
Chemical Profile, Anti-Microbial and Anti-Inflammaging Activities of Santolina rosmarinifolia L. Essential Oil from Portugal

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of the Essential Oil
2.2. Antifungal Activity
2.2.1. S. rosmarinifolia Inhibited the Growth of Several Pathogenic Fungi
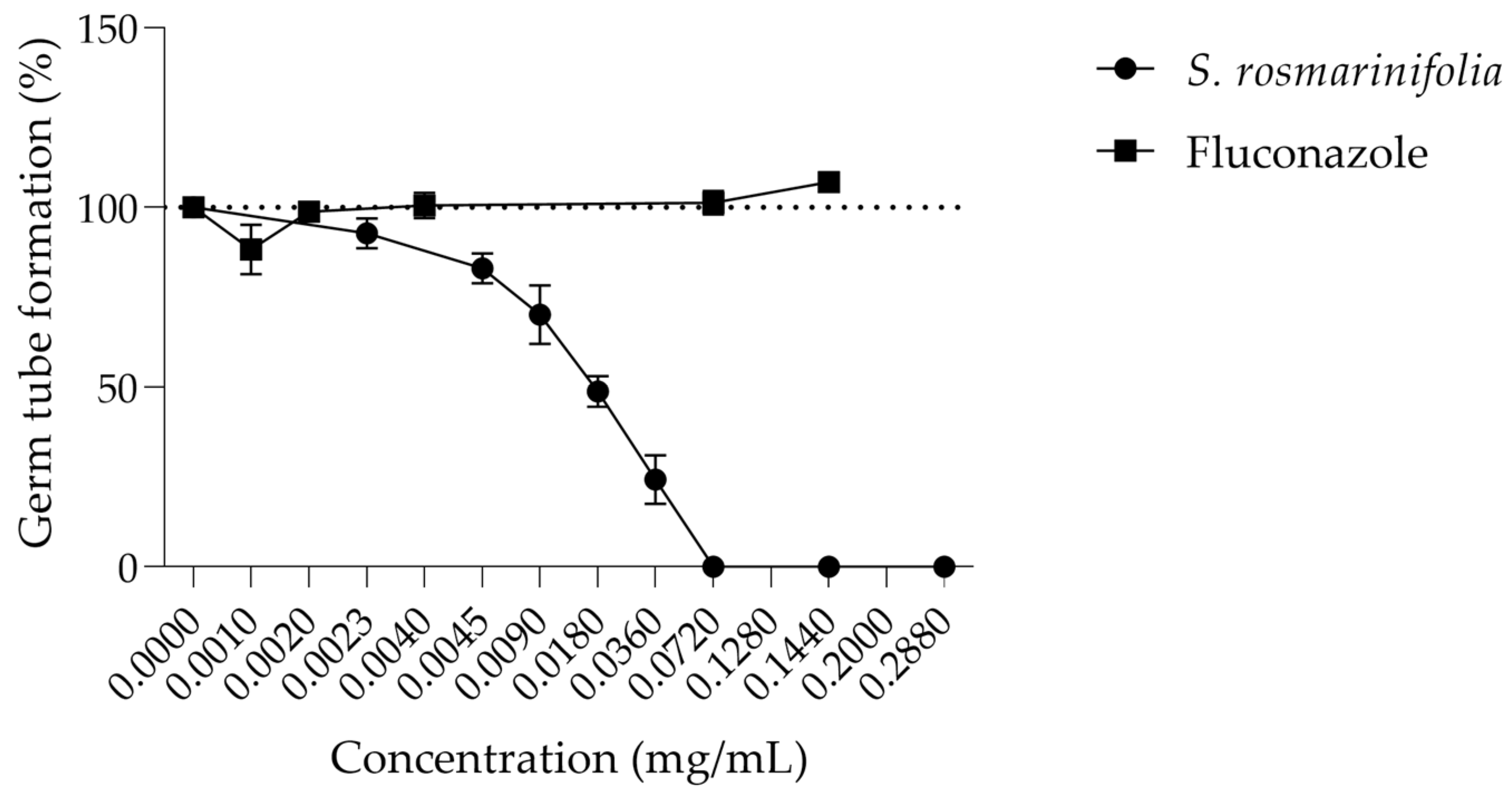
2.2.2. S. rosmarinifolia Essential Oil Inhibited Germ Tube Formation by C. albicans
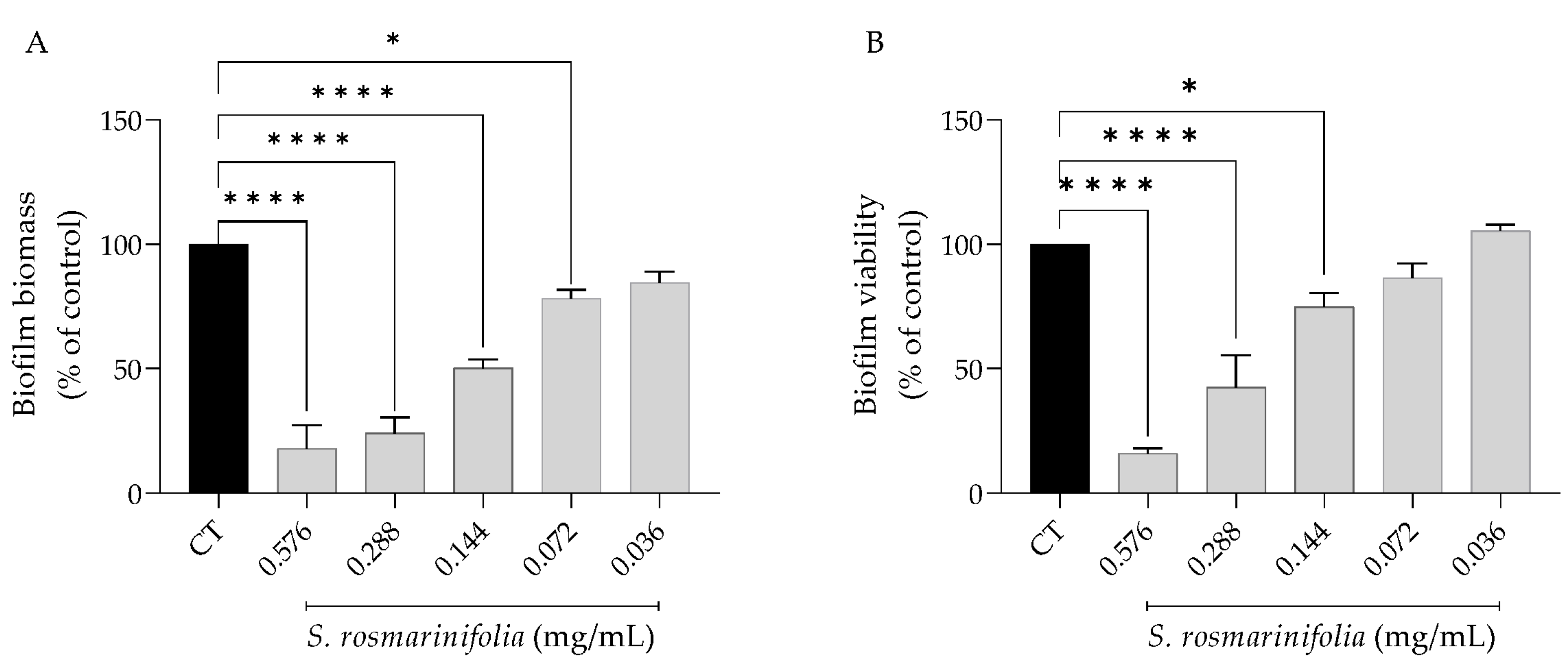
2.2.3. S. rosmarinifolia Inhibited C. albicans Biofilm Formation
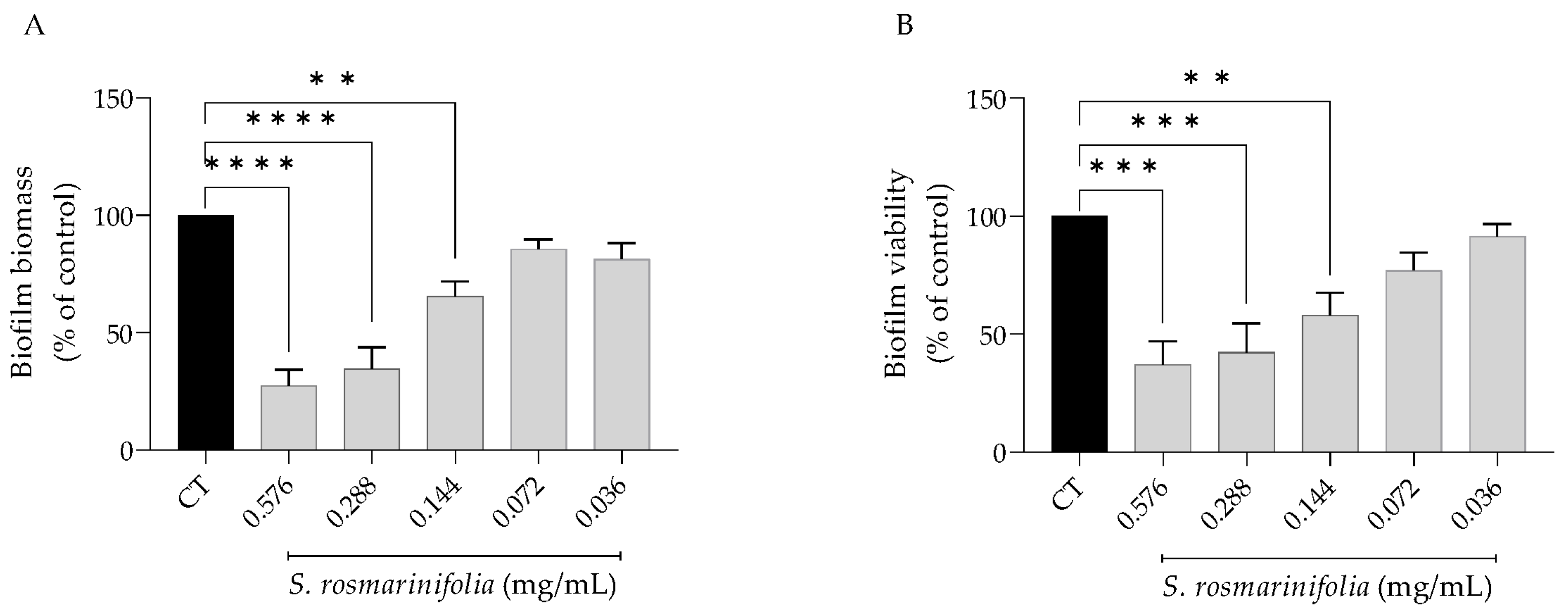
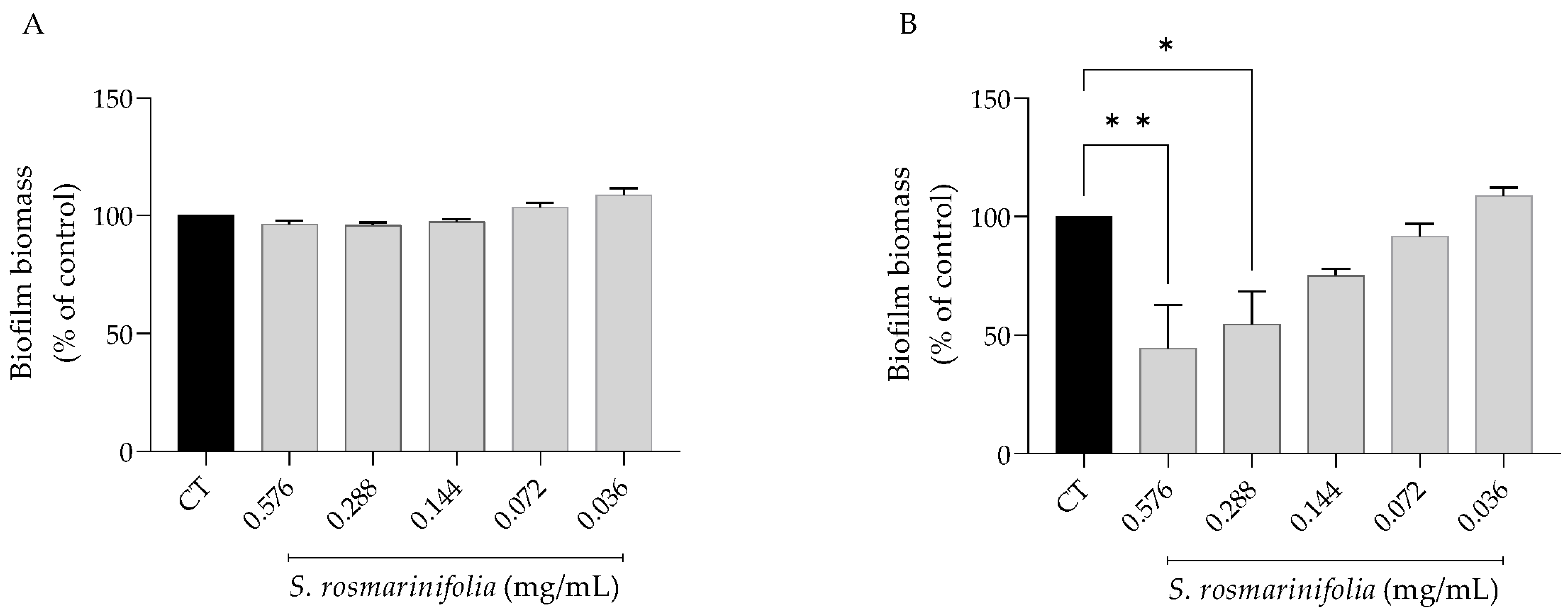
2.2.4. S. rosmarinifolia Disrupted C. albicans Preformed Biofilms
2.3. Anti-Inflammatory Effect
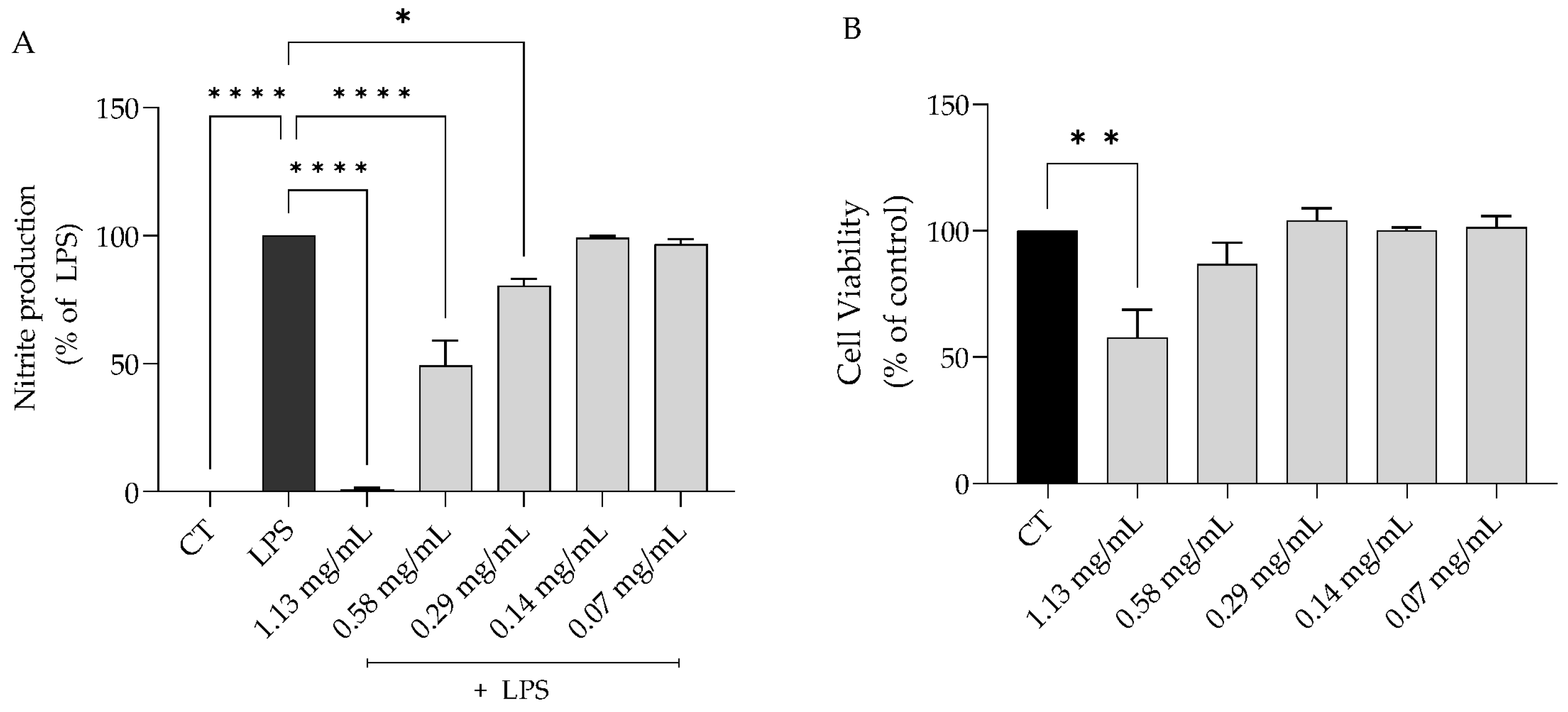
2.3.1. S. rosmarinifolia Inhibited Nitric Oxide Production on LPS-Stimulated Macrophages
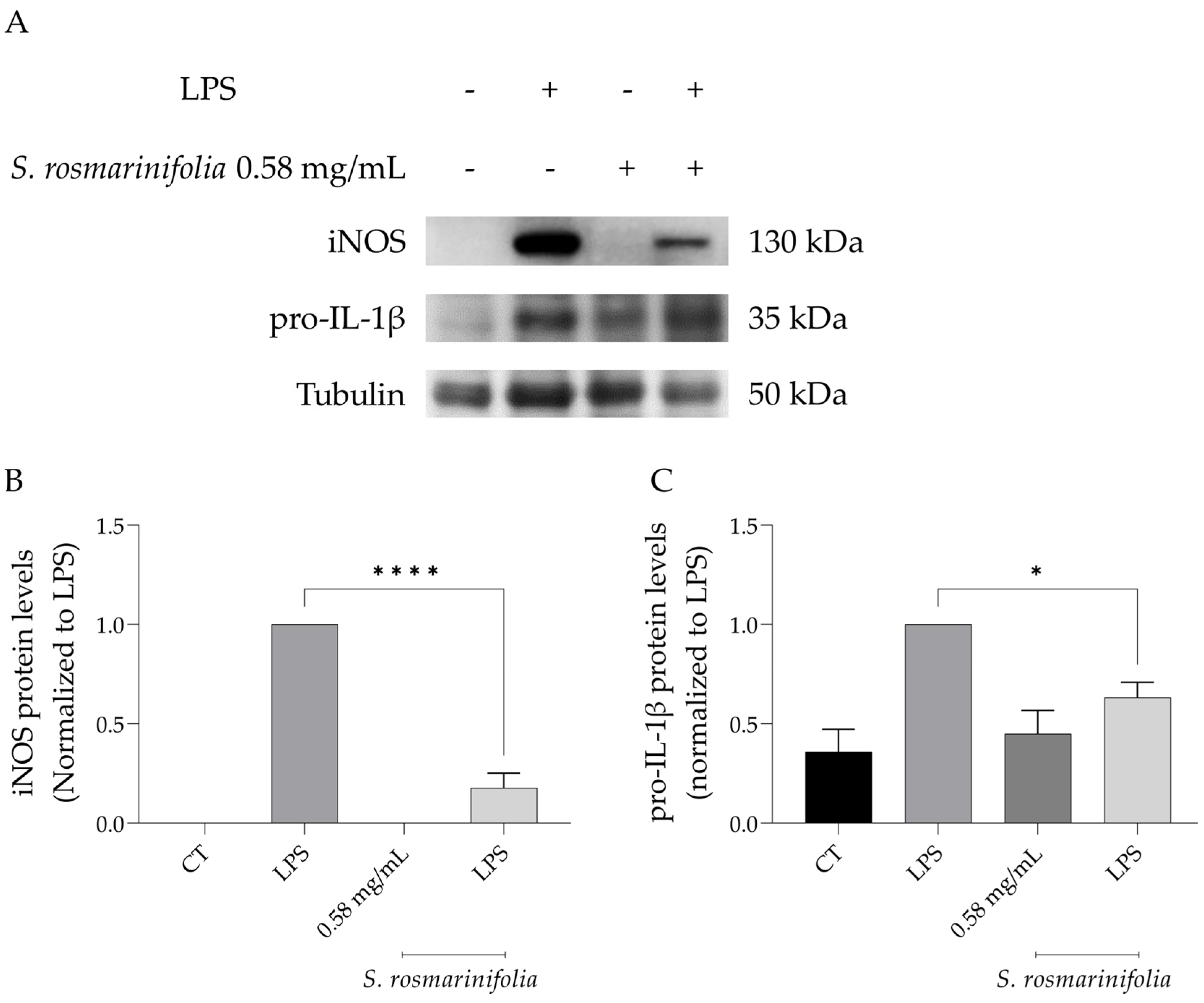
2.3.2. S. rosmarinifolia Inhibited Protein Levels of iNOS and Pro-IL1β
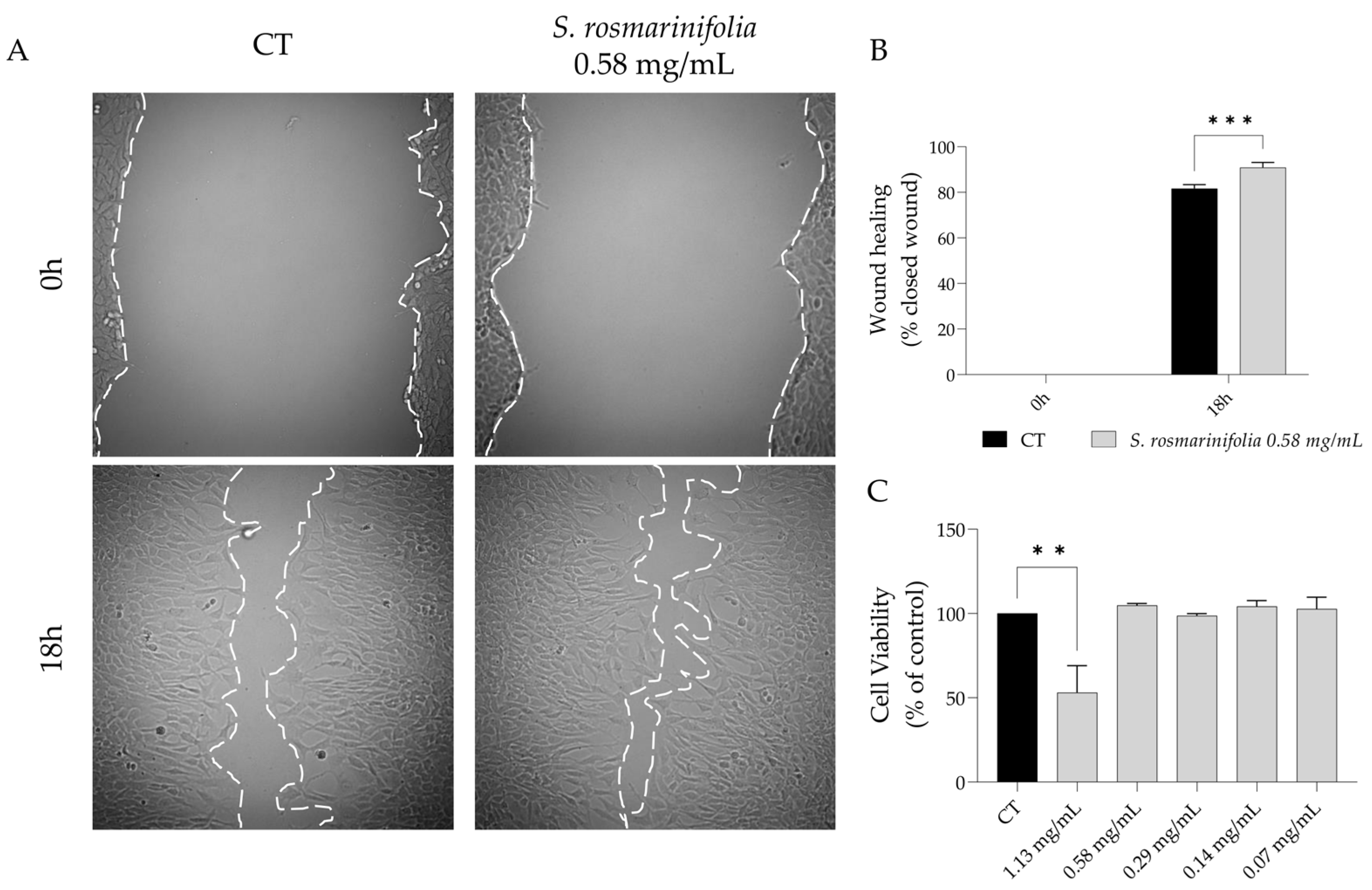
2.4. S. rosmarinifolia Promoted Wound Healing
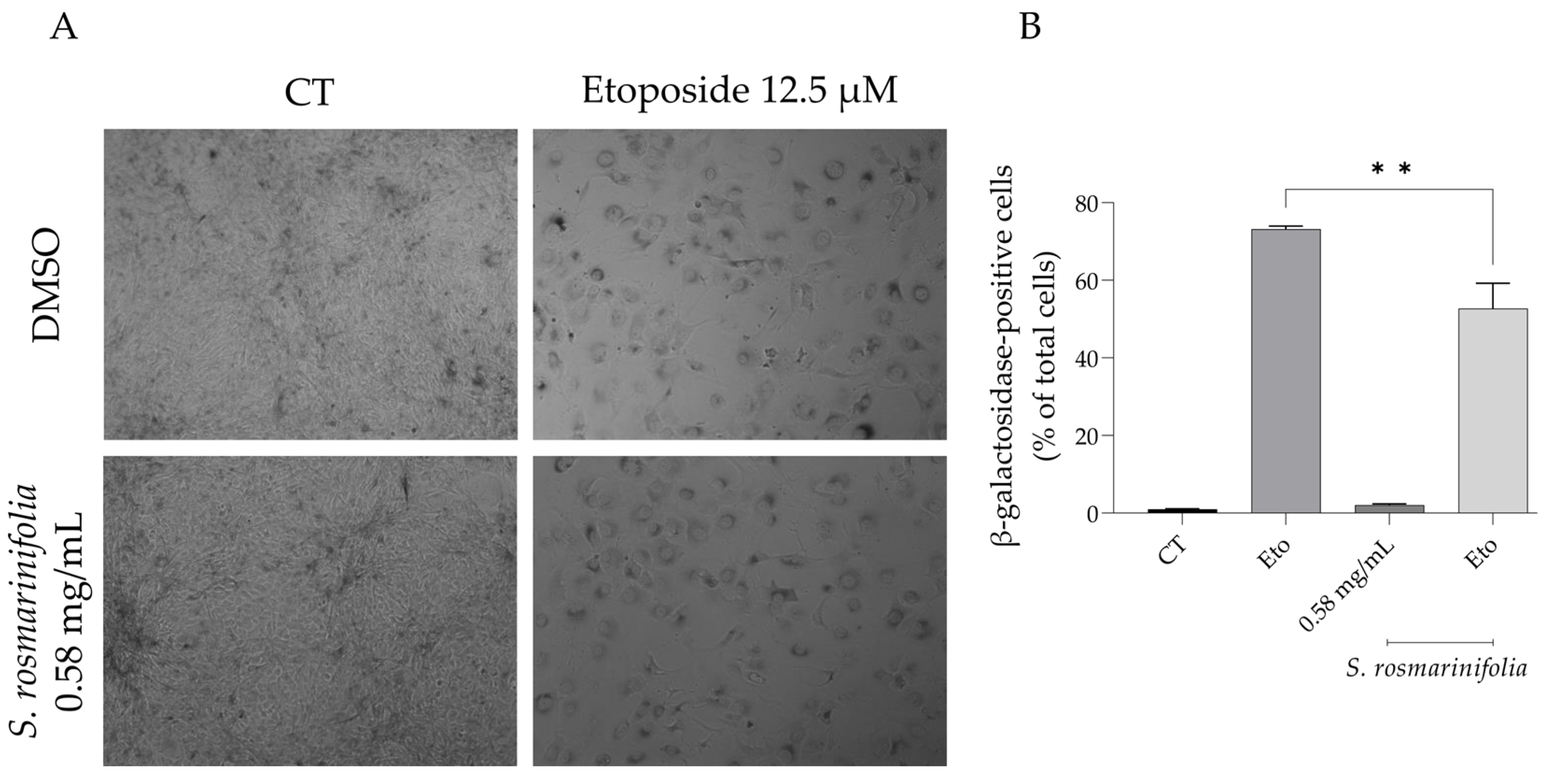
2.5. S. rosmarinifolia Decreased Etoposide-Induced Cell Senescence
3. Discussion
4. Materials and Methods
4.1. Essential Oil Distillation and Analysis
4.1.1. Plant Collection and Essential Oil Distillation
4.1.2. Essential Oil Analysis
4.2. Antifungal Activity
4.2.1. Fungal Strains
4.2.2. Macrodilution Broth Assay
4.2.3. C. albicans Germ Tube Formation
4.2.4. C. albicans Biofilm Formation and Disruption of Preformed Biofilm
4.3. Anti-Inflammatory Assays
4.3.1. Cell Culture
4.3.2. Nitric Oxide Production
4.3.3. Expression of Pro-Inflammatory Proteins, iNOS and pro-IL-1β
4.4. Cell Migration
4.4.1. Cell Culture
4.4.2. Cell Migration Assay
4.5. Cell Viability
4.6. Etoposide-Induced Senescence
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and Multi-National Prevalence of Fungal Dis-eases-Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Pierce, C.; Lopez-Ribot, J.L. Candidiasis Drug Discovery and Development: New Approaches Targeting Vir-ulence for Discovering and Identifying New Drugs. Expert Opin. Drug Discov. 2013, 8, 1117–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campoy, S.; Adrio, J.L. Antifungals. Biochem. Pharm. 2017, 133, 86–96. [Google Scholar] [CrossRef]
- Dadar, M.; Tiwari, R.; Karthik, K.; Chakraborty, S.; Shahali, Y.; Dhama, K. Candida Albicans - Biology, Mo-lecular Characterization, Pathogenicity, and Advances in Diagnosis and Control—An Update. Microb. Pathog. 2018, 117, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Gulati, M.; Nobile, C.J. Candida Albicans Biofilms: Development, Regulation, and Molecular Mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kernien, J.F.; Snarr, B.D.; Sheppard, D.C.; Nett, J.E. The Interface between Fungal Biofilms and Innate Im-munity. Front. Immunol. 2018, 8, 1968. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-H.; Kim, Y.-G.; Gupta, V.K.; Manoharan, R.K.; Lee, J. Suppression of Fluconazole Resistant Candida Albicans Biofilm Formation and Filamentation by Methylindole Derivatives. Front. Microbiol. 2018, 9, 2641. [Google Scholar] [CrossRef]
- Silva, S.; Rodrigues, C.; Araújo, D.; Rodrigues, M.; Henriques, M. Candida Species Biofilms’ Antifungal Resis-tance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.K.; Cooper, E.A. Update in Antifungal Therapy of Dermatophytosis. Mycopathologia 2008, 166, 353–367. [Google Scholar] [CrossRef]
- Piggott, C.D.S.; Friedlander, S.F. Etiologic Agents of Infections Diseases. In Principles and Practice of Pediatric Infections Diseases; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Cardoso, N.N.R.; Alviano, C.S.; Blank, A.F.; Romanos, M.T.V.; Fonseca, B.B.; Rozental, S.; Rodrigues, I.A.; Al-viano, D.S. Synergism Effect of the Essential Oil from Ocimum Basilicum Var. Maria Bonita and Its Major Components with Fluconazole and Its Influence on Ergosterol Biosynthesis. Evid. Based Complement. Altern. Med. 2016, 2016, 5647182. [Google Scholar] [CrossRef]
- Desnos-Ollivier, M.; Letscher-Bru, V.; Neuvéglise, C.; Dromer, F. Yarrowia Lipolytica Causes Sporadic Cases and Local Outbreaks of Infections and Colonisation. Mycoses 2020, 63, 737–745. [Google Scholar] [CrossRef]
- Yan, W.; Zhao, Y.; Xie, K.; Xing, Y.; Xu, F. Aspergillus Fumigatus Influences Gasdermin-D-Dependent Pyrop-tosis of the Lung via Regulating Toll-Like Receptor 2-Mediated Regulatory T Cell Differentiation. J. Immunol. Res. 2021, 2021, 5538612. [Google Scholar] [CrossRef]
- Wang, W.; Deng, Z.; Wu, H.; Zhao, Q.; Li, T.; Zhu, W.; Wang, X.; Tang, L.; Wang, C.; Cui, S.-Z.; et al. A Small Secreted Protein Triggers a TLR2/4-Dependent Inflammatory Response during Invasive Candida Albicans Infection. Nat. Commun. 2019, 10, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celestrino, G.A.; Reis, A.P.C.; Criado, P.R.; Benard, G.; Sousa, M.G.T. Trichophyton Rubrum Elicits Phagocytic and Pro-Inflammatory Responses in Human Monocytes Through Toll-Like Receptor 2. Front. Microbiol. 2019, 10, 2589. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.-C. The Non-Canonical NF-B Pathway in Immunity and Inflammation. Nat. Rev. Immunol. 2017, 17, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Chen, Z.; Shen, W.; Huang, G.; Sedivy, J.M.; Wang, H.; Ju, Z. Inflammation, Epigenetics, and Metabo-lism Converge to Cell Senescence and Ageing: The Regulation and Intervention. Signal Transduct Target 2021, 6, 245. [Google Scholar] [CrossRef]
- Stojanović, S.D.; Fiedler, J.; Bauersachs, J.; Thum, T.; Sedding, D.G. Senescence-Induced Inflammation: An Important Player and Key Therapeutic Target in Atherosclerosis. Eur. Heart J. 2020, 41, 2983–2996. [Google Scholar] [CrossRef] [Green Version]
- Malani, P.N.; McNeil, S.A.; Bradley, S.F.; Kauffman, C.A. Candida Albicans Sternal Wound Infections: A Chronic and Recurrent Complication of Median Sternotomy. Clin. Infect. Dis. 2002, 35, 1316–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, J.; Solis, M.; Higa, A.; Davis, S.C. Candida Albicans Infections: A Novel Porcine Wound Model to Evalu-ate Treatment Efficacy. BMC Microbiol. 2022, 22, 45. [Google Scholar] [CrossRef]
- Kalan, L.; Grice, E.A. Fungi in the Wound Microbiome. Adv. Wound Care 2018, 7, 247–255. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, S. Protective Manifestation of Herbonanoceuticals as Antifungals: A Possible Drug Can-didate for Dermatophytic Infection. Health Sci. Rep. 2022, 5, e775. [Google Scholar] [CrossRef]
- Wu, Y.-K.; Cheng, N.-C.; Cheng, C.-M. Biofilms in Chronic Wounds: Pathogenesis and Diagnosis. Trends Bio-technol. 2019, 37, 505–517. [Google Scholar] [CrossRef]
- Percival, S.L. Importance of Biofilm Formation in Surgical Infection. Br. J. Surg. 2017, 104, e85–e94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; DiPietro, L.A. Factors Affecting Wound Healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef]
- Pfaller, M.A. Antifungal Drug Resistance: Mechanisms, Epidemiology, and Consequences for Treatment. Am. J. Med. 2012, 125, S3–S13. [Google Scholar] [CrossRef] [PubMed]
- Fuentefria, A.M.; Pippi, B.; Dalla Lana, D.F.; Donato, K.K.; de Andrade, S.F. Antifungals Discovery: An Insight into New Strategies to Combat Antifungal Resistance. Lett. Appl. Microbiol. 2018, 66, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Pozzatti, P.; Scheid, L.A.; Spader, T.B.; Atayde, M.L.; Santurio, J.M.; Alves, S.H. In Vitro Activity of Essential Oils Extracted from Plants Used as Spices against Fluconazole-Resistant and Fluconazole-Susceptible Can-dida Spp. Can J. Microbiol. 2008, 54, 950–956. [Google Scholar] [CrossRef]
- Mourad, A.; Perfect, J.R. The War on Cryptococcosis: A Review of the Antifungal Arsenal. Mem. Inst. Oswaldo. Cruz. 2018, 113, e170391. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-Resistant Aspergillosis: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S436–S444. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Rossi, N.M.; Bitencourt, T.A.; Peres, N.T.A.; Lang, E.A.S.; Gomes, E.V.; Quaresemin, N.R.; Martins, M.P.; Lopes, L.; Rossi, A. Dermatophyte Resistance to Antifungal Drugs: Mechanisms and Prospectus. Front. Microbiol. 2018, 9, 1108. [Google Scholar] [CrossRef]
- Colombo, A.L.; Júnior, J.N.; de Almeida Júnior, J.N.; Guinea, J. Emerging Multidrug-Resistant Candida Species. Curr. Opin Infect. Dis. 2017, 30, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Ahmad, A.; Akhtar, F.; Yousuf, S.; Xess, I.; Khan, L.A.; Manzoor, N. Ocimum Sanctum Essential Oil and Its Active Principles Exert Their Antifungal Activity by Disrupting Ergosterol Biosynthesis and Mem-brane Integrity. Res. Microbiol. 2010, 161, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Attiq, A.; Jalil, J.; Husain, K.; Ahmad, W. Raging the War Against Inflammation With Natural Products. Front. Pharm. 2018, 9, 976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, R.J.S.; Xolalpa, N.C.; Castro, A.J.A.; Pérez González, C.; Pérez Ramos, J.; Pérez Gutiérrez, S. Terpenes from Natural Products with Potential Anti-Inflammatory Activity. In Terpenes and Terpenoids; IntechOpen: London, UK, 2018. [Google Scholar]
- Serra, E.; Hidalgo-Bastida, L.; Verran, J.; Williams, D.; Malic, S. Antifungal Activity of Commercial Essential Oils and Biocides against Candida Albicans. Pathogens 2018, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils—A Review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia L. Genus: A Review of Bioactive Essential Oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef] [Green Version]
- Babaei, G.; Aliarab, A.; Abroon, S.; Rasmi, Y.; Aziz, S.G.-G. Application of Sesquiterpene Lactone: A New Promising Way for Cancer Therapy Based on Anticancer Activity. Biomed. Pharmacother. 2018, 106, 239–246. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R. A Review of the Traditional Uses, Phytochemistry and Biological Activities of the Genus Santolina. Planta Med. 2018, 84, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Novais, M.H.; Santos, I.; Mendes, S.; Pinto-Gomes, C. Studies on Pharmaceutical Ethnobotany in Arrabida Natural Park (Portugal). J. Ethnopharmacol. 2004, 93, 183–195. [Google Scholar] [CrossRef]
- Beniston, N.T.; Beniston, W.S. Fleurs d’Algérie; Entreprise Nationale du Livre: Alger, Algérie, 1984. [Google Scholar]
- RIVERO-GUERRA, A.O. Phenotypic Differentiation of Peripheral Populations of Santolina Rosmarinifolia (Asteraceae). Bot. J. Linn. Soc. 2008, 158, 650–668. [Google Scholar] [CrossRef]
- Stappers, M.H.T.; Brown, G.D. Host Immune Responses During Infections with Candida Albicans. In Candida albicans: Cellular and Molecular Biology; Springer International Publishing: Cham, Switzerland, 2017; pp. 145–183. [Google Scholar]
- Guijarro-Muñoz, I.; Compte, M.; Álvarez-Cienfuegos, A.; Álvarez-Vallina, L.; Sanz, L. Lipopolysaccharide Activates Toll-like Receptor 4 (TLR4)-Mediated NF-ΚB Signaling Pathway and Proinflammatory Response in Human Pericytes. J. Biol. Chem. 2014, 289, 2457–2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basak, C.; Pathak, S.K.; Bhattacharyya, A.; Mandal, D.; Pathak, S.; Kundu, M. NF-ΚB- and C/EBPβ-Driven Interleukin-1β Gene Expression and PAK1-Mediated Caspase-1 Activation Play Essential Roles in Interleu-kin-1β Release from Helicobacter Pylori Lipopolysaccharide-Stimulated Macrophages. J. Biol. Chem. 2005, 280, 4279–4288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, A.; Pimpão, R.C.; Fortalezas, S.; Figueira, I.; Miguel, C.; Aguiar, C.; Salgueiro, L.; Cavaleiro, C.; Gon-çalves, M.J.; Clemente, A.; et al. Chemical Characterization and Bioactivity of Phytochemicals from Iberian Endemic Santolina Semidentata and Strategies for Ex Situ Propagation. Ind. Crop. Prod. 2015, 74, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Alves-Silva, J.M.; Zuzarte, M.; Gonçalves, M.J.; Cruz, M.T.; Cavaleiro, C.; Salgueiro, L. Unveiling the Bioac-tive Potential of the Essential Oil of a Portuguese Endemism, Santolina Impressa. J. Ethnopharmacol. 2019, 244, 112–120. [Google Scholar] [CrossRef]
- Palá-Paúl, J.; Pérez-Alonso, M.J.; Velasco-Negueruela, A.; Palá-Paúl, R.; Sanz, J.; Conejero, F. Seasonal Varia-tion in Chemical Constituents of Santolina rosmarinifolia L. Ssp. Rosmarinifolia. Biochem. Syst. Ecol 2001, 29, 663–672. [Google Scholar] [CrossRef]
- Palá-Paúl, J.; Pérez-Alonso, M.J.; Velasco-Negueruela, A.; Ramos-Vázquez, P.; Gómez-Contreras, F.; Sanz, J. Essential Oil OfSantolina Rosmarinifolia L. Ssp.Rosmarinifolia: First Isolation of Capillene, a Diacetylene Derivative. Flavour Fragr. J. 1999, 14, 131–134. [Google Scholar] [CrossRef]
- Mohammadhosseini, M.; Sarker, S.D.; Akbarzadeh, A. Chemical Composition of the Essential Oils and Ex-tracts of Achillea Species and Their Biological Activities: A Review. J. Ethnopharmacol. 2017, 199, 257–315. [Google Scholar] [CrossRef]
- Raut, J.S.; Karuppayil, S.M. A Status Review on the Medicinal Properties of Essential Oils. Ind. Crop. Prod. 2014, 62, 250–264. [Google Scholar] [CrossRef]
- Tabanca, N.; Demirci, B.; Gürbüz, İ.; Demirci, F.; Becnel, J.J.; Wedge, D.E.; Can Başer, K.H. Essential Oil Com-position of Five Collections of Achillea Biebersteinii from Central Turkey and Their Antifungal and Insecti-cidal Activity. Nat. Prod. Commun. 2011, 6, 1934578X1100600526. [Google Scholar]
- Chibani, S.; Amira, L.; Kabouche, A.; Semra, Z.; Smati, F.; Aburjai, T.; Kabouche, Z. Antibacterial Activity and Chemical Composition of Essential Oil of Santolina Rosmarinifolia L. (Asteraceae) from Algeria. Pharm. Lett. 2013, 5, 238–241. [Google Scholar]
- Ioannou, E.; Poiata, A.; Hancianu, M.; Tzakou, O. Chemical Composition and in Vitro Antimicrobial Activity of the Essential Oils of Flower Heads and Leaves of Santolina rosmarinifolia L. Form. Romania. Nat. Prod. Res. 2007, 21, 18–23. [Google Scholar] [CrossRef] [PubMed]
- DJAMEL, S.; HENDEL, N.; HADJER, F.; GİUSEPPE, R.; MADANİ, S. Chemical Constituents of the Aerial Parts of Santolina Rosmarinifolia L. a Wild Algerian Medicinal Plant. Nat. Volatiles Essent. Oils 2021, 8, 22–28. [Google Scholar] [CrossRef]
- Baptiste Hzounda Fokou, J.; Michel Jazet Dongmo, P.; Fekam Boyom, F. Essential Oil’s Chemical Composition and Pharmacological Properties. In Essential Oils—Oils of Nature; IntechOpen: London, UK, 2020. [Google Scholar]
- da Silva, A.C.R.; Lopes, P.M.; de Azevedo, M.M.B.; Costa, D.C.M.; Alviano, C.S.; Alviano, D.S. Biological Ac-tivities of A-Pinene and -Pinene Enantiomers. Molecules 2012, 17, 6305–6316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.-K.; Lee, S.-Y.; Kim, S.-H.; Hong, C.-Y.; Park, M.-J.; Choi, I.-G. Antifungal Activities of Essential Oils from Six Conifers against Aspergillus Fumigatus. J. Korean Wood Sci. Technol. 2012, 40, 133–140. [Google Scholar] [CrossRef] [Green Version]
- de Macêdo Andrade, A.C.; Rosalen, P.L.; Freires, I.A.; Scotti, L.; Scotti, M.T.; Aquino, S.G.; de Castro, R.D. An-tifungal Activity, Mode of Action, Docking Prediction and Anti-Biofilm Effects of (+)-β-Pinene Enantiomers against Candida Spp. Curr. Top. Med. Chem. 2018, 18, 2481–2490. [Google Scholar] [CrossRef] [PubMed]
- Shin, S. Antifungal Activities of Essential Oils from Glehnia Littoralis Alone and in Combination with Keto-conazole. Nat. Prod. Sci. 2005, 11, 92–96. [Google Scholar]
- Iraji, A.; Yazdanpanah, S.; Alizadeh, F.; Mirzamohammadi, S.; Ghasemi, Y.; Pakshir, K.; Yang, Y.; Zomorodi-an, K. Screening the Antifungal Activities of Monoterpenes and Their Isomers against Candida Species. J. Appl. Microbiol. 2020, 129, 1541–1551. [Google Scholar] [CrossRef]
- Yu, H.; Lin, Z.-X.; Xiang, W.-L.; Huang, M.; Tang, J.; Lu, Y.; Zhao, Q.-H.; Zhang, Q.; Rao, Y.; Liu, L. Antifungal Activity and Mechanism of D-Limonene against Foodborne Opportunistic Pathogen Candida Tropicalis. LWT 2022, 159, 113144. [Google Scholar] [CrossRef]
- Cai, R.; Hu, M.; Zhang, Y.; Niu, C.; Yue, T.; Yuan, Y.; Wang, Z. Antifungal Activity and Mechanism of Citral, Limonene and Eugenol against Zygosaccharomyces Rouxii. LWT 2019, 106, 50–56. [Google Scholar] [CrossRef]
- Chee, H.Y.; Kim, H.; Lee, M.H. In Vitro Antifungal Activity of Limonene against Trichophyton Rubrum. Myco-Biol. 2009, 37, 243. [Google Scholar] [CrossRef] [Green Version]
- Thakre, A.; Zore, G.; Kodgire, S.; Kazi, R.; Mulange, S.; Patil, R.; Shelar, A.; Santhakumari, B.; Kulkarni, M.; Kharat, K.; et al. Limonene Inhibits Candida Albicans Growth by Inducing Apoptosis. Med. Mycol. 2018, 56, 565–578. [Google Scholar] [CrossRef] [PubMed]
- ÜNAL, M.Ü.; UÇAN, F.; ŞENER, A.; DİNÇER, S. Research on Antifungal and Inhibitory Effects of DL-Limonene on Some Yeasts. Turk. J. Agric. For. 2012, 36, 576–582. [Google Scholar] [CrossRef]
- Muñoz, J.E.; Rossi, D.C.P.; Jabes, D.L.; Barbosa, D.A.; Cunha, F.F.M.; Nunes, L.R.; Arruda, D.C.; Pelleschi Taborda, C. In Vitro and In Vivo Inhibitory Activity of Limonene against Different Isolates of Candida Spp. J. Fungi 2020, 6, 183. [Google Scholar] [CrossRef] [PubMed]
- Leite-Andrade, M.C.; de Araújo Neto, L.N.; Buonafina-Paz, M.D.S.; de Assis Graciano dos Santos, F.; da Silva Alves, A.I.; de Castro, M.C.A.B.; Mori, E.; de Lacerda, B.C.G.V.; Araújo, I.M.; Coutinho, H.D.M.; et al. Anti-fungal Effect and Inhibition of the Virulence Mechanism of D-Limonene against Candida Parapsilosis. Molecules 2022, 27, 8884. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, S.H.; Hong, C.Y.; Park, M.-J.; Choi, I.-G. Effects of (−)-Borneol on the Growth and Morphology of Aspergillus Fumigatus and Epidermophyton Floccosom. Flavour Fragr. J. 2013, 28, 129–134. [Google Scholar] [CrossRef]
- Cheng, S.-S.; Lin, C.-Y.; Gu, H.-J.; Chang, S.-T. Antifungal Activities and Chemical Composition of Wood and Leaf Essential Oils from Cunninghamia Konishii. J. Wood Chem. Technol. 2011, 31, 204–217. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.; Zhan, F. Effects of Natural Borneol on Germ Tube Formation and Preformed Biofilm Ac-tivity in Candida Albicans. Nat. Prod. Commun. 2022, 17, 1934578X2211291. [Google Scholar] [CrossRef]
- Roxo, M.; Zuzarte, M.; Gonçalves, M.J.; Alves-Silva, J.M.; Cavaleiro, C.; Cruz, M.T.; Salgueiro, L. Antifungal and Anti-Inflammatory Potential of the Endangered Aromatic Plant Thymus Albicans. Sci. Rep. 2020, 10, 18859. [Google Scholar] [CrossRef]
- CHANG, H.; CHENG, Y.; WU, C.; CHANG, S.; CHANG, T.; SU, Y. Antifungal Activity of Essential Oil and Its Constituents from Calocedrus Macrolepis Var. Formosana Florin Leaf against Plant Pathogenic Fungi. Biore-Sour Technol. 2008, 99, 6266–6270. [Google Scholar] [CrossRef]
- Jain, N.; Sharma, M. Inhibitory Effect of Some Selected Essential Oil Terpenes on Fungi Causing Superficial Infection in Human Beings. J. Essent. Oil Bear. Plants 2020, 23, 862–869. [Google Scholar] [CrossRef]
- Tavares, A.C.; Gonçalves, M.J.; Cruz, M.T.; Cavaleiro, C.; Lopes, M.C.; Canhoto, J.; Salgueiro, L.R. Essential Oils from Distichoselinum Tenuifolium: Chemical Composition, Cytotoxicity, Antifungal and An-ti-Inflammatory Properties. J. Ethnopharmacol. 2010, 130, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Betancur-Galvis, L.; Zapata, B.; Baena, A.; Bueno, J.; Ruíz-Nova, C.A.; Stashenko, E.; Mesa-Arango, A.C. An-tifungal, Cytotoxic and Chemical Analyses of Essential Oils of Lippia Origanoides H.B.K Grown in Colombia. Revista de la Universidad Industrial de Santander. Salud 2011, 43, 141–148. [Google Scholar]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Antifungal Activity of the Components of Melaleuca Alternifolia (Tea Tree) Oil. J. Appl. Microbiol. 2003, 95, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Huo, H.; Gu, Y.; Cao, Y.; Liu, N.; Jia, P.; Kong, W. Antifungal Activity of Camphor against Four Phytopatho-gens of Fusarium. S. Afr. J. Bot. 2021, 148, 437–445. [Google Scholar] [CrossRef]
- Wu, K.; Lin, Y.; Chai, X.; Duan, X.; Zhao, X.; Chun, C. Mechanisms of Vapor-phase Antibacterial Action of Essential Oil from Cinnamomum Camphora Var. Linaloofera Fujita against Escherichia Coli. Food Sci. Nutr. 2019, 7, 2546–2555. [Google Scholar] [CrossRef] [Green Version]
- Magiatis, P.; Skaltsounis, A.-L.; Chinou, I.; Haroutounian, S.A. Chemical Composition and In-Vitro Antimicrobial Activity of the Essential Oils of Three Greek Achillea Species. Z. Für Nat. C 2002, 57, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Alves, M.; Gonçalves, M.J.; Zuzarte, M.; Alves-Silva, J.M.; Cavaleiro, C.; Cruz, M.T.; Salgueiro, L. Unveiling the Antifungal Potential of Two Iberian Thyme Essential Oils: Effect on C. Albicans Germ Tube and Pre-formed Biofilms. Front. Pharm. 2019, 10, 446. [Google Scholar] [CrossRef]
- Manoharan, R.K.; Lee, J.-H.; Lee, J. Antibiofilm and Antihyphal Activities of Cedar Leaf Essential Oil, Cam-phor, and Fenchone Derivatives against Candida Albicans. Front. Microbiol. 2017, 8, 1476. [Google Scholar] [CrossRef] [Green Version]
- Thakre, A.D.; Mulange, S.V.; Kodgire, S.S.; Zore, G.B.; Karuppayil, S.M. Effects of Cinnamaldehyde, Ocimene, Camphene, Curcumin and Farnesene on Candida albicans. Adv. Microbiol. 2016, 6, 627–643. [Google Scholar] [CrossRef] [Green Version]
- Mulyaningsih, S.; Youns, M.; El-Readi, M.Z.; Ashour, M.L.; Nibret, E.; Sporer, F.; Herrmann, F.; Reichling, J.; Wink, M. Biological Activity of the Essential Oil of Kadsura Longipedunculata (Schisandraceae) and Its Major Components. J. Pharm. Pharmacol. 2010, 62, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Marei, G.I.K.; Abdel Rasoul, M.A.; Abdelgaleil, S.A.M. Comparative Antifungal Activities and Biochemical Effects of Monoterpenes on Plant Pathogenic Fungi. Pestic Biochem. Physiol. 2012, 103, 56–61. [Google Scholar] [CrossRef]
- Rajput, S.B.; Karuppayil, S.M. Small Molecules Inhibit Growth, Viability and Ergosterol Biosynthesis in Can-dida Albicans. Springerplus 2013, 2, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raut, J.S.; Shinde, R.B.; Chauhan, N.M.; Karuppayil, S.M. Terpenoids of Plant Origin Inhibit Morphogenesis, Adhesion, and Biofilm Formation ByiCandida Albicans/i. Biofouling 2013, 29, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, M.; Kannan, A.; Stojković, D.S.; Glamočlija, J.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sanglard, D.; Soković, M. Camphor and Eucalyptol—Anticandidal Spectrum, Antivirulence Effect, Efflux Pumps Interference and Cytotoxicity. Int. J. Mol. Sci. 2021, 22, 483. [Google Scholar] [CrossRef]
- Malti, C.E.W.; Baccati, C.; Mariani, M.; Hassani, F.; Babali, B.; Atik-Bekkara, F.; Paoli, M.; Maury, J.; Tomi, F.; Bekhechi, C. Biological Activities and Chemical Composition of Santolina Africana Jord. et Fourr. Aerial Part Essential Oil from Algeria: Occurrence of Polyacetylene Derivatives. Molecules 2019, 24, 204. [Google Scholar] [CrossRef] [Green Version]
- Foddai, M.; Marchetti, M.; Ruggero, A.; Juliano, C.; Usai, M. Evaluation of Chemical Composition and An-ti-Inflammatory, Antioxidant, Antibacterial Activity of Essential Oil of Sardinian Santolina Corsica Jord. & Fourr. Saudi. J. Biol. Sci. 2019, 26, 930–937. [Google Scholar] [CrossRef]
- Alves-Silva, J.M.; Piras, A.; Porcedda, S.; Falconieri, D.; Maxia, A.; Gonçalves, M.J.; Cruz, M.T.; Salgueiro, L. Chemical Characterization and Bioactivity of the Essential Oil from Santolina Insularis, a Sardinian Ende-mism. Nat. Prod. Res. 2021, 36, 445–449. [Google Scholar] [CrossRef]
- Coté, H.; Boucher, M.-A.; Pichette, A.; Legault, J. Anti-Inflammatory, Antioxidant, Antibiotic, and Cytotoxic Activities of Tanacetum vulgare L. Essential Oil and Its Constituents. Medicines 2017, 4, 34. [Google Scholar] [CrossRef]
- Rufino, A.T.; Ribeiro, M.; Judas, F.; Salgueiro, L.; Lopes, M.C.; Cavaleiro, C.; Mendes, A.F. Anti-Inflammatory and Chondroprotective Activity of ()--Pinene: Structural and Enantiomeric Selectivity. J. Nat. Prod. 2014, 77, 264–269. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Kushnarenko, S.V.; Özek, G.; Kirpotina, L.N.; Utegenova, G.A.; Kotukhov, Y.A.; Danilova, A.N.; Özek, T.; Başer, K.H.C.; Quinn, M.T. Inhibition of Human Neutrophil Responses by the Essential Oil of Artemisia Kotuchovii and Its Constituents. J. Agric. Food Chem. 2015, 63, 4999–5007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juhás, Š.; Čikoš, Š.; Czikková, S.; Veselá, J.; Il’Ková, G.; Hájek, T.; Domaracká, K.; Domaracký, M.; Bujňáková, D.; Rehák, P.; et al. Effects of Borneol and Thymoquinone on TNBS-Induced Colitis in Mice. Folia. Biol. 2008, 54, 1–7. [Google Scholar] [PubMed]
- Zou, L.; Zhang, Y.; Li, W.; Zhang, J.; Wang, D.; Fu, J.; Wang, P. Comparison of Chemical Profiles, An-ti-Inflammatory Activity, and UPLC-Q-TOF/MS-Based Metabolomics in Endotoxic Fever Rats between Syn-thetic Borneol and Natural Borneol. Molecules 2017, 22, 1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- do Nascimento-Júnior, B.J.; Brito, L.; de Souza Brito, L.; Barros, W.N.; Gonçalves, D.M.; de Souza Matos, L.; Nascimento, C.R.B.; de Araújo Ribeiro, L.A.; de Lima, R.S.; Silva-Júnior, R.G.C.; et al. Anti-Inflammatory and Healing Action of Oral Gel Containing Borneol Monoterpene in Chemotherapy-Induced Mucositis in Rats (Rattus Norvegicus). Braz. J. Pharm. Sci. 2017, 53. [Google Scholar] [CrossRef]
- Ji, J.; Zhang, R.; Li, H.; Zhu, J.; Pan, Y.; Guo, Q. Analgesic and Anti-Inflammatory Effects and Mechanism of Action of Borneol on Photodynamic Therapy of Acne. Environ. Toxicol. Pharm. 2020, 75, 103329. [Google Scholar] [CrossRef]
- da Silva Almeida, J.R.G.; Souza, G.R.; Silva, J.C.; de Lima Saraiva, S.R.G.; de Oliveira Júnior, R.G.; de Souza Siqueira Quintans, J.; de Souza Siqueira Barreto, R.; Bonjardim, L.R.; de Holanda Cavalcanti, S.C.; Junior, L.J.Q. Borneol, a Bicyclic Monoterpene Al-cohol, Reduces Nociceptive Behavior and Inflammatory Response in Mice. Sci. World J. 2013, 2013, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Rufino, A.T.; Ribeiro, M.; Sousa, C.; Judas, F.; Salgueiro, L.; Cavaleiro, C.; Mendes, A.F. Evaluation of the An-ti-Inflammatory, Anti-Catabolic and pro-Anabolic Effects of E-Caryophyllene, Myrcene and Limonene in a Cell Model of Osteoarthritis. Eur. J. Pharm. 2015, 750, 141–150. [Google Scholar] [CrossRef]
- Souza, M.C.; Siani, A.C.; Ramos, M.F.S.; Menezes-de-Lima, O.J.; Henriques, M.G.M.O. Evaluation of An-ti-Inflammatory Activity of Essential Oils from Two Asteraceae Species. Die Pharm. -Int. J. Pharm. Sci. 2003, 58, 582–586. [Google Scholar]
- Liu, R.; Zhang, L.; Lan, X.; Li, L.; Zhang, T.-T.; Sun, J.-H.; Du, G.-H. Protection by Borneol on Cortical Neurons against Oxygen-Glucose Deprivation/Reperfusion: Involvement of Anti-Oxidation and Anti-Inflammation through Nuclear Transcription Factor ΚappaB Signaling Pathway. Neuroscience 2011, 176, 408–419. [Google Scholar] [CrossRef]
- Rehman, M.U.; Tahir, M.; Khan, A.Q.; Khan, R.; Oday-O-Hamiza; Lateef, A.; Hassan, S.K.; Rashid, S.; Ali, N.; Zeeshan, M.; et al. d-Limonene Suppresses Doxorubicin-Induced Oxidative Stress and Inflam-mation via Repression of COX-2, INOS, and NFκB in Kidneys of Wistar Rats. Exp. Biol. Med. 2014, 239, 465–476. [Google Scholar] [CrossRef]
- Yoon, W.-J.; Lee, N.H.; Hyun, C.-G. Limonene Suppresses Lipopolysaccharide-Induced Production of Nitric Oxide, Prostaglandin E2, and Pro-Inflammatory Cytokines in RAW 264.7 Macrophages. J. Oleo. Sci. 2010, 59, 415–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, N.; Chandra, H. Suppression of Inflammatory and Infection Responses in Lung Macrophages by Eu-calyptus Oil and Its Constituent 1,8-Cineole: Role of Pattern Recognition Receptors TREM-1 and NLRP3, the MAP Kinase Regulator MKP-1, and NFκB. PLoS ONE 2017, 12, e0188232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AM, B.; P, Z.; Filipova M, D.; J, L. Effects of 1,8-Cineole on the Activity of Cyclooxygenase and Cyclooxygen-ase 1 and Cyclooxygenase 2 Isoforms. Nat. Prod. Chem. Res. 2017, 5. [Google Scholar] [CrossRef]
- Bastos, V.P.D.; Gomes, A.S.; Lima, F.J.B.; Brito, T.S.; Soares, P.M.G.; Pinho, J.P.M.; Silva, C.S.; Santos, A.A.; Souza, M.H.L.P.; Magalhães, P.J.C. Inhaled 1,8-Cineole Reduces Inflammatory Parameters in Airways of Ovalbumin-Challenged Guinea Pigs. Basic. Clin. Pharm. Toxicol. 2011, 108, 34–39. [Google Scholar] [CrossRef]
- Juergens, L.J.; Racké, K.; Tuleta, I.; Stoeber, M.; Juergens, U.R. Anti-Inflammatory Effects of 1,8-Cineole (Eu-calyptol) Improve Glucocorticoid Effects in Vitro: A Novel Approach of Steroid-Sparing Add-on Therapy for COPD and Asthma? Synergy 2017, 5, 1–8. [Google Scholar] [CrossRef]
- Santos, F.A.; Rao, V.S.N. Antiinflammatory and Antinociceptive Effects of 1,8-Cineole a Terpenoid Oxide Present in Many Plant Essential Oils. Phytother. Res. 2000, 14, 240–244. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Mohammed, S.A.A.; Khan, O.; Ali, H.M. Topical Eucalyptol Ointment Accelerates Wound Healing and Exerts Antioxidant and Anti-Inflammatory Effects in Rats’ Skin Burn Model. J. Oleo. Sci. 2022, 71, ess22214. [Google Scholar] [CrossRef]
- dos Santos, E.; Leitão, M.M.; Aguero Ito, C.N.; Silva-Filho, S.E.; Arena, A.C.; Silva-Comar, F.M.; de SouzaSilva-Comare, F.M.; Naka-mura Cuman, R.K.; Oliveira, R.J.; Nazari Formagio, A.S.; et al. Analgesic and An-ti-Inflammatory Articular Effects of Essential Oil and Camphor Isolated from Ocimum Kilimandscharicum Gürke Leaves. J. Ethnopharmacol. 2021, 269, 113697. [Google Scholar] [CrossRef]
- Adhikari, A.; Bhandari, S.; Pandey, D.P. Anti-Inflammatory Compounds Camphor and Methylsalicylate from Traditionally Used Pain Curing Plant Equisetum arvense L. J. Nepal. Chem. Soc. 2019, 40, 1–4. [Google Scholar] [CrossRef]
- Silva-Filho, S.; de Souza Silva-Comar, F.; Wiirzler, L.; do Pinho, R.; Grespan, R.; Bersani-Amado, C.; Cuman, R. Effect of Camphor on the Behavior of Leukocytes In Vitro and In Vivo in Acute Inflammatory Response. Trop. J. Pharm. Res. 2015, 13, 2031. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, M.; Kakkar, P. Plant Derived Antioxidants – Geraniol and Camphene Protect Rat Alveolar Macro-phages against t-BHP Induced Oxidative Stress. Toxicol. Vitr. 2009, 23, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Zuzarte, M.; Alves-Silva, J.M.; Alves, M.; Cavaleiro, C.; Salgueiro, L.; Cruz, M.T. New Insights on the An-ti-Inflammatory Potential and Safety Profile of Thymus Carnosus and Thymus Camphoratus Essential Oils and Their Main Compounds. J. Ethnopharmacol. 2018, 225, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Serena, T.; Parnall, L.K.S.; Knox, C.; Vargo, J.; Oliver, A.; Merry, S.; Klugh, A.; Bubar, N.; Anderson, N.; Rie-man, L.; et al. Bismuth Subgallate/Borneol (Suile) Is Superior to Bacitracin in the Human Forearm Biopsy Model for Acute Wound Healing. Adv. Ski. Wound Care 2007, 20, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Barreto, R.S.; Quintans, J.S.; Barreto, A.S.; Albuquerque-Júnior, R.L.; Galvão, J.G.; Gonsalves, J.K.; Nunes, R.S.; Camargo, E.A.; Lucca-Júnior, W.; Soares, R.C.; et al. Improvement of Wound Tissue Repair by Chitosan Films Containing (-)-Borneol, a Bicyclic Monoterpene Alcohol, in Rats. Int. Wound J. 2016, 13, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Keskin, I.; Gunal, Y.; Ayla, S.; Kolbasi, B.; Sakul, A.; Kilic, U.; Gok, O.; Koroglu, K.; Ozbek, H. Effects of Foenic-ulum Vulgare Essential Oil Compounds, Fenchone and Limonene, on Experimental Wound Healing. Biotech. Histochem. 2017, 92, 274–282. [Google Scholar] [CrossRef]
- d’Alessio, P.; Mirshahi, M.; Bisson, J.-F.; Bene, M. Skin Repair Properties of D-Limonene and Perillyl Alcohol in Murine Models. Antiinfla. Antiallergy Agents Med. Chem. 2014, 13, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Khan, T.H.; Ansari, M.N.; Ahmad, S.F. Enhanced Wound Healing by Topical Administration of D-Limonene in Alloxan Induced Diabetic Mice through Reduction of pro-Inflammatory Markers and Chemokine Expression. BMC Genom. 2014, 15, P29. [Google Scholar] [CrossRef] [Green Version]
- Chabane, S.; Boudjelal, A.; Napoli, E.; Benkhaled, A.; Ruberto, G. Phytochemical Composition, Antioxidant and Wound Healing Activities of Teucrium Polium Subsp. Capitatum (L.) Briq. Essent. Oil. J. Essent. Oil Res. 2021, 33, 143–151. [Google Scholar] [CrossRef]
- Wu, Z.; Li, Z.; Liang, Y. Myrcene Exerts Anti-Tumor Effects on Oral Cancer Cells in Vitro via Induction of Apoptosis. Trop. J. Pharm. Res. 2022, 21, 933–938. [Google Scholar] [CrossRef]
- Xanthis, V.; Fitsiou, E.; Voulgaridou, G.-P.; Bogadakis, A.; Chlichlia, K.; Galanis, A.; Pappa, A. Antioxidant and Cytoprotective Potential of the Essential Oil Pistacia Lentiscus Var. Chia and Its Major Components Myrcene and α-Pinene. Antioxidants 2021, 10, 127. [Google Scholar] [CrossRef]
- Salas-Oropeza, J.; Jimenez-Estrada, M.; Perez-Torres, A.; Castell-Rodriguez, A.E.; Becerril-Millan, R.; Rodri-guez-Monroy, M.A.; Jarquin-Yañez, K.; Canales-Martinez, M.M. Wound Healing Activity of α-Pinene and α-Phellandrene. Molecules 2021, 26, 2488. [Google Scholar] [CrossRef]
- Rocha Caldas, G.F.; Oliveira, A.R.; da Silva Oliveira, A.R.; Araújo, A.V.; Lafayette, S.S.L.; Albuquerque, G.S.; Silva-Neto, J.; da Costa Silva-Neto, J.; Costa-Silva, J.H.; Ferreira, F.; et al. Gastroprotective Mechanisms of the Mono-terpene 1,8-Cineole (Eucalyptol). PLoS ONE 2015, 10, e0134558. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.A.; Ho, M.T.; Song, Y.W.; Cho, M.; Cho, S.K. Camphor Induces Proliferative and Anti-Senescence Ac-tivities in Human Primary Dermal Fibroblasts and Inhibits UV-Induced Wrinkle Formation in Mouse Skin. Phytother. Res. 2015, 29, 1917–1925. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.; Ngo, H.T.T.; Park, B.; Seo, S.-A.; Yang, J.-E.; Yi, T.-H. Myrcene, an Aromatic Volatile Compound, Ameliorates Human Skin Extrinsic Aging via Regulation of MMPs Production. Am. J. Chin. Med. 2017, 45, 1113–1124. [Google Scholar] [CrossRef]
- Papanastasiou, S.A.; Bali, E.-M.D.; Ioannou, C.S.; Papachristos, D.P.; Zarpas, K.D.; Papadopoulos, N.T. Toxic and Hormetic-like Effects of Three Components of Citrus Essential Oils on Adult Mediterranean Fruit Flies (Ceratitis Capitata). PLoS ONE 2017, 12, e0177837. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Bedmar, Z.; Anter, J.; de La Cruz-Ares, S.; Muñoz-Serrano, A.; Alonso-Moraga, Á.; Pérez-Guisado, J. Role of Citrus Juices and Distinctive Components in the Modulation of Degenerative Processes: Genotoxi-city, Antigenotoxicity, Cytotoxicity, and Longevity in Drosophila. J. Toxicol. Env. Health A 2011, 74, 1052–1066. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, R.; Kanimozhi, G.; Madahavan, N.R.; Agilan, B.; Ganesan, M.; Prasad, N.R.; Rathinaraj, P. Al-pha-Pinene Attenuates UVA-Induced Photoaging through Inhibition of Matrix Metalloproteinases Expres-sion in Mouse Skin. Life Sci. 2019, 217, 110–118. [Google Scholar] [CrossRef]
- Rodenak-Kladniew, B.; Castro, A.; Stärkel, P.; Galle, M.; Crespo, R. 1,8-Cineole Promotes G0/G1 Cell Cycle Arrest and Oxidative Stress-Induced Senescence in HepG2 Cells and Sensitizes Cells to Anti-Senescence Drugs. Life Sci. 2020, 243, 117271. [Google Scholar] [CrossRef]
- Council of Europe European Pharmacopoeia, 7th ed.; Directorate for the Quality of Medicines & HealthCare of the Council of Europe: Strasbourg, France, 2010; ISBN 978-92-871-6700-2.
- Cavaleiro, C.; Salgueiro, L.R.; Miguel, M.G.; Proença da Cunha, A. Analysis by Gas Chromatography–Mass Spectrometry of the Volatile Components of Teucrium Lusitanicum and Teucrium Algarbiensis. J. Chromatogr. A 2004, 1033, 187–190. [Google Scholar] [CrossRef] [Green Version]
- Linstrom, P.; Mallard, W.G. NIST Chemistry WebBook. Available online: https://webbook.nist.gov/chemistry/ (accessed on 23 November 2022).
- McLafferty, F.W. Wiley Registry of Mass Spectral Data 9th/NIST 08; Mass Spectral Libraly: Gaithersburg, MD, USA, 2009. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Clinical and Laboratory Standards Institute Reference Method for Broth Dilution Antifungal Susceptibility Test-ing of Yeasts; Approved Standard M27-A3, 3rd ed.; Clinical and Laboratory Standards Institute: Wanye, PA, USA, 2008; ISBN 1-56238-666-2.
- Clinical and Laboratory Standards Institute Reference Method for Broth Dilution Antifungal Susceptibility Test-ing of Filamentous Fungi; Approved Standard M38-A2, 2nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008; ISBN 1-56238-668-9.
- Marichal, P.; Gorrens, J.; Cutsem, J.; Bossche, H. Vanden Culture Media for the Study of the Effects of Azole Derivatives on Germ Tube Formation and Hyphal Growth of C. Albicans/Nährböden Zur Untersuchung Der Wirkungen von Azolderivaten Auf Die Keimschlauchbildung Und Das Hyphenwachstum Bei C. Albicans. Mycoses. 2009, 29, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Alves-Silva, J.M.; Guerra, I.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Figueirinha, A.; Salgueiro, L. Chemi-cal Composition of Crithmum Maritimum L. Essential Oil and Hydrodistillation Residual Water by GC-MS and HPLC-DAD-MS/MS, and Their Biological Activities. Ind. Crop. Prod. 2020, 149, 112329. [Google Scholar] [CrossRef]
- Taweechaisupapong, S.; Singhara, S.; Lertsatitthanakorn, P.; Khunkitti, W. Antimicrobial Effects of Boesen-bergia Pandurata and Piper Sarmentosum Leaf Extracts on Planktonic Cells and Biofilm of Oral Pathogens. Pak. J. Pharm. Sci. 2010, 23, 224–231. [Google Scholar] [PubMed]
- Taweechaisupapong, S.; Ngaonee, P.; Patsuk, P.; Pitiphat, W.; Khunkitti, W. Antibiofilm Activity and Post Antifungal Effect of Lemongrass Oil on Clinical Candida Dubliniensis Isolate. S. Afr. J. Bot. 2012, 78, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Saharkhiz, M.J.; Motamedi, M.; Zomorodian, K.; Pakshir, K.; Miri, R.; Hemyari, K. Chemical Composition, Antifungal and Antibiofilm Activities of the Essential Oil of Mentha piperita L. ISRN Pharm. 2012, 2021, 718645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of Nitrate, Nitrite, and [15N]Nitrate in Biological Fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Piras, A.; Maccioni, A.; Falconieri, D.; Porcedda, S.; Gonçalves, M.J.; Alves-Silva, J.M.; Silva, A.; Cruz, M.T.; Salgueiro, L.; Maxia, A. Chemical Composition and Biological Activity of Essential Oil of Teucrium Scordium L. Subsp. Scordioides (Schreb.) Arcang. (Lamiaceae) from Sardinia Island (Italy). Nat. Prod. Res. 2021, 36, 1–8. [Google Scholar] [CrossRef]
- Martinotti, S.; Ranzato, E. Scratch Wound Healing Assay. In Epidermal Cells. Methods in Molecular Biology; Turksen, K., Ed.; Humana: New York, NY, USA, 2019; Volume 2109, pp. 225–229. [Google Scholar]
- Moreira, P.; Sousa, F.J.; Matos, P.; Brites, G.S.; Gonçalves, M.J.; Cavaleiro, C.; Figueirinha, A.; Salgueiro, L.; Batista, M.T.; Branco, P.C.; et al. Chemical Composition and Effect against Skin Alterations of Bioactive Ex-tracts Obtained by the Hydrodistillation of Eucalyptus Globulus Leaves. Pharmaceutics 2022, 14, 561. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp. RIa | Ref. RIa | Exp. RIa | Ref. RIb | Compound * | % |
|---|---|---|---|---|---|
| 919 | 920 | 1030 | 1030 | Tricyclene | 0.2 |
| 928 | 930 | 1030 | 1030 | α-Pinene | 2.3 |
| 943 | 943 | 1075 | 1075 | Camphene | 2.6 |
| 968 | 970 | 1117 | 1118 | β-Pinene | 29.6 |
| 978 | 980 | 1160 | 1162 | Myrcene | 15.4 |
| 1011 | 1011 | 1275 | 1275 | p-Cymene | 0.3 |
| 1018 | 1019 | 1215 | 1215 | 1,8-Cineole | 2.1 |
| 1018 | 1019 | 1204 | 1205 | Limonene | 5.7 |
| 1033 | 1035 | 1246 | 1245 | E-β-Ocimene | 0.3 |
| 1044 | 1046 | 1249 | 1249 | γ-Terpinene | 0.2 |
| 1074 | 1076 | 1288 | 1088 | Terpinolene | 0.2 |
| 1118 | 1118 | 1517 | 1515 | Camphor | 3.2 |
| 1119 | 1119 | 1647 | 1647 | E-Pinocarveol | 0.9 |
| 1144 | 1144 | 1695 | 1695 | Borneol | 16.9 |
| 1159 | 1158 | 1595 | 1597 | Terpinene-4-ol | 1.0 |
| 1166 | 1165 | 1624 | 1622 | Myrtenal | 0.9 |
| 1171 | 1169 | 1694 | 1692 | α-terpineol | 0.5 |
| 1176 | 1176 | 1786 | 1786 | Myrtenol | 0.2 |
| 1199 | 1196 | 1831 | 1830 | trans-Carveol | 0.0 |
| 1231 | 1232 | 1849 | 1852 | Geraniol | 0.4 |
| 1237 | 1235 | n.d. | - | Chrysanthenyl acetate | 0.2 |
| 1261 | 1264 | 1575 | 1574 | Bornyl acetate | 2.6 |
| 1362 | 1359 | 1750 | 1746 | Geranyl acetate | 0.2 |
| 1369 | 1368 | 1487 | 1487 | α-Copaene | 0.1 |
| 1442 | 1445 | 1637 | 1637 | allo-Aromadendrene | 0.3 |
| 1470 | 1466 | 1703 | 1699 | Germacrene D | 0.1 |
| 1477 | 1482 | 1735 | 1735 | Bicyclogermacrene | 0.1 |
| 1507 | 1508 | 1751 | 1751 | δ-Cadinene | 0.3 |
| 1522 | 1521 | 1763 | 1763 | β-Sesquiphellandrene | 0.1 |
| 1526 | 1525 | 2070 | 2073 | Elemol | 0.2 |
| 1550 | 1553 | 2113 | 2113 | Spathulenol | 0.5 |
| 1555 | 1557 | 1977 | 1975 | Caryophyllene oxide | 0.4 |
| 1574 | 1578 | 2074 | 2077 | Guaiol | 0.4 |
| 1621 | 1619 | 2180 | 2187 | α-Muurolol | 1.0 |
| Monoterpene hydrocarbons | 56.8 | ||||
| Oxygen-containing monoterpenes | 29.1 | ||||
| Sesquiterpene hydrocarbons | 1.0 | ||||
| Oxygen-containing sesquiterpenes | 2.7 | ||||
| Total identified | 89.6 | ||||
| Strains | S. rosmarinifolia Essential Oil | |
|---|---|---|
| MIC (a) | MLC (a) | |
| Candida albicans ATCC 10231 | 0.29 | 0.29 |
| C. tropicalis YPO128 | 0.29 | 0.57 |
| C. krusei LF33 | 0.29 | 0.29 |
| C. guillermondii MAT23 | 0.14 | 0.29 |
| C. parapsilosis ATCC 90018 | 0.29 | 0.57 |
| Cryptococcus neoformans YPO186 | 0.14 | 0.29 |
| Yarrowia lipolytica ISA 1774 | 0.14 | 0.29 |
| Trichophyton mentagrophytes FF7 | 0.07 | 0.07 |
| T. rubrum CECT 2794 | 0.14 | 0.14 |
| T. mentagrophytes var. interdigitale CECT 2958 | 0.07 | 0.14 |
| T. verrucosum CECT 2992 | 0.07 | 0.29 |
| Microsporum canis FF1 | 0.07 | 0.14 |
| M. gypseum CECT 2908 | 0.07 | 0.14 |
| Epidermophyton floccosum FF9 | 0.14 | 0.14 |
| Aspergillus niger ATCC16404 | 0.29 | 1.13 |
| A. fumigatus ATCC 46645 | 0.14 | 0.57 |
| A. flavus F44 | 0.29 | 0.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves-Silva, J.M.; Gonçalves, M.J.; Silva, A.; Cavaleiro, C.; Cruz, M.T.; Salgueiro, L. Chemical Profile, Anti-Microbial and Anti-Inflammaging Activities of Santolina rosmarinifolia L. Essential Oil from Portugal. Antibiotics 2023, 12, 179. https://doi.org/10.3390/antibiotics12010179
Alves-Silva JM, Gonçalves MJ, Silva A, Cavaleiro C, Cruz MT, Salgueiro L. Chemical Profile, Anti-Microbial and Anti-Inflammaging Activities of Santolina rosmarinifolia L. Essential Oil from Portugal. Antibiotics. 2023; 12(1):179. https://doi.org/10.3390/antibiotics12010179
Chicago/Turabian StyleAlves-Silva, Jorge M., Maria José Gonçalves, Ana Silva, Carlos Cavaleiro, Maria Teresa Cruz, and Lígia Salgueiro. 2023. "Chemical Profile, Anti-Microbial and Anti-Inflammaging Activities of Santolina rosmarinifolia L. Essential Oil from Portugal" Antibiotics 12, no. 1: 179. https://doi.org/10.3390/antibiotics12010179
APA StyleAlves-Silva, J. M., Gonçalves, M. J., Silva, A., Cavaleiro, C., Cruz, M. T., & Salgueiro, L. (2023). Chemical Profile, Anti-Microbial and Anti-Inflammaging Activities of Santolina rosmarinifolia L. Essential Oil from Portugal. Antibiotics, 12(1), 179. https://doi.org/10.3390/antibiotics12010179