.avma.org/view/journals/ajvr/70/3/ajvr.70.3.383.xml (accessed on 4 August 2022). [CrossRef]
- Rodas-Suzáre, O.R.; Quiñones-Ramírez, E.I.; Fernández, F.J.; Vázquez-Salinas, C. Listeria monocytogenes strains isolated from dry milk samples in Mexico: Occurrence and antibiotic sensitivity. J. Environ. Health 2013, 76, 32–37. [Google Scholar]
- Şanlibaba, P.; Uymaz Tezel, B.; Çakmak, G.A. Detection of Listeria Spp. in Raw Milk and Dairy Products Retailed in Ankara. Gida/J. Food 2018, 43, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Ogihara, H.; Fukuda, N.; Fujiwara, A.; Suzuki, H.; Yamamoto, H.; Kyoui, D.; Okada, Y.; Igimi, S. Prevalence of Cronobacter spp. In Retail Foods and Farm-associated Environments in Japan. Food Sci. Technol. Res. 2019, 25, 265–275. [Google Scholar] [CrossRef]
- Mozrová, V.; Břeňová, N.; Mrázek, J.; Lukešová, D.; Marounek, M. Surveillance and characterisation of Cronobacter spp. in Czech retail food and environmental samples. Folia Microbiol. 2014, 59, 63–68. [Google Scholar] [CrossRef]
- Molloy, C.; Cagney, C.; O’Brien, S.; Iversen, C.; Fanning, S.; Duffy, G. Surveillance and characterisation by Pulsed-Field Gel Electrophoresis of Cronobacter spp. in farming and domestic environments, food production animals and retail foods. Int. J. Food Microbiol. 2009, 136, 198–203. [Google Scholar] [CrossRef]
- El-Gamal, M.S.; El Dairouty, R.K.; Okda, A.Y.; Salah, S.H.; El-Shamy, S.M. Incidence and interrelation of Cronobacter sakazakii and other foodborne bacteria in some milk products and infant formula milks in Cairo and Giza area. World Appl. Sci. J. 2013, 26, 1129–1141. [Google Scholar] [CrossRef]
- Berthold-Pluta, A.; Garbowska, M.; Stefańska, I.; Stasiak-Różańska, L.; Aleksandrzak-Piekarczyk, T.; Pluta, A. Microbiological quality of nuts, dried and candied fruits, including the prevalence of cronobacter spp. Pathogens 2021, 10, 900. [Google Scholar] [CrossRef] [PubMed]
- AL-Ashmawy, M.A.; Gwida, M.; Abdelgalil, K.H. Prevalence, Detection Methods and Antimicrobial Susceptibility of Listeria monocytogens Isolated from Milk and Soft Cheeses and its Zoonotic Importance. World Appl. Sci. J. 2014, 29, 869–878. [Google Scholar] [CrossRef]
- Abiral Hasib Shourav, A. Hasib Shourav, Mahedy Hasan, M. Hasan.; Sangita Ahmed, S. Ahmed. Antibiotic susceptibility pattern of Listeria spp. isolated from cattle farm environment in Bangladesh. J. Agric. Food Res. 2020, 2, 100082. [Google Scholar] [CrossRef]
- Andriyanov, P.A.; Zhurilov, P.A.; Liskova, E.A.; Karpova, T.I.; Sokolova, E.V.; Yushina, Y.K.; Zaiko, E.V.; Bataeva, D.S.; Voronina, O.L.; Psareva, E.K.; et al. Antimicrobial resistance of listeria monocytogenes strains isolated from humans, animals, and food products in russia in 1950–1980, 2000–2005, and 2018–2021. Antibiotics 2021, 10, 1206. [Google Scholar] [CrossRef] [PubMed]
- Adesokan, H.K.; Kehinde, E.G. Prevalence, Antibiogram and Biofilm Production of Listeria Monocytogenes from Faeces and Foetuses of Slaughtered Pregnant Cows: Environmental and Public Health Implications. Folia Vet. 2021, 65, 17–26. [Google Scholar] [CrossRef]
- Fei, P.; Jiang, Y.; Feng, J.; Forsythe, S.J.; Li, R.; Zhou, Y.; Man, C. Antibiotic and desiccation resistance of Cronobacter sakazakii and C. malonaticus isolates from powdered infant formula and processing environments. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra-Flores, J.; Holý, O.; Riffo, F.; Lepuschitz, S.; Maury-Sintjago, E.; Rodríguez-Fernández, A.; Cruz-Córdova, A.; Xicohtencatl-Cortes, J.; Mancilla-Rojano, J.; Troncoso, M.; et al. Profiling the Virulence and Antibiotic Resistance Genes of Cronobacter sakazakii Strains Isolated From Powdered and Dairy Formulas by Whole-Genome Sequencing. Front. Microbiol. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- El-Sharoud, W.M.; O’Brien, S.; Negredo, C.; Iversen, C.; Fanning, S.; Healy, B. Characterization of Cronobacter recovered from dried milk and related products. BMC Microbiol. 2009, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Androutsopoulou, C.; Christopoulou, S.D.; Hahalis, P.; Kotsalou, C.; Lamari, F.N.; Vantarakis, A. Evaluation of essential oils and extracts of rose geranium and rose petals as natural preservatives in terms of toxicity, antimicrobial, and antiviral activity. Pathogens 2021, 10, 494. [Google Scholar] [CrossRef]
- Shohayeb, M.; Abdel-Hameed, E.S.S.; Bazaid, S.A.; Maghrabi, I. Antibacterial and antifungal activity of Rosa damascena MILL. essential oil, different extracts of rose petals. Glob. J. Pharmacol. 2014, 8, 1–7. [Google Scholar] [CrossRef]
- Settanni, L.; Palazzolo, E.; Guarrasi, V.; Aleo, A.; Mammina, C.; Moschetti, G.; Germanà, M.A. Inhibition of foodborne pathogen bacteria by essential oils extracted from citrus fruits cultivated in Sicily. Food Control. 2012, 26, 326–330. [Google Scholar] [CrossRef]
- Geraci, A.; Di Stefano, V.; Di Martino, E.; Schillaci, D.; Schicchi, R. Essential oil components of orange peels and antimicrobial activity. Nat. Prod. Res. 2017, 31, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Fraňková, A.; Marounek, M.; Mozrová, V.; Weber, J.; Klouček, P.; Lukešová, D. Antibacterial activities of plant-derived compounds and essential oils toward cronobacter sakazakii and cronobacter malonaticus. Foodborne Pathog. Dis. 2014, 11, 795–797. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Febres, V.J.; Jones, J.B.; Moore, G.A. A survey of FLS2 genes from multiple citrus species identifies candidates for enhancing disease resistance to Xanthomonas citri ssp. citri. Hortic. Res. 2016, 3, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzi Gulel, G.; Gucukoglu, A.; Cadirci, O.; Saka, E.; Alisarli, M. Serotyping and antibiotic resistance of Listeria monocytogenes isolated from raw water buffalo milk and milk products. J. Food Sci. 2020, 85, 2889–2895. [Google Scholar] [CrossRef]
- Matar, G.M.; Hayes, P.S.; Bibb, W.F.; Swaminathan, B. Listeriolysin O-based latex agglutination test for the rapid detection of Listeria monocytogenes in foods. J. Food Prot. 1997, 60, 1038–1040. [Google Scholar] [CrossRef]
- Kaur, S.; Malik, S.V.S.; Vaidya, V.M.; Barbuddhe, S.B. Listeria monocytogenes in spontaneous abortions in humans and its detection by multiplex PCR. J. Appl. Microbiol. 2007, 103, 1889–1896. [Google Scholar] [CrossRef]
- Carter, L.; Lindsey, L.A.; Grim, C.J.; Sathyamoorthy, V.; Jarvis, K.G.; Gopinath, G.; Lee, C.; Sadowski, J.A.; Trach, L.; Pava-Ripoll, M.; et al. Multiplex PCR assay targeting a diguanylate cyclase-encoding gene, cgcA, to differentiate species within the genus Cronobacter. Appl. Environ. Microbiol. 2013, 79, 734–737. [Google Scholar] [CrossRef] [Green Version]
- Hindler, J.A.; Matuschek, E.; Cullen, S.K.; Castanheira, M.; Giske, C.G.; Kahlmeter, G.; Koeth, L.M.; Traczewski, M.; Turnidge, J.D.; Wootton, M. Procedure for Optimizing Disk Contents (Potencies) for Disk Diffusion Testing of Antimicrobial Agents Using Harmonized CLSI and EUCAST Criteria; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020; ISBN 978-1-68440-079-9. [Google Scholar]
- Prabuseenivasan, S.; Jayakumar, M.; Ignacimuthu, S. In vitro antibacterial activity of some plant essential oils. BMC Complement. Altern. Med. 2006, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal House | Samples | Total Examined No | L. monocytogenes-Positive No. (%) | C. sakazakii-Positive No. (%) | p-Value |
| Household I | Animal samples | ||||
| Fecal samples | 40 | 9 (22.5) | 3 (7.5) | 0.06 | |
| Milk | 40 | 5 (12.5) | 0 (0) | 0.02 | |
| Total | 80 | 14 (17.5) | 3 (3.7) | 0.004 | |
| Environment | |||||
| Straw bedding | 40 | 7 (17.5) | 2 (5) | 0.076 | |
| Total | 120 | 21 (17.5) | 5 (4.1) | 0.001 | |
| Household II | Animal samples | ||||
| Fecal samples | 40 | 11 (27.5) | 0 (0) | ≤0.001 | |
| Milk | 40 | 4 (10) | 0 (0) | 0.04 | |
| Total | 80 | 15 (18.7) | 0 (0) | ≤0.001 | |
| Environment | 0 (0) | ||||
| Straw bedding | 40 | 5 (12.5) | 0 (0) | 0.02 | |
| Total | 120 | 20 (16.6) | 0 (0) | ≤0.001 | |
| Household III | Animal samples | ||||
| Fecal samples | 40 | 4 (10) | 0 (0) | 0.04 | |
| Milk | 40 | 0 (0) | 0 (0) | - | |
| Total | 80 | 4 (5) | 0 (0) | 0.043 | |
| Environment | |||||
| Straw bedding | 40 | 3 (7.5) | 0 (0) | 0.077 | |
| Total | 120 | 7 (5.8) | 0 (0) | 0.007 | |
| Total prevalence | Animal samples | ||||
| Fecal samples | 120 | 24 (20) | 3 (2.5) | ≤0.001 | |
| Milk | 120 | 9 (7.5) | 0 (0) | 0.002 | |
| Total | 240 | 33 (13.7) | 3 (1.2) | ≤0.001 | |
| Environment | |||||
| Straw bedding | 120 | 15 (12.5) | 2 (1.6) | 0.001 | |
| Total | 360 | 48 (13.3) | 5 (5.3) | ≤0.001 |
| Antibiotics | Sensitive No. (%) | Resistant No. (%) |
|---|---|---|
| Imipenem | 4/48 (8.3%) | 44/48 (91.6%) |
| Penicillin G | 2/48 (4.1%) | 46/48 (95.8%) |
| Erythromycin | 29/48 (60.4) | 19/48 (39.6%) |
| Amikacin | 25/48 (52.1%) | 23/48 (47.9%) |
| Streptomycin | 2/48 (4.1%) | 46/48 (95.8%) |
| Gentamicin | 28/48 (58.3%) | 20/48 (41.7%) |
| Vancomycin | 47/48 (97.9%) | 1/48 (2.1%) |
| SXT 1 | 0 (0%) | 48/48 (100%) |
| Levofloxacin | 40/49 (81.6%) | 8/48 (16.6%) |
| Samples ID | Sample Source | Distribution of Virulence Gene | Antimicrobial Profile | MAR Index | ||
|---|---|---|---|---|---|---|
| iap | hylA | actA | ||||
| 5 | Feces | + | + | IMP, Pen G, STM, SXT, E, AK, LEV | 0.777 | |
| 7 | Feces | + | IMP, Pen G, STM, SXT, E | 0.555 | ||
| 10 | Feces | + | IMP, Pen G, STM, SXT, AK | 0.555 | ||
| 11 | Feces | + | IMP, Pen G, STM, SXT, AK, G | 0.666 | ||
| 13 | Feces | + | IMP, Pen G, STM, SXT, G | 0.555 | ||
| 17 | Feces | + | IMP, Pen G, STM, SXT, E, G | 0.666 | ||
| 19 | Feces | + | IMP, Pen G, STM, SXT, AK | 0.555 | ||
| 22 | Feces | + | + | IMP, Pen G, STM, SXT, AK, G | 0.666 | |
| 28 | Feces | + | + | IMP, Pen G, STM, SXT, AK, G, LEV | 0.777 | |
| 43 | Milk | + | + | IMP, Pen G, STM, SXT, E, AK, LEV | 0.777 | |
| 48 | Milk | + | IMP, Pen G, STM, SXT, G | 0.555 | ||
| 76 | Milk | + | IMP, Pen G, STM, SXT, VA, E, G | 0.777 | ||
| 78 | Milk | + | IMP, Pen G, STM, SXT, E, G | 0.666 | ||
| 79 | Milk | + | + | + | IMP, Pen G, STM, SXT, E, AK, G, LEV | 0.888 |
| 89 | Bedding | + | + | IMP, Pen G, STM, SXT, AK | 0.555 | |
| 90 | Bedding | + | IMP, Pen G, STM, SXT, LEV | 0.555 | ||
| 101 | Bedding | + | IMP, Pen G, STM, SXT, G | 0.555 | ||
| 107 | Bedding | + | IMP, Pen G, STM, SXT, E | 0.555 | ||
| 110 | Bedding | + | + | IMP, Pen G, STM, SXT, AK | 0.555 | |
| 112 | Bedding | + | + | + | IMP, Pen G, STM, SXT, E, AK, G | 0.777 |
| 113 | Bedding | + | IMP, Pen G, STM, SXT, G | 0.555 | ||
| Prevalence of virulence genes in Household I no. (%) p-value ≤ 0.001 * | iap 21 (100) | hylA 8 (38) | actA 2 (9.5) | Collective MAR index in Household I = 0.644 | ||
| 9 | Feces | + | IMP, Pen G, STM, SXT, | 0.444 | ||
| 12 | Feces | + | + | IMP, Pen G, STM, SXT, AK | 0.555 | |
| 15 | Feces | + | IMP, Pen G, STM, SXT, G, LEV | 0.666 | ||
| 20 | Feces | + | IMP, Pen G, STM, SXT, E | 0.555 | ||
| 23 | Feces | + | + | IMP, Pen G, STM, SXT, AK, G | 0.666 | |
| 28 | Feces | + | + | IMP, Pen G, STM, SXT, E, AK | 0.666 | |
| 30 | Feces | + | IMP, Pen G, STM, SXT, E | 0.555 | ||
| 34 | Feces | + | + | IMP, Pen G, STM, SXT, AK | 0.555 | |
| 37 | Feces | + | IMP, Pen G, STM, SXT | 0.444 | ||
| 38 | Feces | + | IMP, Pen G, STM, SXT, AK | 0.555 | ||
| 40 | Feces | + | IMP, Pen G, STM, SXT, G, LEV | 0.666 | ||
| 49 | Milk | + | + | + | IMP, Pen G, STM, SXT, E, AK | 0.666 |
| 50 | Milk | + | + | IMP, Pen G, STM, SXT, E, AK | 0.666 | |
| 56 | Milk | + | + | IMP, Pen G, STM, SXT, AK | 0.555 | |
| 70 | Milk | + | IMP, Pen G, STM, SXT, E, G | 0.666 | ||
| 87 | Bedding | + | + | IMP, Pen G, STM, SXT, AK | 0.555 | |
| 105 | Bedding | + | IMP, Pen G, STM, SXT, AK, G | 0.666 | ||
| 107 | Bedding | + | + | + | IMP, Pen G, STM, SXT, E, AK, G | 0.777 |
| 111 | Bedding | + | IMP, Pen G, STM, SXT, E, G | 0.666 | ||
| 112 | Bedding | + | + | IMP, Pen G, STM, SXT, AK, G | 0.666 | |
| Prevalence of virulence genes in Household II no. (%) at p-value ≤ 0.001 * | iap 20 (100) | hylA 9 (45) | actA 3 (15) | Collective MAR index in Household II = 0.610 | ||
| 1 | Feces | + | IMP, STM, SXT, | 0.333 | ||
| 3 | Feces | + | + | Pen G, STM, SXT, E | 0.444 | |
| 5 | Feces | + | + | IMP, Pen G, SXT, E | 0.444 | |
| 7 | Feces | + | + | STM, SXT, LEV | 0.333 | |
| 90 | Bedding | + | IMP, Pen G, SXT, | 0.333 | ||
| 91 | Bedding | + | + | Pen G, STM, SXT, | 0.333 | |
| 93 | Bedding | + | Pen G, STM, SXT, | 0.333 | ||
| Prevalence of virulence genes in Household III no. (%) at p-value 0.011 * | iap 7 (100) | hylA 2 (28.5) | actA 2 (28.5) | Collective MAR index in Household III = 0.364 | ||
| Microorganisms | Rose Water (mg/L) | Rose Oil (mg/L) | Orange Oil (mg/L) | |||
|---|---|---|---|---|---|---|
| MIC | MMC | MIC | MMC | MIC | MMC | |
| L. monocytogenes | 10 | 10 | 2.5 | 20 | 7.8 | 7.8 |
| C. sakazakii | 20 | 20 | 20 | 40 | 7.8 | 7.8 |
| Target Gene | Oligonucleotide Sequence (5′ → 3′) | Virulence Factor | Size (bp) | Reference |
|---|---|---|---|---|
| iap (F) | ACAAGCTGCACCTGTTGCAG | Invasive associated protein | 131 | [12] |
| iap (R) | TGACAGCGTGTGTAGTAGCA | |||
| hlyA (F) | GCAGTTGCAAGCGCTTGGAGTGAA | Hemolysin | 456 | [12] |
| hlyA (R) | GCAACGTATCCTCCAGAGTGATCG | |||
| actA (F) | CGCCGCGGAAATTAAAAAAAGA | Actin polymerization protein | 839 | [26] |
| actA (R) | ACGAAGGAACCGGGCTGCTAG | |||
| cgcA (F) | GGCGGACGAAGCCTCAGAGAGT | diguanylate cyclase -Encoding Gene, cgcA (species specific) | 492 | [49] |
| cgcA (R) | TTAGGGCCATTCGGAAATCCGAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badawy, B.; Gwida, M.; Sadat, A.; EL-Toukhy, M.; Sayed-Ahmed, M.; Alam, N.; Ahmad, S.; Ali, M.S.; Elafify, M. Prevalence and Antimicrobial Resistance of Virulent Listeria monocytogenes and Cronobacter sakazakii in Dairy Cattle, the Environment, and Dried Milk with the In Vitro Application of Natural Alternative Control. Antibiotics 2022, 11, 1087. https://doi.org/10.3390/antibiotics11081087
Badawy B, Gwida M, Sadat A, EL-Toukhy M, Sayed-Ahmed M, Alam N, Ahmad S, Ali MS, Elafify M. Prevalence and Antimicrobial Resistance of Virulent Listeria monocytogenes and Cronobacter sakazakii in Dairy Cattle, the Environment, and Dried Milk with the In Vitro Application of Natural Alternative Control. Antibiotics. 2022; 11(8):1087. https://doi.org/10.3390/antibiotics11081087
Chicago/Turabian StyleBadawy, Basma, Mayada Gwida, Asmaa Sadat, Marwa EL-Toukhy, Mohamed Sayed-Ahmed, Nawazish Alam, Sarfaraz Ahmad, MD Sajid Ali, and Mahmoud Elafify. 2022. "Prevalence and Antimicrobial Resistance of Virulent Listeria monocytogenes and Cronobacter sakazakii in Dairy Cattle, the Environment, and Dried Milk with the In Vitro Application of Natural Alternative Control" Antibiotics 11, no. 8: 1087. https://doi.org/10.3390/antibiotics11081087
APA StyleBadawy, B., Gwida, M., Sadat, A., EL-Toukhy, M., Sayed-Ahmed, M., Alam, N., Ahmad, S., Ali, M. S., & Elafify, M. (2022). Prevalence and Antimicrobial Resistance of Virulent Listeria monocytogenes and Cronobacter sakazakii in Dairy Cattle, the Environment, and Dried Milk with the In Vitro Application of Natural Alternative Control. Antibiotics, 11(8), 1087. https://doi.org/10.3390/antibiotics11081087