
Bioinformatics and Genomic Analyses of the Suitability of Eight Riboswitches for Antibacterial Drug Targets

Abstract
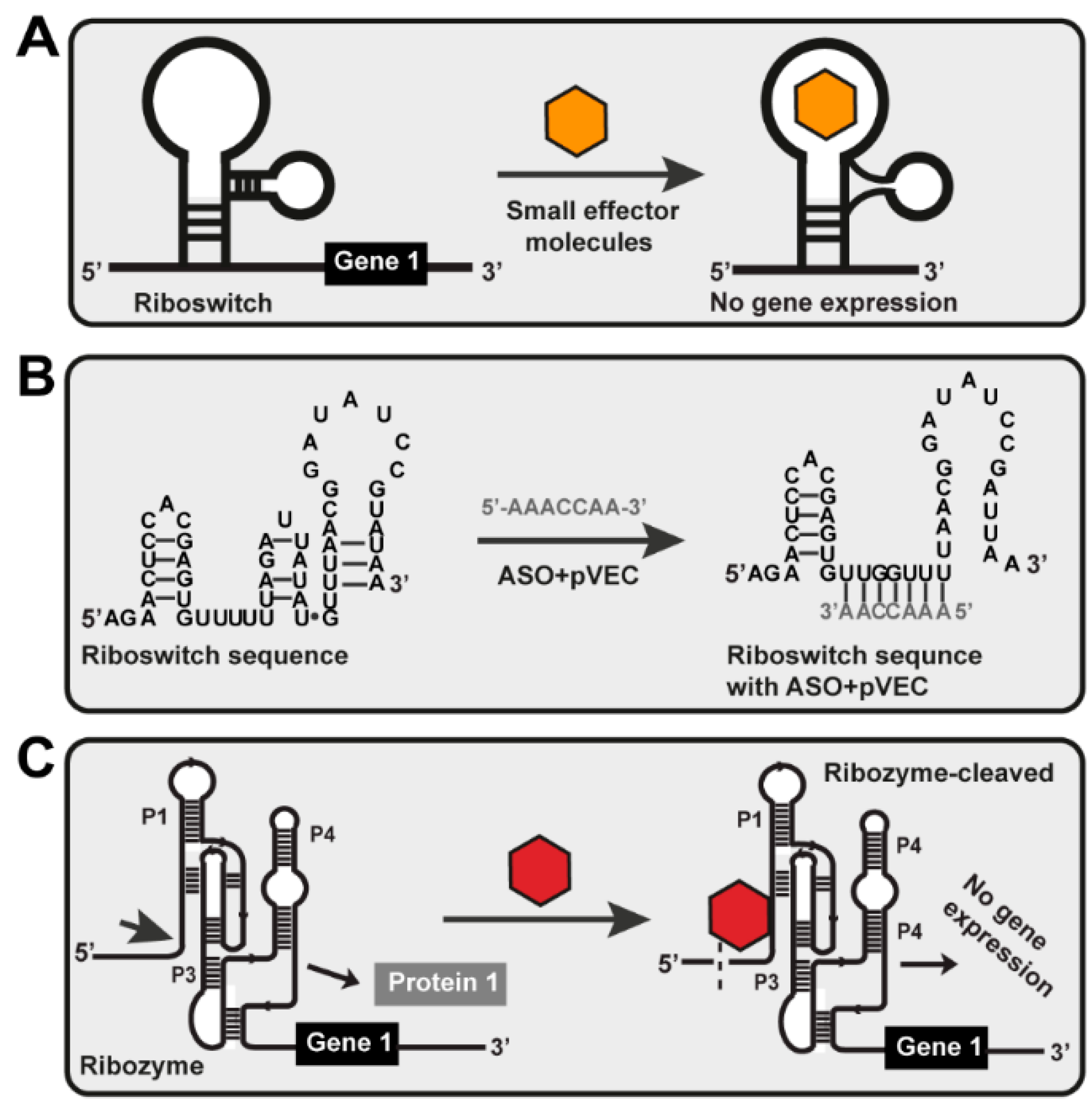
:1. Introduction
2. Results
2.1. Most Suitable Riboswitches
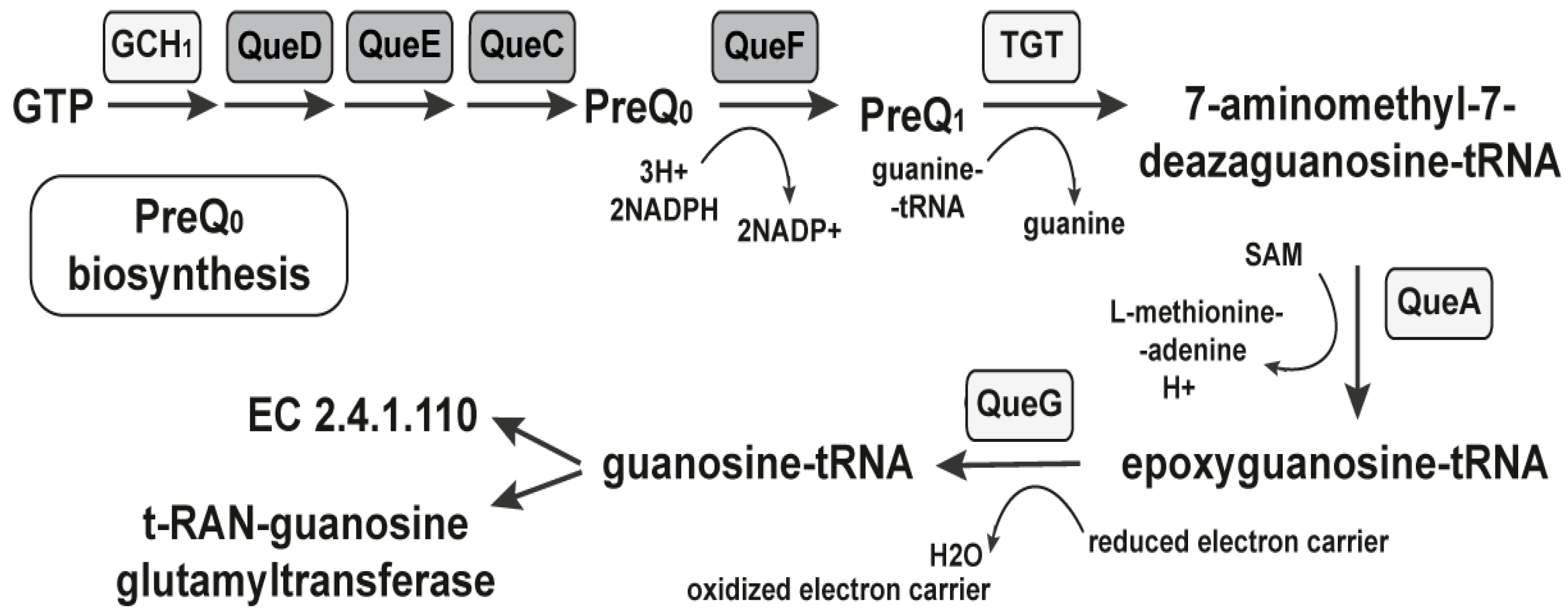
2.1.1. The PreQ1-I Riboswitch
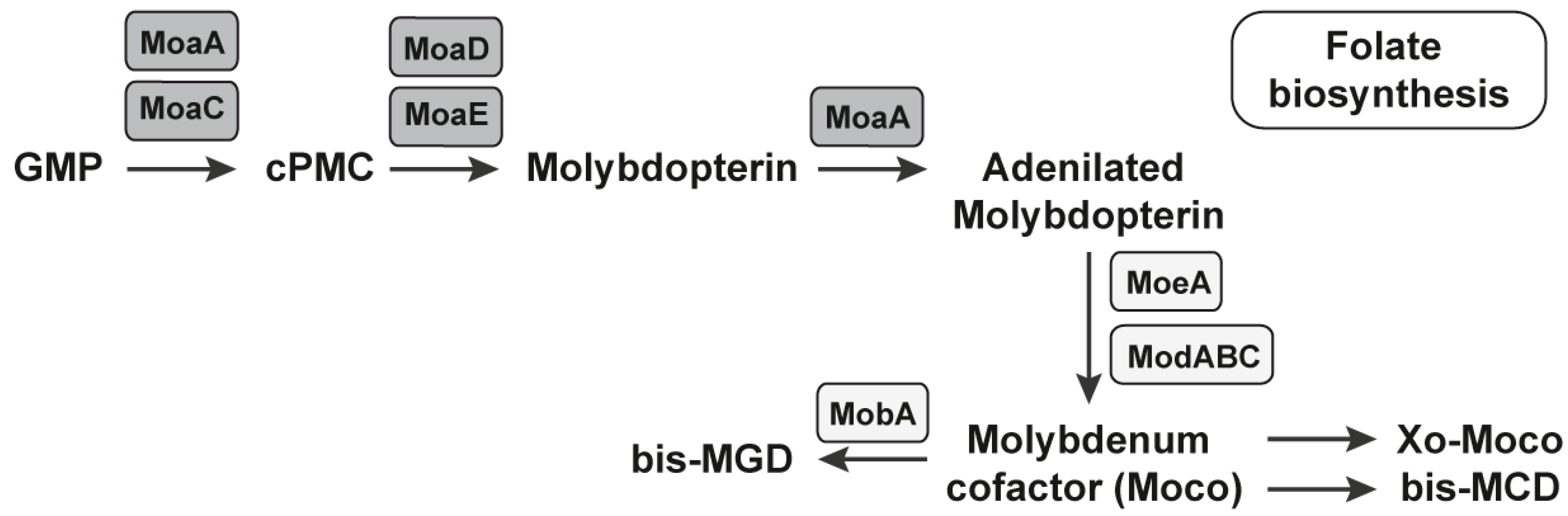
2.1.2. The MoCo RNA Motif Riboswitch
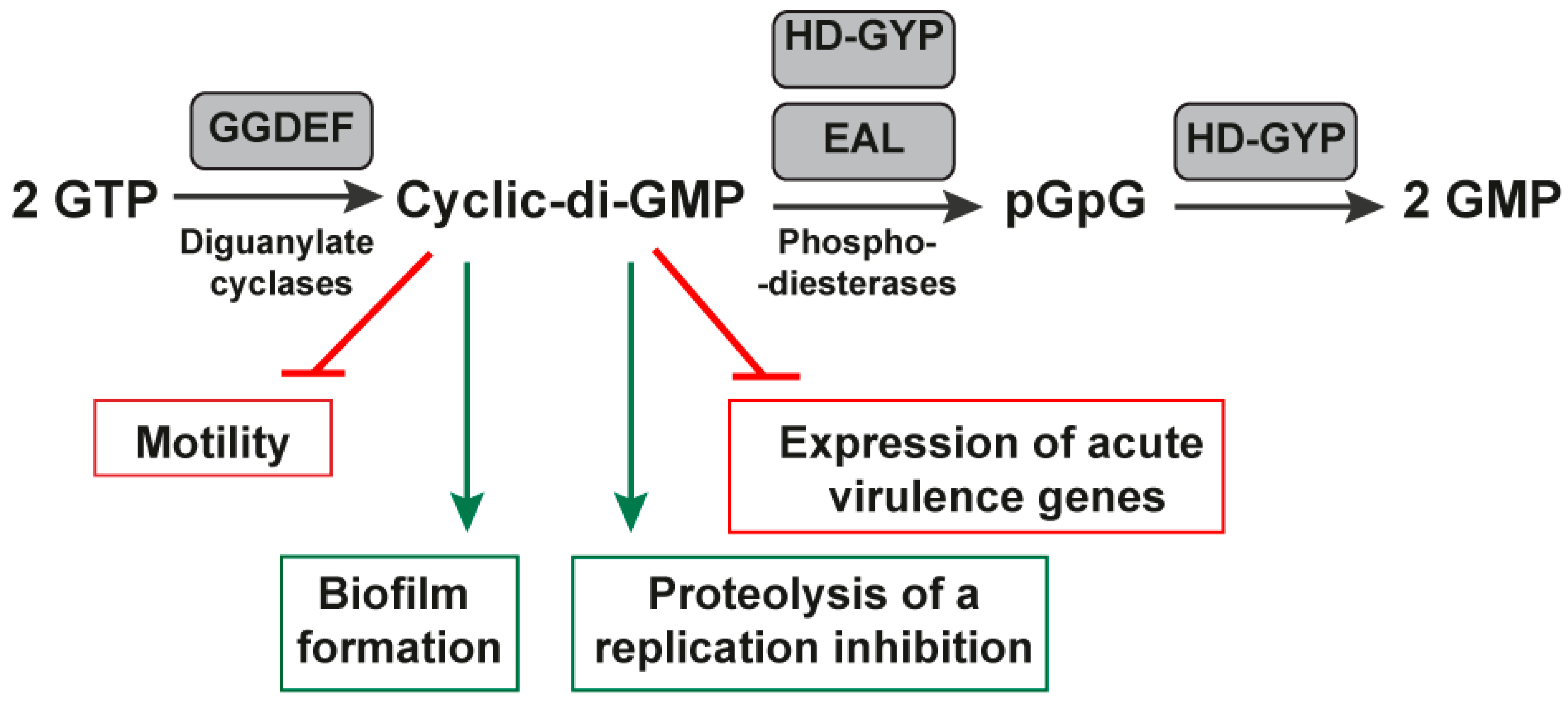
2.2. Very Suitable Riboswitches:
Cyclic-Di-GMP-I and Cyclic-Di-GMP-II Riboswitches
2.3. Suitable Riboswitches
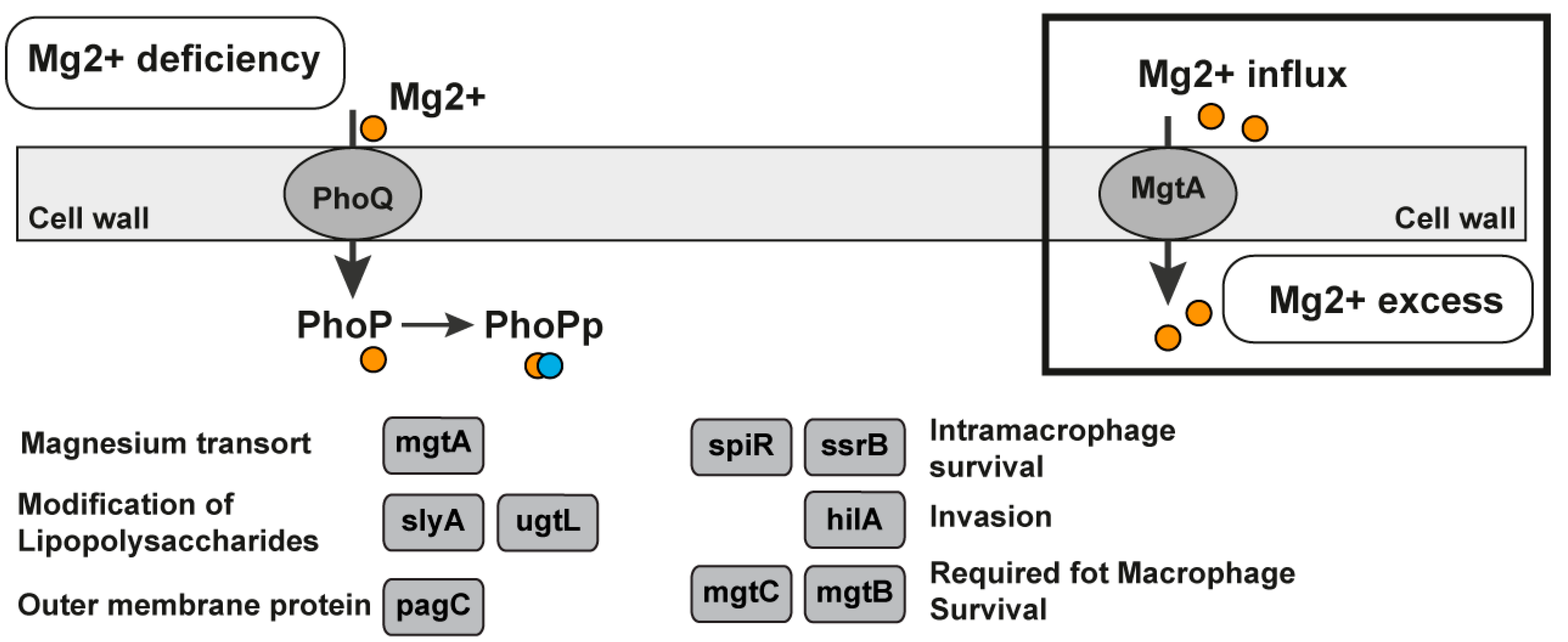
Mg2+ Sensor Riboswitch
2.4. Not Suitable Riboswitches
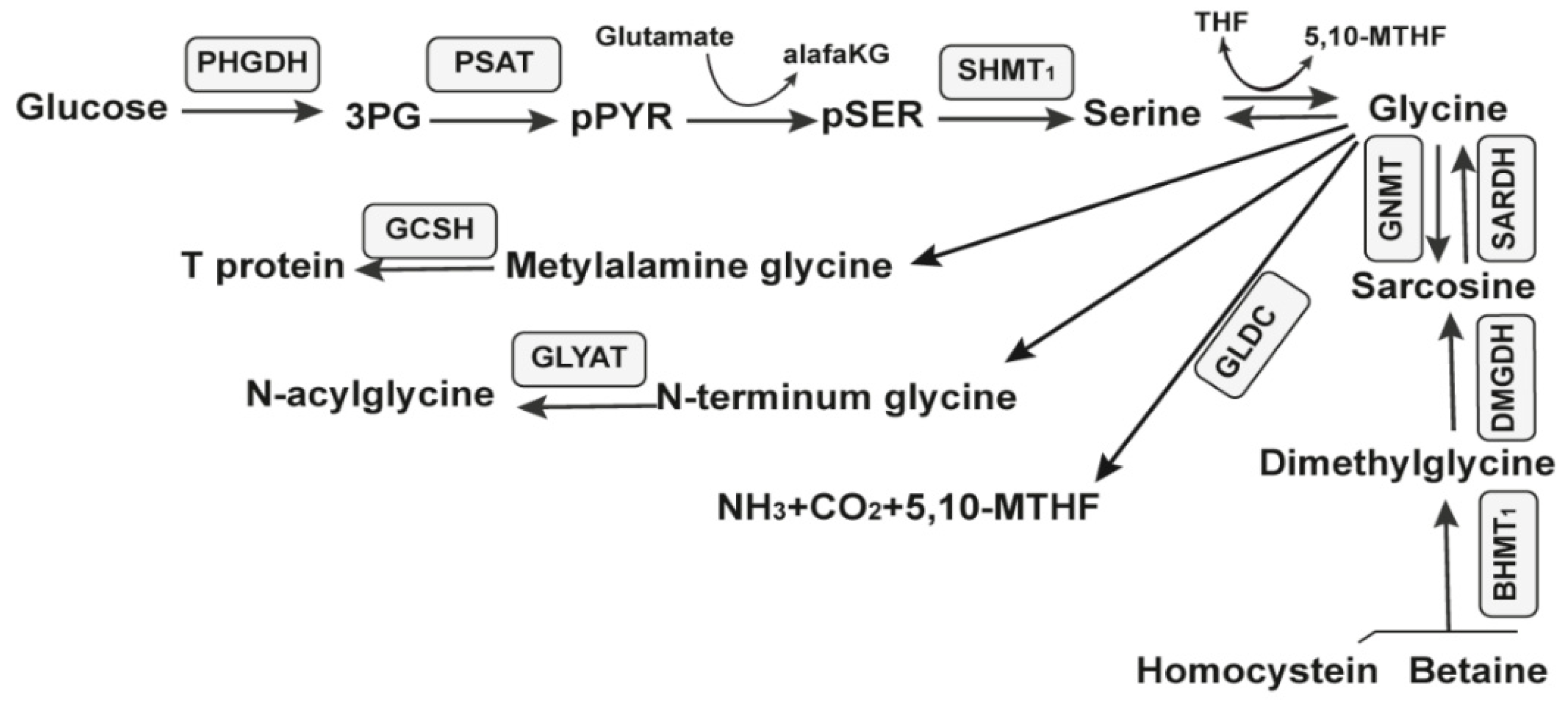
2.4.1. The Glycine Riboswitch
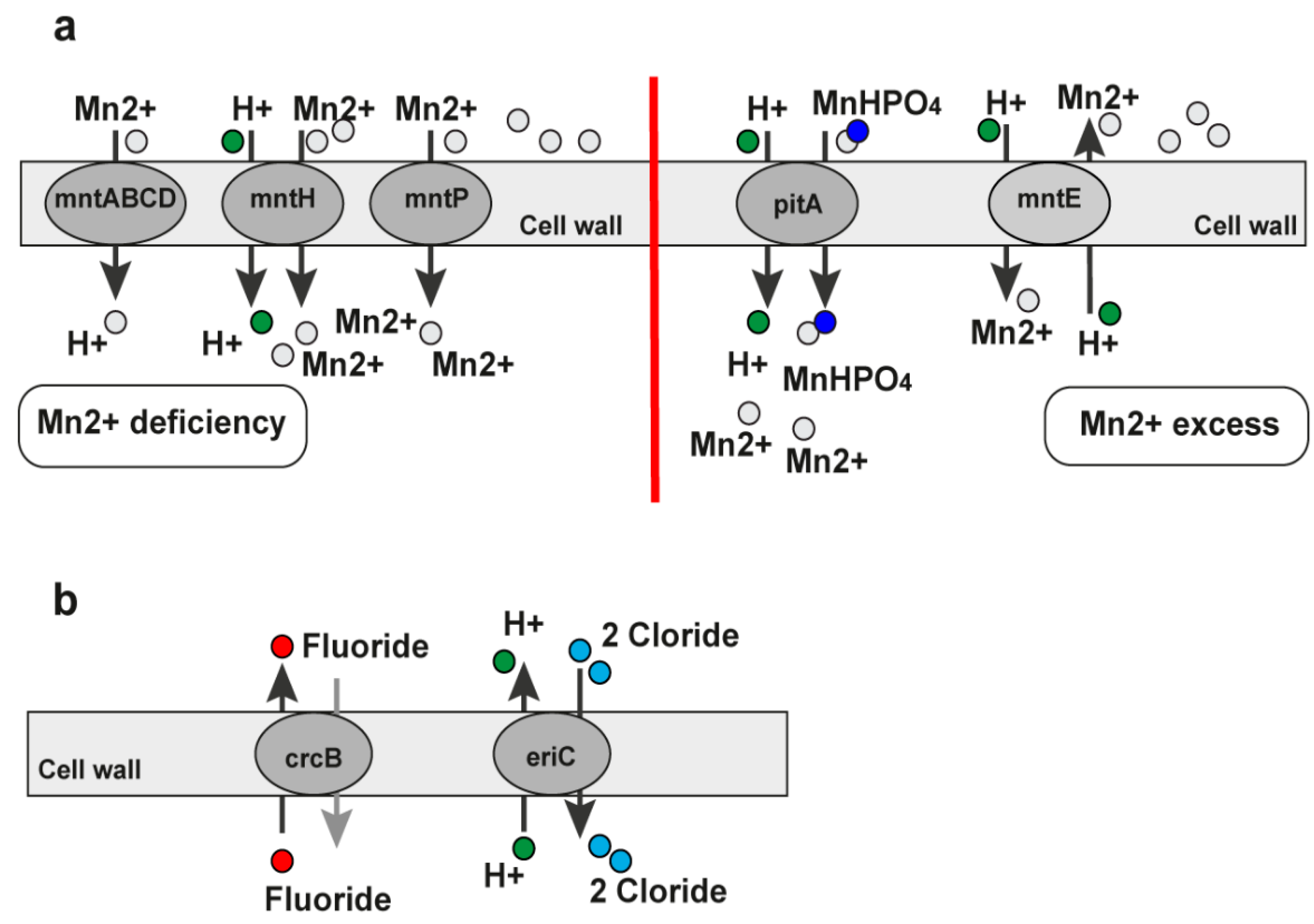
2.4.2. The Mn2+ Riboswitch
2.4.3. The Fluoride Riboswitch
3. Discussion
4. Materials and Methods
4.1. Databases Used
4.2. Software Used
4.3. Evaluation procedure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Teodoro, A.; Goncalves, F.J.M.; Oliveira, H.; Marques, S. Venom of Viperidae: A Perspective of its Antibacterial and Antitumor Potential. Curr. Drug Targets 2022, 23, 126–144. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Webster, T.J. Bacteria antibiotic resistance: New challenges and opportunities for implant-associated orthopedic infections. J. Orthop. Res. 2018, 36, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Eleraky, N.E.; Allam, A.; Hassan, S.B.; Omar, M.M. Nanomedicine Fight against Antibacterial Resistance: An Overview of the Recent Pharmaceutical Innovations. Pharmaceutics 2020, 12, 142. [Google Scholar] [CrossRef] [PubMed]
- Ungureanu, A.; Zlatian, O.; Mitroi, G.; Drocaş, A.; Ţîrcă, T.; Călina, D.; Dehelean, C.; Docea, A.O.; Izotov, B.N.; Rakitskii, V.N.; et al. Staphylococcus aureus colonisation in patients from a primary regional hospital. Mol. Med. Rep. 2017, 16, 8771–8780. [Google Scholar] [CrossRef]
- Jin, H.; Qiu, F.; Ji, H.J.; Lu, Q. Analysis of drug resistance in 1861 strains of Acinetobacter baumannii. Biomed. Rep. 2016, 4, 463–466. [Google Scholar] [CrossRef]
- Willyard, C. The drug-resistant bacteria that pose the greatest health threats. Nature 2017, 543, 15. [Google Scholar] [CrossRef]
- Pavlova, N.; Penchovsky, R. Genome-wide bioinformatics analysis of FMN, SAM-I, glmS, TPP, lysine, purine, cobalamin, and SAH riboswitches for their applications as allosteric antibacterial drug targets in human pathogenic bacteria. Expert Opin. Ther. Targets 2019, 23, 631–643. [Google Scholar] [CrossRef]
- Călina, D.; Docea, A.O.; Rosu, L.; Zlatian, O.; Rosu, A.F.; Anghelina, F.; Rogoveanu, O.; Arsene, A.L.; Nicolae, A.C.; Drăgoi, C.M.; et al. Antimicrobial resistance development following surgical site infections. Mol. Med. Rep. 2017, 15, 681–688. [Google Scholar] [CrossRef]
- Thu, H.E.; Hussain, Z.; Mohamed, I.N.; Shuid, A.N. Recent Advances in Antibacterial, Antiprotozoal and Antifungal Trends of Eurycoma longifolia: A Review of Therapeutic Implications and Future Prospects. Curr. Drug Targets 2018, 19, 1657–1671. [Google Scholar] [CrossRef]
- Prosperi, M.; Marini, S. KARGA: Multi-platform Toolkit for k-mer-based Antibiotic Resistance Gene Analysis of High-throughput Sequencing Data. In Proceedings of the 2021 IEEE EMBS International Conference on Biomedical and Health Informatics (BHI), Athens, Greece, 27–30 July 2021. [Google Scholar]
- Pandey, S.; Kong, T.; Backes, N.; Phillips, G.J. Open Droplet Microfluidics for Testing Multi-Drug Resistance and Antibiotic Resilience in Bacteria. In Proceedings of the 2021 21st International Conference on Solid-State Sensors, Actuators and Microsystems (Transducers), Orlando, FL, USA, 20–24 June 2021. [Google Scholar]
- Robson, B. Computers and preventative diagnosis. A survey with bioinformatics examples of mitochondrial small open reading frame peptides as portents of a new generation of powerful biomarkers. Comput. Biol. Med. 2022, 140, 105116. [Google Scholar] [CrossRef] [PubMed]
- Medina-Pizzali, M.L.; Venkatesh, A.; Riveros, M.; Cuicapuza, D.; Salmon-Mulanovich, G.; Mäusezahl, D.; Hartinger, S.M. Whole-Genome Characterisation of ESBL-Producing E. coli Isolated from Drinking Water and Dog Faeces from Rural Andean Households in Peru. Antibiotics 2022, 11, 692. [Google Scholar] [CrossRef]
- Losito, A.R.; Raffaelli, F.; Del Giacomo, P.; Tumbarello, M. New Drugs for the Treatment of Pseudomonas aeruginosa Infections with Limited Treatment Options: A Narrative Review. Antibiotics 2022, 11, 579. [Google Scholar] [CrossRef] [PubMed]
- Popova, K.B.; Valsamatzi-Panagiotou, A.; Penchovsky, R. New drug discovery strategies for targeting drug-resistant bacteria. Environ. Chem. Lett. 2021, 19, 1995–2004. [Google Scholar] [CrossRef]
- Panchal, V.; Brenk, R. Riboswitches as Drug Targets for Antibiotics. Antibiotics 2021, 10, 45. [Google Scholar] [CrossRef]
- Nudler, E.; Mironov, A.S. The riboswitch control of bacterial metabolism. Trends Biochem. Sci. 2004, 29, 11–17. [Google Scholar] [CrossRef]
- Tucker, B.J.; Breaker, R.R. Riboswitches as versatile gene control elements. Curr. Opin. Struct. Biol. 2005, 15, 342–348. [Google Scholar] [CrossRef]
- Mehta, N.B.; Balaji, P. Riboswitches: Classification, function and insilico approach. Int. J. 2010, 1, 409–420. [Google Scholar]
- Winkler, W.C.; Breaker, R.R. Genetic control by metabolite-binding riboswitches. Chembiochem 2003, 4, 1024–1032. [Google Scholar] [CrossRef]
- Mandal, M.; Breaker, R.R. Gene regulation by riboswitches. Nat. Rev. Mol. Cell Biol. 2004, 5, 451–463. [Google Scholar] [CrossRef]
- Penchovsky, R.; Traykovska, M. Designing drugs that overcome antibacterial resistance: Where do we stand and what should we do? Expert Opin. Drug Discov. 2015, 10, 631–650. [Google Scholar] [CrossRef] [PubMed]
- McCown, P.J.; Corbino, K.A.; Stav, S.; Sherlock, M.E.; Breaker, R.R. Riboswitch diversity and distribution. RNA 2017, 23, 995–1011. [Google Scholar] [CrossRef] [PubMed]
- Penchovsky, R.; Stoilova, C.C. Riboswitch-based antibacterial drug discovery using high-throughput screening methods. Expert Opin. Drug Discov. 2013, 8, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Mandal, M.; Lee, M.; Barrick, J.E.; Weinberg, Z.; Emilsson, G.M.; Ruzzo, W.L.; Breaker, R.R. A glycine-dependent riboswitch that uses cooperative binding to control gene expression. Science 2004, 306, 275–279. [Google Scholar] [CrossRef]
- Bastet, L.; Chauvier, A.; Singh, N.; Lussier, A.; Lamontagne, A.M.; Prevost, K.; Masse, E.; Wade, J.T.; Lafontaine, D.A. Translational control and Rho-dependent transcription termination are intimately linked in riboswitch regulation. Nucleic Acids Res. 2017, 45, 7474–7486. [Google Scholar] [CrossRef]
- Hollands, K.; Proshkin, S.; Sklyarova, S.; Epshtein, V.; Mironov, A.; Nudler, E.; Groisman, E.A. Riboswitch control of Rho-dependent transcription termination. Proc. Natl. Acad. Sci. USA 2012, 109, 5376–5381. [Google Scholar] [CrossRef]
- Loh, E.; Dussurget, O.; Gripenland, J.; Vaitkevicius, K.; Tiensuu, T.; Mandin, P.; Repoila, F.; Buchrieser, C.; Cossart, P.; Johansson, J. A trans-acting riboswitch controls expression of the virulence regulator PrfA in Listeria monocytogenes. Cell 2009, 139, 770–779. [Google Scholar] [CrossRef]
- Deigan, K.E.; Ferré-D’Amaré, A.R. Riboswitches: Discovery of drugs that target bacterial gene-regulatory RNAs. Acc. Chem. Res. 2011, 44, 1329–1338. [Google Scholar] [CrossRef]
- Blount, K.F.; Wang, J.X.; Lim, J.; Sudarsan, N.; Breaker, R.R. Antibacterial lysine analogs that target lysine riboswitches. Nat. Chem. Biol. 2007, 3, 44–49. [Google Scholar] [CrossRef]
- Edwards, T.E.; Ferre-D’Amare, A.R. Crystal structures of the thi-box riboswitch bound to thiamine pyrophosphate analogs reveal adaptive RNA-small molecule recognition. Structure 2006, 14, 1459–1468. [Google Scholar] [CrossRef]
- Shahidi, S.; Shahraeini, S.S.; Farahani, Y.F.; Sardari, S. Thiamine pyrophosphate riboswitch regulation: A new possible mechanism involved in the action of nalidixic acid. Turk. J. Biochem. 2020, 45, 777–784. [Google Scholar] [CrossRef]
- Matern, A.; Pedrolli, D.; Grosshennig, S.; Johansson, J.; Mack, M. Uptake and Metabolism of Antibiotics Roseoflavin and 8-Demethyl-8-Aminoriboflavin in Riboflavin-Auxotrophic Listeria monocytogenes. J. Bacteriol. 2016, 198, 3233–3243. [Google Scholar] [CrossRef] [PubMed]
- Serganov, A.; Huang, L.; Patel, D.J. Coenzyme recognition and gene regulation by a flavin mononucleotide riboswitch. Nature 2009, 458, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.R.; Blount, K.F.; Breaker, R.R. Roseoflavin is a natural antibacterial compound that binds to FMN riboswitches and regulates gene expression. RNA Biol. 2009, 6, 187–194. [Google Scholar] [CrossRef]
- Mora-Lugo, R.; Stegmuller, J.; Mack, M. Metabolic engineering of roseoflavin-overproducing microorganisms. Microb. Cell Fact. 2019, 18, 146. [Google Scholar] [CrossRef]
- Blount, K.; Puskarz, I.; Penchovsky, R.; Breaker, R. Development and application of a high-throughput assay for glmS riboswitch activators. RNA Biol. 2006, 3, 77–81. [Google Scholar] [CrossRef]
- Penchovsky, R. Computational design and biosensor applications of small molecule-sensing allosteric ribozymes. Biomacromolecules 2013, 14, 1240–1249. [Google Scholar] [CrossRef]
- Felden, B.; Cattoir, V. Bacterial Adaptation to Antibiotics through Regulatory RNAs. Antimicrob. Agents Chemother. 2018, 62, e02503–e02517. [Google Scholar] [CrossRef]
- Penchovsky, R. Computational design of allosteric ribozymes as molecular biosensors. Biotechnol. Adv. 2014, 32, 1015–1027. [Google Scholar] [CrossRef]
- Penchovsky, R.; Breaker, R.R. Computational design and experimental validation of oligonucleotide-sensing allosteric ribozymes. Nat. Biotechnol. 2005, 23, 1424–1433. [Google Scholar] [CrossRef]
- Li, S.; Breaker, R.R. Eukaryotic TPP riboswitch regulation of alternative splicing involving long-distance base pairing. Nucleic Acids Res. 2013, 41, 3022–3031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traykovska, M.; Popova, K.B.; Penchovsky, R. Targeting glmS Ribozyme with Chimeric Antisense Oligonucleotides for Antibacterial Drug Development. ACS Synth. Biol. 2021, 10, 3167–3176. [Google Scholar] [CrossRef] [PubMed]
- Traykovska, M.; Penchovsky, R. Engineering Antisense Oligonucleotides as Antibacterial Agents That Target FMN Riboswitches and Inhibit the Growth of Staphylococcus aureus, Listeria monocytogenes, and Escherichia coli. ACS Synth. Biol. 2022, 11, 1845–1855. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.; Yao, R.; Gomes, M.; Jørgensen, P.T.; Wengel, J.; Azevedo, N.F.; Sobral Santos, R. Can Vitamin B12 Assist the Internalization of Antisense LNA Oligonucleotides into Bacteria? Antibiotics 2021, 10, 379. [Google Scholar] [CrossRef]
- Valsamatzi-Panagiotou, A.; Popova, K.B.; Penchovsky, R. Methods for prevention and constraint of antimicrobial resistance: A review. Environ. Chem. Lett. 2021, 19, 2005–2012. [Google Scholar] [CrossRef]
- Yu, C.; Yu, Y.; Xu, Z.; Li, H.; Yang, D.; Xiang, M.; Zuo, Y.; Li, S.; Chen, Z.; Yu, Z. Antisense oligonucleotides targeting human telomerase mRNA increases the radiosensitivity of nasopharyngeal carcinoma cells. Mol. Med. Rep. 2015, 11, 2825–2830. [Google Scholar] [CrossRef]
- Funato, T.; Satou, J.; Kozawa, K.; Fujimaki, S.; Miura, T.; Kaku, M. Use of c-myb antisense oligonucleotides to increase the sensitivity of human colon cancer cells to cisplatin. Oncol. Rep. 2001, 8, 807–810. [Google Scholar] [CrossRef]
- Abbadi, M.; Spurgeon, S.; Khan, N.; Warren, M. Understanding the Control of a Vitamin B12 Riboswitch. In Proceedings of the 2018 UKACC 12th International Conference on Control (CONTROL), Sheffield, UK, 5–7 September 2018. [Google Scholar]
- McCown, P.J.; Liang, J.J.; Weinberg, Z.; Breaker, R.R. Structural, functional, and taxonomic diversity of three preQ1 riboswitch classes. Chem. Biol. 2014, 21, 880–889. [Google Scholar] [CrossRef]
- Liberman, J.A.; Suddala, K.C.; Aytenfisu, A.; Chan, D.; Belashov, I.A.; Salim, M.; Mathews, D.H.; Spitale, R.C.; Walter, N.G.; Wedekind, J.E. Structural analysis of a class III preQ1 riboswitch reveals an aptamer distant from a ribosome-binding site regulated by fast dynamics. Proc. Natl. Acad. Sci. USA 2015, 112, E3485–E3494. [Google Scholar] [CrossRef]
- Roth, A.; Winkler, W.C.; Regulski, E.E.; Lee, B.W.; Lim, J.; Jona, I.; Barrick, J.E.; Ritwik, A.; Kim, J.N.; Welz, R.; et al. A riboswitch selective for the queuosine precursor preQ1 contains an unusually small aptamer domain. Nat. Struct. Mol. Biol. 2007, 14, 308–317. [Google Scholar] [CrossRef]
- Klein, D.J.; Edwards, T.E.; Ferre-D’Amare, A.R. Cocrystal structure of a class I preQ1 riboswitch reveals a pseudoknot recognizing an essential hypermodified nucleobase. Nat. Struct. Mol. Biol. 2009, 16, 343–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.; Peterson, R.; Feigon, J. Structural Insights into riboswitch control of the biosynthesis of queuosine, a modified nucleotide found in the anticodon of tRNA. Mol. Cell 2009, 33, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, N.; Kaloudas, D.; Penchovsky, R. Riboswitch distribution, structure, and function in bacteria. Gene 2019, 708, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Reader, J.S.; Metzgar, D.; Schimmel, P.; de Crecy-Lagard, V. Identification of four genes necessary for biosynthesis of the modified nucleoside queuosine. J. Biol. Chem. 2004, 279, 6280–6285. [Google Scholar] [CrossRef]
- Kim, J.N.; Roth, A.; Breaker, R.R. Guanine riboswitch variants from Mesoplasma florum selectively recognize 2′-deoxyguanosine. Proc. Natl. Acad. Sci. USA 2007, 104, 16092–16097. [Google Scholar] [CrossRef]
- Eichhorn, C.D.; Kang, M.; Feigon, J. Structure and function of preQ1 riboswitches. Biochim. Biophys Acta 2014, 1839, 939–950. [Google Scholar] [CrossRef]
- Rieder, U.; Kreutz, C.; Micura, R. Folding of a transcriptionally acting preQ1 riboswitch. Proc. Natl. Acad. Sci. USA 2010, 107, 10804–10809. [Google Scholar] [CrossRef]
- Suddala, K.C.; Wang, J.; Hou, Q.; Walter, N.G. Mg(2+) shifts ligand-mediated folding of a riboswitch from induced-fit to conformational selection. J. Am. Chem. Soc. 2015, 137, 14075–14083. [Google Scholar] [CrossRef]
- Suddala, K.C.; Rinaldi, A.J.; Feng, J.; Mustoe, A.M.; Eichhorn, C.D.; Liberman, J.A.; Wedekind, J.E.; Al-Hashimi, H.M.; Brooks, C.L., 3rd; Walter, N.G. Single transcriptional and translational preQ1 riboswitches adopt similar pre-folded ensembles that follow distinct folding pathways into the same ligand-bound structure. Nucleic Acids Res. 2013, 41, 10462–10475. [Google Scholar] [CrossRef]
- Kuchino, Y.; Kasai, H.; Nihei, K.; Nishimura, S. Biosynthesis of the modified nucleoside Q in transfer RNA. Nucleic Acids Res. 1976, 3, 393–398. [Google Scholar] [CrossRef]
- Kierzek, E.; Kierzek, R. Conscious uncoupling of riboswitch functions. J. Biol. Chem. 2020, 295, 2568–2569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhagdikar, D.; Grundy, F.J.; Henkin, T.M. Transcriptional and translational S-box riboswitches differ in ligand-binding properties. J. Biol. Chem. 2020, 295, 6849–6860. [Google Scholar] [CrossRef] [PubMed]
- Connelly, C.M.; Numata, T.; Boer, R.E.; Moon, M.H.; Sinniah, R.S.; Barchi, J.J.; Ferre-D’Amare, A.R.; Schneekloth, J.S., Jr. Synthetic ligands for PreQ1 riboswitches provide structural and mechanistic insights into targeting RNA tertiary structure. Nat. Commun. 2019, 10, 1501. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, Z.; Barrick, J.E.; Yao, Z.; Roth, A.; Kim, J.N.; Gore, J.; Wang, J.X.; Lee, E.R.; Block, K.F.; Sudarsan, N.; et al. Identification of 22 candidate structured RNAs in bacteria using the CMfinder comparative genomics pipeline. Nucleic Acids Res. 2007, 35, 4809–4819. [Google Scholar] [CrossRef]
- Regulski, E.E.; Moy, R.H.; Weinberg, Z.; Barrick, J.E.; Yao, Z.; Ruzzo, W.L.; Breaker, R.R. A widespread riboswitch candidate that controls bacterial genes involved in molybdenum cofactor and tungsten cofactor metabolism. Mol. Microbiol. 2008, 68, 918–932. [Google Scholar] [CrossRef]
- Leimkuhler, S.; Wuebbens, M.M.; Rajagopalan, K.V. The History of the Discovery of the Molybdenum Cofactor and Novel Aspects of its Biosynthesis in Bacteria. Coord. Chem. Rev. 2011, 255, 1129–1144. [Google Scholar] [CrossRef]
- Schwarz, G.; Mendel, R.R.; Ribbe, M.W. Molybdenum cofactors, enzymes and pathways. Nature 2009, 460, 839–847. [Google Scholar] [CrossRef]
- Johnson, J.L.; Bastian, N.R.; Schauer, N.L.; Ferry, J.G.; Rajagopalan, K.V. Identification of molybdopterin guanine dinucleotide in formate dehydrogenase from Methanobacterium formicicum. FEMS Microbiol. Lett. 1991, 61, 213–216. [Google Scholar] [CrossRef]
- Hover, B.M.; Tonthat, N.K.; Schumacher, M.A.; Yokoyama, K. Mechanism of pyranopterin ring formation in molybdenum cofactor biosynthesis. Proc. Natl. Acad. Sci. USA 2015, 112, 6347–6352. [Google Scholar] [CrossRef]
- Iobbi-Nivol, C.; Leimkuhler, S. Molybdenum enzymes, their maturation and molybdenum cofactor biosynthesis in Escherichia coli. Biochim. Biophys Acta 2013, 1827, 1086–1101. [Google Scholar] [CrossRef]
- Pallegar, P.; Pena-Castillo, L.; Langille, E.; Gomelsky, M.; Lang, A.S. Cyclic di-GMP-Mediated Regulation of Gene Transfer and Motility in Rhodobacter capsulatus. J. Bacteriol. 2020, 202, e00554-19. [Google Scholar] [CrossRef] [PubMed]
- Sudarsan, N.; Lee, E.R.; Weinberg, Z.; Moy, R.H.; Kim, J.N.; Link, K.H.; Breaker, R.R. Riboswitches in eubacteria sense the second messenger cyclic di-GMP. Science 2008, 321, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.D.; Lipchock, S.V.; Ames, T.D.; Wang, J.; Breaker, R.R.; Strobel, S.A. Structural basis of ligand binding by a c-di-GMP riboswitch. Nat. Struct. Mol. Biol. 2009, 16, 1218–1223. [Google Scholar] [CrossRef] [PubMed]
- Kulshina, N.; Baird, N.J.; Ferre-D’Amare, A.R. Recognition of the bacterial second messenger cyclic diguanylate by its cognate riboswitch. Nat. Struct. Mol. Biol. 2009, 16, 1212–1217. [Google Scholar] [CrossRef] [PubMed]
- Kellenberger, C.A.; Wilson, S.C.; Hickey, S.F.; Gonzalez, T.L.; Su, Y.; Hallberg, Z.F.; Brewer, T.F.; Iavarone, A.T.; Carlson, H.K.; Hsieh, Y.F.; et al. GEMM-I riboswitches from Geobacter sense the bacterial second messenger cyclic AMP-GMP. Proc. Natl. Acad. Sci. USA 2015, 112, 5383–5388. [Google Scholar] [CrossRef]
- Nelson, J.W.; Sudarsan, N.; Phillips, G.E.; Stav, S.; Lunse, C.E.; McCown, P.J.; Breaker, R.R. Control of bacterial exoelectrogenesis by c-AMP-GMP. Proc. Natl. Acad. Sci. USA 2015, 112, 5389–5394. [Google Scholar] [CrossRef]
- Lee, E.R.; Baker, J.L.; Weinberg, Z.; Sudarsan, N.; Breaker, R.R. An allosteric self-splicing ribozyme triggered by a bacterial second messenger. Science 2010, 329, 845–848. [Google Scholar] [CrossRef]
- Tamayo, R. Cyclic diguanylate riboswitches control bacterial pathogenesis mechanisms. PLoS Pathog. 2019, 15, e1007529. [Google Scholar] [CrossRef]
- Hall, C.L.; Lee, V.T. Cyclic-di-GMP regulation of virulence in bacterial pathogens. Wiley Interdiscip. Rev. RNA 2018, 9, e1454. [Google Scholar] [CrossRef]
- McKee, R.W.; Harvest, C.K.; Tamayo, R. Cyclic Diguanylate Regulates Virulence Factor Genes via Multiple Riboswitches in Clostridium difficile. mSphere 2018, 3, e00423-18. [Google Scholar] [CrossRef]
- Solano, C.; Garcia, B.; Latasa, C.; Toledo-Arana, A.; Zorraquino, V.; Valle, J.; Casals, J.; Pedroso, E.; Lasa, I. Genetic reductionist approach for dissecting individual roles of GGDEF proteins within the c-di-GMP signaling network in Salmonella. Proc. Natl. Acad. Sci. USA 2009, 106, 7997–8002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romling, U.; Galperin, M.Y.; Gomelsky, M. Cyclic di-GMP: The first 25 years of a universal bacterial second messenger. Microbiol. Mol. Biol. Rev. 2013, 77, 1–52. [Google Scholar] [CrossRef]
- Bordeleau, E.; Fortier, L.C.; Malouin, F.; Burrus, V. c-di-GMP turn-over in Clostridium difficile is controlled by a plethora of diguanylate cyclases and phosphodiesterases. PLoS Genet. 2011, 7, e1002039. [Google Scholar] [CrossRef] [PubMed]
- Kellenberger, C.A.; Wilson, S.C.; Sales-Lee, J.; Hammond, M.C. RNA-based fluorescent biosensors for live cell imaging of second messengers cyclic di-GMP and cyclic AMP-GMP. J. Am. Chem. Soc. 2013, 135, 4906–4909. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Wilson, S.C.; Hammond, M.C. Next-generation RNA-based fluorescent biosensors enable anaerobic detection of cyclic di-GMP. Nucleic Acids Res. 2016, 44, e139. [Google Scholar] [CrossRef] [PubMed]
- Antoniani, D.; Rossi, E.; Rinaldo, S.; Bocci, P.; Lolicato, M.; Paiardini, A.; Raffaelli, N.; Cutruzzola, F.; Landini, P. The immunosuppressive drug azathioprine inhibits biosynthesis of the bacterial signal molecule cyclic-di-GMP by interfering with intracellular nucleotide pool availability. Appl. Microbiol. Biotechnol. 2013, 97, 7325–7336. [Google Scholar] [CrossRef]
- De, N.; Pirruccello, M.; Krasteva, P.V.; Bae, N.; Raghavan, R.V.; Sondermann, H. Phosphorylation-independent regulation of the diguanylate cyclase WspR. PLoS Biol. 2008, 6, e67. [Google Scholar] [CrossRef]
- Cromie, M.J.; Shi, Y.; Latifi, T.; Groisman, E.A. An RNA sensor for intracellular Mg(2+). Cell 2006, 125, 71–84. [Google Scholar] [CrossRef]
- Groisman, E.A.; Cromie, M.J.; Shi, Y.; Latifi, T. A Mg2+-responding RNA that controls the expression of a Mg2+ transporter. Cold Spring Harb. Symp. Quant. Biol. 2006, 71, 251–258. [Google Scholar] [CrossRef]
- Ramesh, A.; Winkler, W.C. Magnesium-sensing riboswitches in bacteria. RNA Biol. 2010, 7, 77–83. [Google Scholar] [CrossRef]
- Spinelli, S.V.; Pontel, L.B.; Garcia Vescovi, E.; Soncini, F.C. Regulation of magnesium homeostasis in Salmonella: Mg(2+) targets the mgtA transcript for degradation by RNase E. FEMS Microbiol. Lett. 2008, 280, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Butler, E.B.; Xiong, Y.; Wang, J.; Strobel, S.A. Structural basis of cooperative ligand binding by the glycine riboswitch. Chem. Biol. 2011, 18, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Serganov, A.; Patel, D.J. Structural insights into ligand recognition by a sensing domain of the cooperative glycine riboswitch. Mol. Cell. 2010, 40, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Torgerson, C.D.; Hiller, D.A.; Strobel, S.A. The asymmetry and cooperativity of tandem glycine riboswitch aptamers. RNA 2020, 26, 564–580. [Google Scholar] [CrossRef]
- Sherlock, M.; Breaker, R.R. Former Orphan Riboswitches Reveal Unexplored Areas of Bacterial Metabolism, Signaling and Gene Control Processes. RNA 2020, 26, 675–693. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Strobel, S.A. Chemical basis of glycine riboswitch cooperativity. RNA 2008, 14, 25–34. [Google Scholar] [CrossRef]
- Sherman, E.M.; Esquiaqui, J.; Elsayed, G.; Ye, J.D. An energetically beneficial leader-linker interaction abolishes ligand-binding cooperativity in glycine riboswitches. RNA 2012, 18, 496–507. [Google Scholar] [CrossRef]
- Babina, A.M.; Lea, N.E.; Meyer, M.M. In Vivo Behavior of the Tandem Glycine Riboswitch in Bacillus subtilis. mBio 2017, 8, e01602–e01617. [Google Scholar] [CrossRef]
- Sur, C.; Kinney, G.G. Glycine transporter 1 inhibitors and modulation of NMDA receptor-mediated excitatory neurotransmission. Curr. Drug Targets 2007, 8, 643–649. [Google Scholar] [CrossRef]
- Martin, J.E.; Le, M.T.; Bhattarai, N.; Capdevila, D.A.; Shen, J.; Winkler, M.E.; Giedroc, D.P. A Mn-sensing riboswitch activates expression of a Mn2+/Ca2+ ATPase transporter in Streptococcus. Nucleic Acids Res. 2019, 47, 6885–6899. [Google Scholar] [CrossRef]
- Price, I.R.; Gaballa, A.; Ding, F.; Helmann, J.D.; Ke, A. Mn(2+)-sensing mechanisms of yybP-ykoY orphan riboswitches. Mol. Cell 2015, 57, 1110–1123. [Google Scholar] [CrossRef] [Green Version]
- Waters, L.S.; Sandoval, M.; Storz, G. The Escherichia coli MntR miniregulon includes genes encoding a small protein and an efflux pump required for manganese homeostasis. J. Bacteriol. 2011, 193, 5887–5897. [Google Scholar] [CrossRef] [PubMed]
- Dambach, M.; Sandoval, M.; Updegrove, T.B.; Anantharaman, V.; Aravind, L.; Waters, L.S.; Storz, G. The ubiquitous yybP-ykoY riboswitch is a manganese-responsive regulatory element. Mol. Cell. 2015, 57, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Bachas, S.T.; Ferre-D’Amare, A.R. Convergent Use of Heptacoordination for Cation Selectivity by RNA and Protein Metalloregulators. Cell Chem. Biol. 2018, 25, 962–973.e5. [Google Scholar] [CrossRef]
- Juttukonda, L.J.; Skaar, E.P. Manganese homeostasis and utilization in pathogenic bacteria. Mol. Microbiol. 2015, 97, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Hohle, T.H.; O’Brian, M.R. Magnesium-dependent processes are targets of bacterial manganese toxicity. Mol. Microbiol. 2014, 93, 736–747. [Google Scholar] [CrossRef]
- Baker, J.L.; Sudarsan, N.; Weinberg, Z.; Roth, A.; Stockbridge, R.B.; Breaker, R.R. Widespread genetic switches and toxicity resistance proteins for fluoride. Science 2012, 335, 233–235. [Google Scholar] [CrossRef]
- Stockbridge, R.B.; Lim, H.H.; Otten, R.; Williams, C.; Shane, T.; Weinberg, Z.; Miller, C. Fluoride resistance and transport by riboswitch-controlled CLC antiporters. Proc. Natl. Acad. Sci. USA 2012, 109, 15289–15294. [Google Scholar] [CrossRef]
- Ren, A.; Rajashankar, K.R.; Patel, D.J. Fluoride ion encapsulation by Mg2+ ions and phosphates in a fluoride riboswitch. Nature 2012, 486, 85–89. [Google Scholar] [CrossRef]
- Yost, K.G.; VanDemark, P.J. Growth inhibition of Streptococcus mutans and Leuconostoc mesenteroides by sodium fluoride and ionic tin. Appl. Environ. Microbiol. 1978, 35, 920–924. [Google Scholar] [CrossRef]
- Weinberg, Z.; Wang, J.X.; Bogue, J.; Yang, J.; Corbino, K.; Moy, R.H.; Breaker, R.R. Comparative genomics reveals 104 candidate structured RNAs from bacteria, archaea, and their metagenomes. Genome Biol. 2010, 11, R31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regulski, E.E.; Breaker, R.R. In-line probing analysis of riboswitches. Methods Mol. Biol. 2008, 419, 53–67. [Google Scholar] [PubMed]
- Matulef, K.; Howery, A.E.; Tan, L.; Kobertz, W.R.; Du Bois, J.; Maduke, M. Discovery of potent CLC chloride channel inhibitors. ACS Chem. Biol. 2008, 3, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Dutzler, R.; Campbell, E.B.; Cadene, M.; Chait, B.T.; MacKinnon, R. X-ray structure of a ClC chloride channel at 3.0 A reveals the molecular basis of anion selectivity. Nature 2002, 415, 287–294. [Google Scholar] [CrossRef]
- Johnston, N.R.; Strobel, S.A. Principles of fluoride toxicity and the cellular response: A review. Arch. Toxicol. 2020, 94, 1051–1069. [Google Scholar] [CrossRef]
- Hemlata; Tiwari, A. Applications of bioinformatics tools to combat the antibiotic resistance. In Proceedings of the 2015 International Conference on Soft Computing Techniques and Implementations (ICSCTI), Faridabad, India, 8–10 October 2015. [Google Scholar]
- Kopniczky, M.B.; Moore, S.J.; Freemont, P.S. Multilevel Regulation and Translational Switches in Synthetic Biology. IEEE Trans. Biomed. Circuits Syst. 2015, 9, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Penchovsky, R. Methods for creating novel antibacterial agent using chimeric antisense oligonucleotides. WIPO/PCT International Patent Application WIPO/PCT. Available online: https://www.wipo.int/pct/en/ (accessed on 17 August 2022).
- Kaloudas, D.; Pavlova, N.; Penchovsky, R. EBWS: Essential Bioinformatics Web Services for Sequence Analyses. IEEE/ACM Trans. Comput. Biol. Bioinform. 2018, 16, 942–953. [Google Scholar] [CrossRef]
- Penchovsky, R.; Pavlova, N.; Kaloudas, D. RSwitch: A Novel Bioinformatics Database on Riboswitches as Antibacterial Drug Targets. IEEE/ACM Trans. Comput. Biol. Bioinform 2021, 18, 804–808. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human Pathogenic Bacteria | Glycine | PreQ1 | Mn2+ | Fluoride | MoCo RNA | Cyclic-di-GMP I | Mg2+ sensor | Cyclic-di-GMP II | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Acinetobacter baumannii | + | + | + | |||||
| 2 | Bacillus anthracis | + | + | + | + | + | |||
| 3 | Bacillus cereus | + | + | + | + | + | |||
| 4 | Bordetella pertussis | + | + | + | |||||
| 5 | Brucella abortus | + | |||||||
| 6 | Clostridium botulinum | + | + | + | + | + | |||
| 7 | Clostridium difficile | + | + | + | + | + | |||
| 8 | Clostridium perfringens | + | + | + | + | + | + | ||
| 9 | Clostridium tetani | + | + | + | |||||
| 10 | Enterococcus faecalis | + | + | ||||||
| 11 | Enterococcus faecium | + | + | ||||||
| 12 | Enterobacter sp. | + | + | + | |||||
| 13 | Escherichia coli | + | + | + | + | ||||
| 14 | Haemophilus influenzae | + | + | + | |||||
| 15 | Klebsiella pneumoniae | + | + | + | |||||
| 16 | Listeria monocytogenes | + | + | + | |||||
| 17 | Mycobacterium tuberculosis | + | |||||||
| 18 | Mycobacterium ulcerans | + | |||||||
| 19 | Neisseria gonorrhoeae | + | + | ||||||
| 20 | Neisseria meningitidis | + | + | ||||||
| 21 | Pseudomonas aeruginosa | + | + | + | |||||
| 22 | Salmonella enterica | + | + | + | |||||
| 23 | Salmonella typhi | + | + | ||||||
| 24 | Shigella sonnei | + | + | ||||||
| 25 | Staphylococcus aureus | + | + | ||||||
| 26 | Staphylococcus epidermidis | + | + | + | |||||
| 27 | Staphylococcus saprophyticus | + | + | ||||||
| 28 | Streptococcus agalactiae | + | + | ||||||
| 29 | Streptococcus mutans | + | + | ||||||
| 30 | Streptococcus prenumoniae | + | + | ||||||
| 31 | Streptococcus pyogenes | + | |||||||
| 32 | Vibrio cholerae | + | + | + | + | ||||
| 33 | Yersinia pestis | + | + | + | |||||
| Number | 24 | 15 | 14 | 13 | 10 | 7 | 6 | 4 | |
| Riboswitch | Riboswitch-Controlled Biosynthetic Pathway | Transporter Protein for Essential Metabolite | Alternative Biosynthetic Pathway Not under Riboswitch Control | Suitability |
|---|---|---|---|---|
| PreQ1 | ✔ | - | - | +++ |
| MoCo RNA | ✔ | - | - | +++ |
| Cyclic-di-GMP I | ✔ | - | ✔ | ++ |
| Cyclic-di-GMP II | ✔ | - | ✔ | ++ |
| Mg2+ sensor | ✔ | ✔ | ✔/- | ++/+ |
| Glycine | - | - | - | -/+ |
| Mn2+ | - | ✔ | - | -/+ |
| Fluoride | - | ✔ | - | -/+ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlova, N.; Penchovsky, R. Bioinformatics and Genomic Analyses of the Suitability of Eight Riboswitches for Antibacterial Drug Targets. Antibiotics 2022, 11, 1177. https://doi.org/10.3390/antibiotics11091177
Pavlova N, Penchovsky R. Bioinformatics and Genomic Analyses of the Suitability of Eight Riboswitches for Antibacterial Drug Targets. Antibiotics. 2022; 11(9):1177. https://doi.org/10.3390/antibiotics11091177
Chicago/Turabian StylePavlova, Nikolet, and Robert Penchovsky. 2022. "Bioinformatics and Genomic Analyses of the Suitability of Eight Riboswitches for Antibacterial Drug Targets" Antibiotics 11, no. 9: 1177. https://doi.org/10.3390/antibiotics11091177
APA StylePavlova, N., & Penchovsky, R. (2022). Bioinformatics and Genomic Analyses of the Suitability of Eight Riboswitches for Antibacterial Drug Targets. Antibiotics, 11(9), 1177. https://doi.org/10.3390/antibiotics11091177